
Abstract
Introduction
C
VIM-1 was first described in Verona, Italy, from a clinical Pseudomonas aeruginosa isolate. 24 The gene of this β-lactamase-type was found on a class 1 integron located on the chromosome. 24 More recently, blaVIM-1 has been found to be predominantly associated with class 1 integrons located on plasmids of the incompatibility type IncN.6,9 Occurrence of these plasmids has also been linked to resistance to other antimicrobial classes such as aminoglycosides and chloramphenicol. 11 Plasmid-borne carbapenemases are mainly found in Enterobacteriaceae, P. Aeruginosa, and Acinetobacter baumannii. 9 K. pneumoniae is presumed to be the main reservoir of plasmid-mediated carbapenemases. 1
In Europe, carbapenemase-producing bacteria are usually cultured from hospital settings and are only rarely identified in the community 5 and in the environment. 36
In a recent study, we cultured an environmental isolate of K. pneumoniae containing a blaVIM-1 gene, from river water in Switzerland. 37 The DNA sequence and subsequent analysis of this plasmid is presented in this study.
Materials and Methods
Bacterial strain
K. pneumoniae spp. pneumoniae OW16C2 with multilocus sequence typing (MLST) sequence type (ST)-1335 was originally isolated from the Rhine River in Switzerland. 37 MLST typing was performed as described by Diancourt et al. 14
Conjugation experiments of pOW16C2 and plasmid characterization
Conjugation experiments were performed with the plasmid-free recipient strain Escherichia coli HK225 (Strepr, Rifr). 23 Briefly, single colonies of the donor and recipient were inoculated in LB broth (Difco Laboratories) and grown overnight at 37°C. Subsequently, equal volumes of the donor and recipient cultures were mixed and incubated overnight at 37°C without shaking. Serial dilutions were then plated on LB agar (Difco Laboratories) selection plates supplemented with a combination of 600 μg/ml streptomycin (Sigma-Aldrich) and 2 μg/ml imipenem (Sigma-Aldrich). Colonies from the selection plates were examined by pulsed-field gel electrophoresis (PFGE) to ensure acquisition of the plasmid. 18
The conjugation frequency per donor was determined by plating serial dilutions of the mating on selective plates on which the donor strain and the conjugant can grow (LB-agar supplemented with 2 μg/ml imipenem) as well as on plates on which only the transconjugants can grow (LB-agar supplemented with 600 μg/ml streptomycin, 100 μg/ml rifampicin, and 2 μg/ml imipenem). The transfer frequency was calculated by the quotient of the number of transconjugants over the number of transconjugant plus donor. Southern blot analysis with a DIG-labeled blaVIM probe (PCR DIG Probe Synthesis Kit; Roche) was performed to confirm the presence of blaVIM on the transferred plasmid. 21 PCR was performed to (1) identify the blaVIM gene acquired by the transconjugants 17 and (2) for replicon typing of the transferred plasmid containing blaVIM. 8 The plasmid size was estimated using S1 nuclease (Thermo Scientific) as described elsewhere.3,35
OW16C2, the acceptor strain E. coli HK225 and the transconjugant (TC OW16C2) were subjected to susceptibility testing against 12 antimicrobial agents by the disc diffusion method according to the CLSI protocols and evaluated according to the CLSI criteria (Clinical and Laboratory Standards Institute, 2011). 10 The antibiotics tested were: ampicillin (AM), amoxicillin/clavulanic acid (AMC), cephalothin (CF), cefotaxime (CTX), ciprofloxacin (CIP), gentamicin (GM), tetracycline (TE), streptomycin (S), chloramphenicol (C), nalidixic acid (NA), sulfamethoxazole (SMZ), and trimethoprim (TMP) (Becton Dickinson). Minimal inhibitory concentration (MIC) for imipenem was determined using Etest strips (bioMérieux).
Plasmid extraction and sequencing
The plasmid pOW16C2 was extracted using the Qiagen Large-Construct Kit (Qiagen) according the manufacturer's protocol. After DNA extraction, an additional phenol purification of the plasmid-DNA was performed. 31
The complete plasmid sequence was determined using the Pacific Biosciences SMRT sequencing approach (Functional Genomics Center) applying P4 chemistry and an RSII device. A 120 min movie was recorded from a 10 kb insert library.
Bioinformatics
Sequence assembly was carried out using the SMRT-Analysis software version 2.0 with the RS_HGAP_Assembly.1 algorithm and default settings. The plasmid sequence was automatically annotated using the online Rapid Annotation Subsequencing Technology (RAST) 2 and the CLC Genomics Workbench Version 6.5 (CLC bio). Automated annotation was manually refined.
Nucleotide sequence
The GenBank accession number for pOW16C2 plasmid is KF977034.
Results
Transferability of pOW16C2
The blaVIM-1-carrying plasmid pOW16C2 could be transferred by conjugation from strain OW16C2 to the recipient strain E. coli HK225 with a conjugation frequency of 2.8×10−3 per donor cell. Transfer of only one plasmid into the recipient was confirmed by PFGE (data not shown) along with PCR-based replicon typing. 8 Moreover, the presence of blaVIM was confirmed by PCR and Southern blot with a DIG-labeled probe.
Resistance profiles of donor (OW16C2), recipient (HK225), and transconjugant (TC OW16C2) strains were summarized in Table 1. According to the resistance profiles, resistance genes conferring a reduced susceptibility to β-lactams, including the carbapenem, imipenem, sulfamethoxazole, and trimethoprim were transferred. These findings are consistent with the resistance genes on the plasmid that are listed below.
Resistance to specific antibiotics are in bold type.
AM, ampicillin; AMC, amoxicillin-clavulanic acid; CF, cephalothin; CTX, cefotaxime; CIP, ciprofloxacin; NA, nalidixic acid; GM, gentamicin; S, streptomycin; TE, tetracycline; C, chloramphenicol; SMZ, sulfamethoxazole; TMP, trimethoprim; IP, imipenem.
pOW16C2 DNA sequence
The complete sequence of pOW16C2 was assembled from 44,335 post-filter reads with a mean read length of 4,376 bp and an individual read accuracy of 0.855. The final coverage over the full plasmid sequence is 2,463 and the accuracy of the assembled plasmid sequence was determined to be 99.99831%. The plasmid consists of 59,228 bp and belongs to incompatibility group N (G+C content, 52.1%) and pMLST ST 7. Plasmid pOW16C2 contains 66 predicted open reading frames (ORF), including blaVIM-1, aacA4, aphA15, aadA1, catB2, qnrS1, sul1, and dfrA14 conferring resistance to carbapenems and other β-lactams, aminoglycosides, chloramphenicol, quinolones, sulfonamides, and trimethoprim, respectively (Table 2).
The backbone of pOW16C2 contains conserved regions typical of an IncN structure, including the repA replicase gene, the tra genes involved in conjugal transfer, 27 the mucA-umuC operon (mutagenesis: UV and chemical) involved in UV protection, 29 the ArdA-ArdB anti-restriction system, 13 as well as the StbA-StbB-StbC system responsible for a stable plasmid inheritance during bacterial multiplication (Fig. 1). 27

Major structural features of pOW162C in comparison with plasmids pKOX105 (accession no.: HM126016) and pR46 (accession no.: AY046276). Areas shaded in gray indicate homologies in plasmid scaffold regions. Antibiotic resistance genes are indicated by red boxes. The individual conjugation-related genes (associated with tra) are indicated with capital letters inside the yellow boxes. Blue boxes denote transposon-, integron-, or replication-associated genes. Green boxes indicate maintenance- and stability-related genes and white boxes indicate hypothetical proteins. The origin of transfer oriT is indicated by a blue circle. (The figure is not drawn to scale.)
Furthermore, the plasmid encodes for the EcoRII endonuclease and EcoRII cytosine-methyltransferase as well as KikA, which is responsible for the killing of K. pneumoniae phenotype, as well as the Kik phenotype cognate-protecting region(s) korA and korB. 32
Ancillary sequences identified on pOW16C2
Plasmid pOW16C2 contains a Tn3-like composite transposon (G+C content: 54.6%) with IS26-like insertion elements as flanking regions (Fig. 1). Within this transposon lies a class 1 integron (29,946–35,837 bp, G+C content: 54.2%) containing a 5′-conserved structure (CS) with the site-specific integrase encoded by int1, and it is a locus containing five gene cassettes, oriented in a classical head-to-tail arrangement and consisting of the genes blaVIM-1-aacA4-aphaA15-aadA1-catB2. The 3′-CS includes the characteristic qacEΔ1-encoding gene 28 along with sulI, which confers resistance to sulfonamides. 33 Furthermore, a qnrS1 gene was also present within the transposon region; however, the latter was not part of the class 1 integron.
Apart from the integron within the composite transposon region, we identified another class 1 integron in the 3,423 bp resistance region. Carattoli et al. described the same integron in a blaVIM-1 harboring plasmid pKOX105, including the 3′-CS, the dfrA14 trimethoprim resistance gene, and it is as well truncated by the insertion of an IS6100 element. 7
Comparison of pOW16C2 with other IncN plasmids
A comparative analysis of pOW16C2 with other IncN plasmids was performed; Salmonella typhimurium plasmid pR46, the IncN reference plasmid (AY046276), and Klebsiella oxytoca pKOX105, an IncN and blaVIM-1-carrying plasmid isolated in Italy (HM126016), were used for comparison (Fig. 1). All three plasmids exhibit similarities in functionally conserved regions, such as replication, stability, and transfer systems.
BLASTn searches revealed that all of the building blocks of this Tn3 region could be identified on different plasmids, but the assembly of the genes in the Tn3-like transposon flanking this class 1 integron appears to be a novel feature (Fig. 2). The integron showed a 99% nucleotide sequence similarity to an integron located on a plasmid isolated from a K. pneumoniae (JX185132.1), however this integron carried all of the resistance determinants, except for blaVIM-1 that was replaced by blaVIM-34, which encodes a VIM-type enzyme that differs from VIM-1 by the single amino acid change, valine (113) to isoleucine.
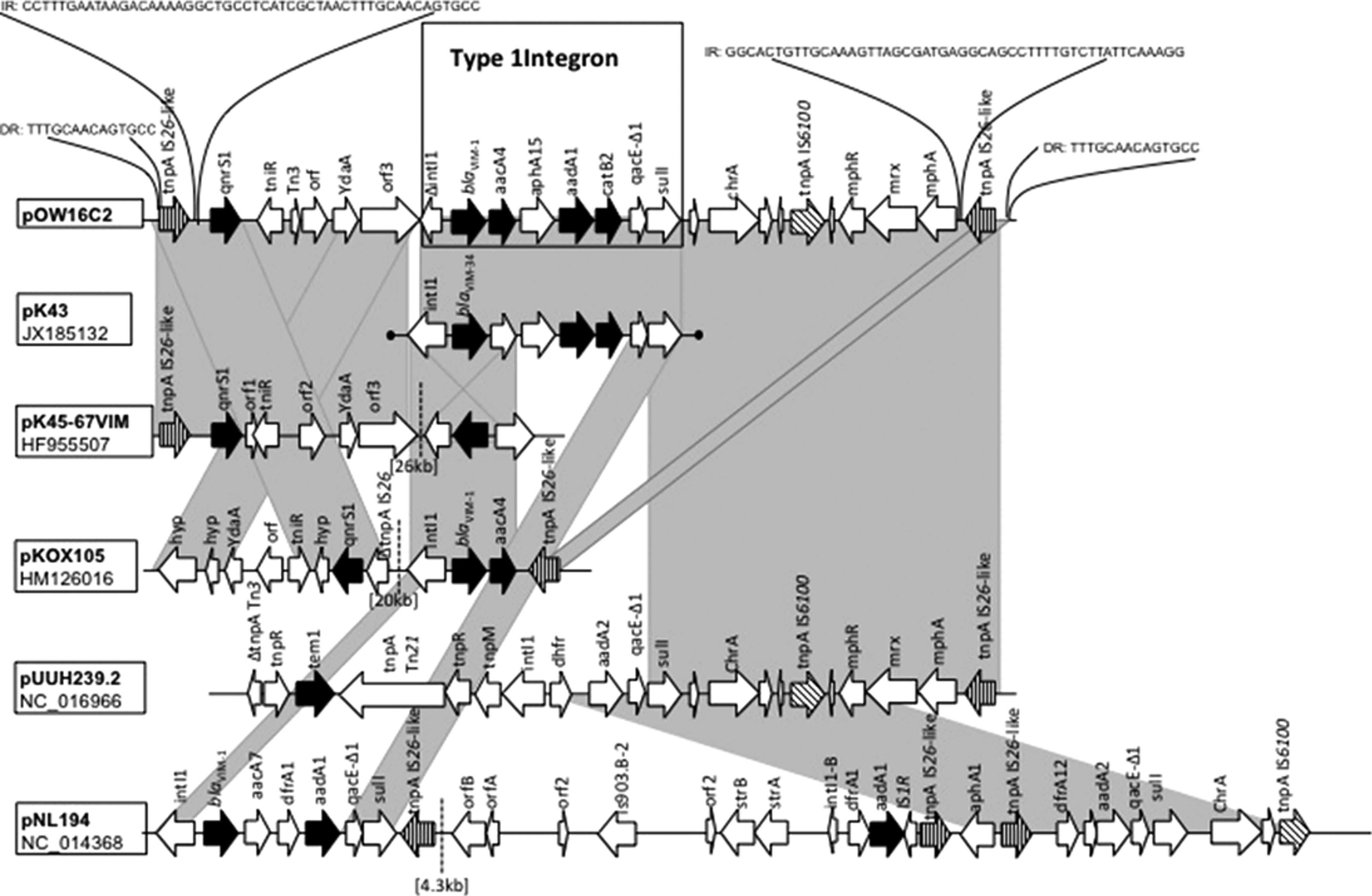
Structural comparison of VIM-1 encoding transposon region of pOW16C2 with MDR plasmids (pK45-67VIM: HF955507; pKOX105: HM126016; pUUH239.2: NC_016966; pNL194: NC_014368) and the integron region of pK43 (JX185132). Antibiotic resistance determinants that are present on pOW16C2 and on the plasmids, which it was compared to, are marked black. Gray shading connects similar sequences. Dashed lines indicate missing sequence. DR indicates a direct repeat sequence and IR indicates an inverted repeat sequence.
The region proximal to the class 1 integron (located between bases 23,953 and 29,913) includes an IS26-like transposase, the qnrS1 gene, the resolvase/integrase TniR protein, the Tn3-like transposase, two hypothetical proteins, and ydaA (putative transposon resolvase), and this region displays 100% nucleotide sequence identity to K. pneumoniae plasmid pK45-67VIM (HF955507.1). Plasmid pK45-67VIM also contains a blaVIM-1 gene in an IncN type plasmid, although in this case, the VIM-containing locus is found to be inverted and distally located some 26-kb from the qnrS1-carrying region described above. A similar situation was found for the IncN VIM-1 positive plasmid pKOX105 (HM126016) in K. oxytoca. Comparison with the transposon-containing region of pOW16C2 was similar, but inverted (Fig. 2), and as with pK45-67VIM, the blaVIM-1-carrying integron is distally located.
The sequence distal of the 3′-CS of the integron contains a chromate resistance encoded by chrA along with a macrolide resistance operon. This structural arrangement was also identified in pUUH239.2 (NC_016966).
We compared pOW16C2 to pNL194, 25 a blaVIM-1-carrying IncN plasmid isolated from a K. pneumoniae strain in Athens, Greece. Plasmid pOW16C2 varies considerably from pNL194, except in the IncN backbone, although a notable amount of related (resistance) genes were present on both plasmids.
Discussion
Over the last few years, the DNA sequences of a number of IncN type MDR plasmids carrying blaVIM-1 have been obtained and the results of comparative genomics lead to the suggestion that there may be a common ancestor. However, the acquisition of a class 1 integron harboring blaVIM-1 may have occurred several times independently through horizontal gene transfer.7,19,25,30 Studies focusing on IncN plasmids carrying blaCTX-M-1 from animals, environment, and human origin 15 and studies investigating IncN plasmid from waste water treatment plants 16 revealed IncN plasmids exhibiting two major hot spot integration sites of transposable elements. These two regions are between the fipA and nuc genes or in proximity of the EcoRII restriction/antirestriction system, 15 which correspond to the integration site found in pOW16C2. The wide distribution of IncN plasmids in Enterobacteriaceae, the conjugation at high frequency and the stable maintenance in the bacterial host cell is associated with the partitioning and antirestriction system present in the IncN scaffold. 15 Cullik et al. hypothesized that the complete resistance region might jump between different plasmid backbones as composite transposons mediated by an IS26 transposition event. 12 In regard to plasmid evolution, studies addressing plasmid restriction fragment length polymorphism and pMLST gives evidence that there are specific IncN plasmids spreading in Europe disseminating specific resistance genes, whereas the main player for the evolution of IncN plasmids are resistance genes and insertion sequences, which likely can be exchanged even between different plasmid types and the chromosome.12,15
An ongoing epidemic of blaVIM-1-positive K. pneumoniae is found in Greece5,30 and a few local outbreaks were detected in other southern European countries. 5 Plasmid MLST revealed that all blaVIM-1-carrying plasmids from Greece belong to closely related ST 10 or 11, whereas plasmids such as pKOX105 from Italy belong to ST 7. It was, therefore, suggested that blaVIM-1 has spread in Italy and in Greece by different IncN plasmids types. 19 Interestingly, pKOX105 from Italy shares the same ST as pOW16C2.
The occurrence of carbapenemase- and ESBL-producing Enterobacteriaceae in surface water is highly worrisome. The selection of these, often multi-drug resistant (MDR) bacteria, is multifaceted. It has been shown that β-lactam agents are rapidly hydrolyzed, especially under basic conditions, which prevail in municipal sewage and hospital effluents. 22 Thus, other resistance mechanisms might be responsible for the selection, persistence, and dissemination of these MDR bacteria. Several studies have shown that antibiotics exert a selective pressure even at very low concentration, favoring MDR bacteria. 20 A candidate for the co-selection could be fluoroquinolones. High concentrations of fluoroquinolones are found in rivers, in and downstream of wastewater treatment plants. 4 Although quinolone resistance is mainly due to chromosomal mutation in the fluoroquinolone target genes, plasmid-mediated quinolone resistance genes, such as qnrS, which is also found on pOW16C2, might be important in the selection of the MDR bacteria.
To our knowledge, this is the first report of an IncN plasmid carrying blaVIM-1 from a K. pneumoniae strain isolated from surface water. Klebsiella species may play an important role in the dissemination of antibiotic resistance genes, since they are commonly found in the environment and in hospitals. 33 The spread of carbapenemase-producing Klebsiella spp. in surface waters is of concern, since it may be just a matter of time until these plasmids are identified in other Enterobacteriaceae and ubiquitously distributed similarly to the events leading to the emergence of the ESBL-carrying plasmids.
Footnotes
Acknowledgments
The authors thank Magdalena Nüesch-Inderbinen for critically reading the manuscript.
Disclosure Statement
No competing financial interests exist.