
Abstract
Eighty-four multidrug-resistant Klebsiella pneumoniae (MDR-KP) isolates from a Chinese hospital from January to October 2012 were evaluated to characterize the coexistence of 16S rRNA methylase, extended-spectrum β-lactamase, carbapenemase, and plasmid-mediated quinolone resistance determinants and their association with mobile genetic elements. Among the 84 MDR-KP isolates studied, 19 isolates exhibited high-level resistance to amikacin mediated by the production of the 16S rRNA methylase. They carried 19 armA genes (22.9%) and three rmtB genes (3.6%). CTX-M genes were found in all of the isolates. Among these armA- or rmtB/CTX-M-producing K. pneumoniae isolates, 31.6% carried the carbapenemase genes (blaKPC-2 [26.3%], blaIMP-4 [10.5%], and blaNDM-1 [5.3%]), which made them resistant to imipenem (minimum inhibitory concentration [MIC] ≥16 mg/L). All positive strains possessed qnr-like genes (16 qnrA1, 10 qnrS1, and 7 qnrB4 genes) and 18 harbored an aac(6′)-Ib-cr gene. Mobile elements ISEcp1, IS26, ISCR1, ISAba125, and sul-1 integrons were detected in 19/19 (100%), 16/19 (84.2%), 18/19 (94.7%), 9/19 (47.4%), and 18/19 (94.7%) isolates, respectively. The mobilizing elements occurred in different combinations in the study isolates. Majority of armA and qnr genes were in MDR-KP strains carrying integrons containing the ISCR1. Close to 80% of blaTEM-1 and blaSHV-12 were linked to IS26 while ≥90% of blaCTX-Ms and blaCMYs were linked to ISEcp1. ISAba125 was located upstream of blaNDM-1 and some blaCMY-2 genes. In addition, seven transconjugants were available for further analysis, and armA, qnrS1, acc(6′)-Ib-cr, blaCTX-M-15, blaTEM-1, and blaNDM-1 were cotransferred. This study points to the dissemination of 16S rRNA methylase genes and the prevalence of selected elements implicated in evolution of resistance determinants in collection of clinical K. pneumoniae in China.
Introduction
K
The 16S rRNA methylase providing high-level resistance to aminoglycoside antibiotics—including amikacin, tobramycin, and gentamicin—has been identified in various gram-negative bacilli.9,15 Additionally, various 16S rRNA methylase genes—including armA, rtmB, and rtmA—have been widely reported in Japan, 34 Taiwan, 21 Europe, 9 and China. 17 ISEcp1, IS26, ISCR1, and sul-1 integrons are known to mediate transposition and/or expression of multiple resistance genes in their close proximity.30,32 Carriage of physically linked elements, each containing a set of resistance genes, may increase the chances of en bloc horizontal transfer of multiple resistance determinants to susceptible strains. Carriage of multiple resistance elements may in turn confer unique advantages to the host and enable them survive a strong antimicrobial selection pressure especially in hospital settings. 27
To date, the association of 16S rRNA methylase and extended-spectrum β-lactamase (ESBL) determinants has been punctually identified, an occurrence that complicates the natural-selection picture even in the absence of antibiotics. However, the aim of this study was to characterize the mechanisms of resistance to beta-lactams and quinolones associated to 16S rRNA methylase, and to determine the presence of genetic mobile elements described in the literature in multidrug-resistant K. pneumoniae (MDR-KP) isolates collected in China.
Materials and Methods
Bacterial strains and antimicrobial susceptibilities
A total of 84 nonduplicate MDR-KP clinical isolates with MIC ≥2 mg/L to amikicin were collected between January and October 2012 in a university hospital located in the southeastern region of China. The identification of K. pneumoniae was performed with a Vitek 2 automated system (Biomerieux). Antimicrobial susceptibility testing was performed using the automated broth microdilution system (Vitek 2), according to the Clinical and Laboratory Standards Institute. 5 Escherichia coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality control strains in susceptibility tests with results within the expected ranges. E. coli J53 was used in the conjugation experiments. The presence of metallo-β-lactamase (MBL) production was determined in the K. pneumoniae with the three screening methods (Imipenem-EDTA combined disc test, IMP-EDTA double-disc synergy test, and MBL E-test).16,36 All isolates fulfilled the CLSI criterion for performing carbapenemase detection by the modified Hodge test (MHT) as previously described. 16
DNA preparation, polymerase chain reaction amplification, and sequencing
Genomic DNA was extracted by boiling bacterial colonies for 10 min in sterile distilled water. After centrifugation, the supernatant was used as a DNA template. Single polymerase chain reaction (PCR) was used to analyze genes 16S rRNA methylase (armA, rmtB, npmA, rmtC, and rmtD), plasmid-mediated quinolone resistance [PMQR; qnrA, qnrB, qnrS, and acc(6′)-Ib-cr], ESBL (CTX-M, SHV, and TEM), and carbapenemase genes (KPC, IMP, VIM, NDM, and OXA-48) with specific primers for each one, as previously reported.14,20,35 The strains were also screened for the presence of other β-lactamase genes (blaOXA-1 and blaOXA-10) as well as various mobile elements, such as ISEcp1, ISCR1, IS26, ISAba125, and sul-1 integron. 13 PCR products were purified by using a Qiaquick PCR purification kit (Qiagen). Bidirectional DNA sequencing was performed using the Big-Dye Terminator kit and analyzed by ABI 7300 sequencer (Applied Biosystems). The nucleotides were compared with reference sequences deposited in GenBank databases.
Screening of physical linkages between genetic elements and resistance genes
Physical linkages between insertion sequences or integron and resistance genes were determined using a combination of primers specific for various genetic elements (ISEcp1, ISCR1, IS26, ISAba125, and sul-1 integron) and consensus primers for all the resistance genes. A long-range polymerase (LongAmp Taq DNA Polymerase; New England Biolabs) was used in all reactions for physical linkages. A slow ramping rate of between 0.2°C/sec and 0.3°C/sec was set for the annealing step. The extension time was set at 72°C for 2 min and a final extension of 72°C for 15 min was carried out after 40 cycles of denaturation, annealing, and extension.
Conjugation, transformation, southern hybridization, and plasmid replicon typing
E. coli J53 was used as the recipient in the conjugation studies, and mating was carried out on blood agar medium without selection. After 18 hr, the mixed cultures were taken from the plates, suspended in saline, and plated onto MacConkey medium containing sodium azide (150 μg/ml) and amikacin (30 μg/ml). Confirmation that conjugation had taken place in E. coli J53 was determined by the Vitek 2 automated system and testing for the presence of the armA and rmtB genes by PCR analysis. Plasmid DNAs of donor isolates and E. coli transconjugants were extracted using the Kieser technique. 12 Plasmid DNA was purified by the Qiagen Plasmid Midikit (Qiagen, Inc.). Purified plasmids were used to transform MAX Efficiency DH5α E. coli chemically competent cells (Invitrogen). DH5α was chosen as the recipient because it is capable of being transformed efficiently with large plasmids.
Southern hybridization was performed by standard methods with armA-specific and rmtB-specific digoxigenin (DIG)–labeled probes formed by the PCR DIG detection system (Roche Diagnostics GmbH) as described previously. 37 Plasmid profiles and size estimation were determined by electrophoresis using a 0.7% agarose gel. PCR-based replicon typing of the main plasmid incompatibility groups reported for K. pneumoniae was performed as described. 3 Specifically, 18 replicon types (FIA, FIB, FIC, HI1, HI2, I1-Iγ, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and FIIA) were screened by the methods of Carattoli et al., 3 and additional three types (ColE, R, and U) were investigated by the method of Pallecchi et al. 23 The colE PCR was devised to amplify all the colE-like plasmids. Amplicons were sequenced for confirmation.
Enterobacterial repetitive intergenic consensus polymerase chain reaction and multilocus sequence typing
Enterobacterial repetitive intergenic consensus polymerase chain reaction (ERIC-PCR) was performed and isolates were typed according to their band patterns as described previously. 18 BioNumerics software (version 4.0; Applied Maths) was used to analyze the DNA patterns and determine their similarity. Multilocus sequence typing (MLST) of K. pneumoniae was performed as described by Liu et al. 19 Experimentally determined DNA sequences were uploaded into the MLST database (www.pasteur.fr/recherche/genopole/PF8/mlst/K.pneumoniae.html), and allelic numbers as well as sequence types (STs) were obtained.
Results
Antimicrobial drug susceptibility
Among the 84 MDR-KP isolates, 19 highly amikacin-resistant isolates (MICs >32 mg/L) were collected and the aminoglycoside resistance was due to the production of the 16S rRNA methylase (armA or rmtB). The majority of the isolates were recovered from blood samples (9/47.4%), followed by surgical wounds (5/26.3%), urine (3/15.8%), and catheter tips (2/10.5%). Antimicrobial resistance rates against 19 MDR-KP isolates are shown in Table 1. Most of the isolates showed high-level resistance for aminoglycosides (amikacin, gentamicin, and tobramycin) and quinolone (ciprofloxacin and levofloxacin); for the aminolycosides, MIC50 was >64 mg/L and for the quinolone, MIC50 was >16 mg/L. Thirteen out of the 19 isolates showed resistance to imipenem (MIC50 of 32 mg/L) and all of the isolates were susceptible to tigecycline (MIC50 of 0.38 mg/L, respectively). Seven isolates showed a positive result in the MHT. In addition, three isolates were phenotypically positive for MBL production by the three screening methods used. Overall, all of the isolates showed resistance to most of the tested antimicrobial agents excluding tigecycline (Table 1).
MIC, minimum inhibitory concentration.
Resistance genes
PCR and sequencing performed as previously described revealed that all 19 isolates possessed armA or rmtB gene. PCR on the total DNA of all armA- or rmtB-positive isolates showed different β-lactamase gene combinations (Table 2). In addition to blaCTX-Ms, isolates also harbored several acquired and plasmid-encoded genes: blaSHV-12, 31.6%; blaTEM-1, 100%; blaOXA-1, 100%; blaKPC-2, 26.3%; blaNDM-1, 5.3%; blaIMP-4, 10.5%; and blaCMY-2, 42.1% (Table 2).
Resistance genes and plasmids observed in the respective transconjugants are underlined.
ERIC-PCR, enterobacterial repetitive intergenic consensus polymerase chain reaction; ESBL, extended-spectrum β-lactamase; ND, not determined or not present; PMQR, plasmid-mediated quinolone resistance; ST, sequence type.
Interestingly, genes for qnr or aac(6′)-Ib-cr were detected in 100% of the 19 isolates (Table 2). Cooccurrence of PMQR determinants was often observed, being of different types [qnrA1, qnrB4, qnrS1, and aac(6′)-Ib-cr]. Among the PMQR genes, the presence of qnrA1 was detected in 16 isolates, including 15 coexisting with acc(6′)-Ib-cr gene, 9 coharboring with qnrS1 gene, and 7 cocarrying with qnrB4 gene. Various combinations of blaCTX-Ms, blaCMY-2, blaKPC-2, qnr, aac(6′)-Ib-cr, and armA genes were found in majority of the isolates.
Mobile elements ISEcp1, IS26, ISCR1, ISAba125, and sul-1 integron were detected in 19/19 (100%), 16/19 (84.2%), 18/19 (94.7%), 9/19 (47.4%), and 18/19 (94.7%) isolates, respectively. The isolates were also analyzed in reference to various combinations of observed genetic elements, and ISEcp1 plus ISCR1 plus sul-1 integron was found to be the most common combination, followed by ISEcp1 plus IS26 plus ISCR1 plus sul-1 integron. The frequencies of occurrence of mobile element combinations were different in the different resistance genes (Table 3).
The bold figures mean that the resistance genes have more common associations with the genetic elements.
Analysis and transferability of armA-carrying plasmids
Nine of the 18 replicons tested—including I1, L/M, FrepB, FIA, FIB, FIC, A/C, colE, and N—were involved in the donors. In case of their incompatibility groups, IncL/M (9/19), IncA/C (7/19), and IncFIA (7/19) were the dominant ones. High-level aminoglycoside resistance could be transferred to E. coli J53 by conjugation with frequencies between 10−5 and 10−7 only from seven armA-positive donors through repeat attempts including using filter mating (data not shown). All the seven donor strains belonged to ST15. The transconjugants were highly resistant to amikacin, gentamicin, and tobramycin. The results of PCR and sequence analysis revealed that qnrS1, acc(6′)-Ib-cr, blaTEM-1, blaCMY-2, blaNDM-1, and blaCTX-M-15 were cotransferred with armA on plasmids to the recipients (Table 2). To analyze plasmids carrying the armA gene, plasmid DNA was prepared and transfer to E. coli DH5α by electroporation was attempted. armA-positive plasmids were successfully transferred by transformation from all the parental strains to the recipient E. coli DH5α strain. The transformants harbored a single plasmid with a size of ∼65 kb (data not shown). The plasmids of armA-positive isolates and their transconjugants were hybridized with the armA-specific probe (Fig. 1). The results of PCR typing revealed that the armA-carrying plasmids in this study all belonged to the IncA/C incompatibility group.
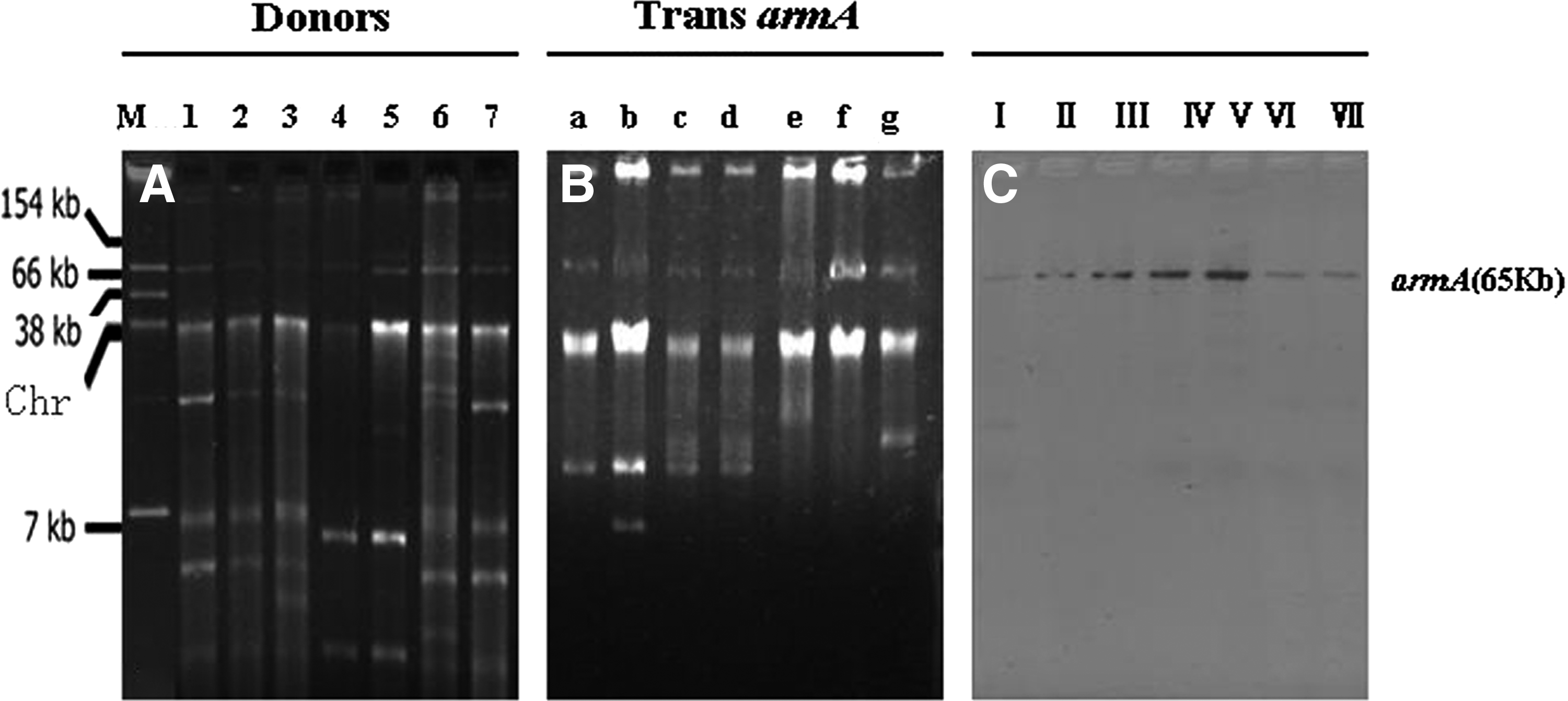
The plasmid profile of the donors
Physical linkages between resistance genes and genetic elements
Table 3 illustrates selected examples of physical linkages between resistance genes and different genetic elements. Thirteen (68.4%) of the 19 armA were borne on strains containing the ISCR1 while 26.3% were borne on strains linked to the ISEcp1. However, only the ISCR1 element and sulI integrons were found upstream of rmtB. Close to 80% of isolates carrying blaTEM-1 and blaSHV-12 were physically linked to the IS26 as reported in Belgium and Germany (Table 3). In all cases, the ISEcp1 was the most common IS element associated with blaCTX-M-14, blaCTX-M-15, blaOXA-1, and blaCMY-2. Majority (81.3%) of qnr genes were physically linked to either integron-associated ISCR1 or the ISEcp1. In all cases tested, the qnrA1 gene was found immediately downstream of ISCR1 as part of a complex class 1 integron that included the 3′CS duplication. All isolates carrying aac(6′)-lb-cr or the qnr gene contained multiple genetic elements and were all MDR. ISAba125 was located upstream of the blaNDM-1 gene and blaCMY-2 gene.
MLST and ERIC-PCR
The 19 AmrA- or RmtB/CTX-M-producing isolates were evaluated by using the PubMLST (http://pubmlst.org/). By combining the seven gene loci, seven different STs (ST11, ST15, ST23, ST37, ST39, ST147, and ST395) were assigned for the isolates and the most dominant clone was ST15, with seven isolates (36.8%)—four belonged to ST395. The analysis of STs by eBURST (http://eburst.mlst.net/) showed that ST 11 and ST 395 are single-locus variants of ST 258. Of these isolates, seven belonged to CC14 and six belonged to CC11. These results mostly corresponded with the results of ERIC profile analysis, which revealed two clusters showing >60% similarity (clusters I and II). Cluster I comprised seven isolates belonging to CC14 and cluster II comprised six isolates belonging to CC11 (data not shown).
Discussion
K. pneumoniae has globally emerged as one of the most troublesome pathogens due to its remarkable ability to acquire genetic determinants for antibiotic resistance. Carbapenem, aminoglycosides, and quinolone antibiotics have been efficiently used in the treatment of the infection with K. pneumoniae. However, increased manifestations of antimicrobial resistance mediated by plasmids, especially 16S rRNA methylase, PMQR, and carbapenemase in MDR K. pneumoniae, have been observed in China. 35
In this study, we tried to identify β-lactamase and PMQR genes in K. pneumoniae isolates highly resistant to aminoglycosides. Nineteen highly amikacin-resistant isolates carried armA gene while rmtB was found in three K. pneumoniae isolates. However, the MICs of amikacin for the other 65 isolates negative for armA and rmtB in the PCR assays ranged between 2 and 16 mg/L. The aac(6′)-Ib gene (encoding an aminoglycoside modifying enzyme) was detected in association with either ant(2″)-Ia or aac(3)-IIc in all armA- and rmtB-negative strains (data not shown). Additionally, most isolates had high-level resistance to beta-lactams by the expression of ESBLs and carbapenemase genes. The ESBL CTX-M types were found in all of the armA- or rmtB-positive isolates distributed as CTX-M-14 and CTX-M-15. Additionally, only six isolates possessed the carbapenemase genes, but 14 of the 19 isolates had an imipenem MIC of >4 mg/L, suggesting an alternative mechanism of carbapenem resistance, like expression of ESBL plus impermeability. Previous reports have described the concurrence of blaNDM-1 together with 16S rRNA methylase and additional ESBLs, mainly armA and CTX-M-15. 29 In our study, one donor strain had armA, qnrA1, qnrS1, acc(6′)-Ib-cr, blaKPC-2, blaCTX-M-15, blaTEM-1, acc(6′)-Ib, blaOXA-1, and blaNDM-1, but only qnrS1, acc(6′)-Ib-cr, blaCTX-M-15, blaTEM-1, and blaNDM-1 were cotransferred with armA (Kpn14/ST15). In addition, some donor strains transferred two or more conjugative plasmids by conjugation. Occasionally plasmids in two or more different incompatibility groups have closely similar conjugation systems.
On the other hand, the strain Kpn14 that coproduced the NDM-1 and KPC-2 carbapenemase belonged to the ST15, which was previously associated with the spread of the NDM-1 carbapenemase in China. 11 On the contrary, ST15 has been associated not only with NDM carbapenemase but also with CTX-M-15 and armA in different parts of the world.6,25
Association of 16S rRNA methylase and ESBL genes in one transferable plasmid was suggestive for seven donor isolates, according to the resistance phenotypes displayed by the transconjugants. Hybridization experiments confirmed that the armA gene was located on an ∼65-kb plasmid. The plasmid harbored by the transconjugants belonged to the incompatibility group IncA/C as determined by PCR typing. The IncA/C plasmid family plays a major role in the dissemination of antimicrobial resistance in K. pneumoniae.1,31 This type of plasmid was previously known to be involved in the transmission of NDM-1, CMY-2, and CTX-M-15 among K. pneumoniae isolates and has been also prevalent in E. coli species.4,26 The IncA/C plasmids may be quite unstable, as reported previously. 22 Highly efficient transmission of armA-carrying plasmids may explain the diversity and worldwide spread of armA-carrying Enterobacteriaceae. In our study, IncFIA, IncA/C, and IncL/M were detected most frequently. We hypothesize that plasmids of different incompatibility groups have acquired similar or identical sets of resistance genes and this acquisition is mediated by genetic elements such as those investigated in this study. Therefore, there is a possibility that such elements act as genetic shuttles between plasmids of different in-compatibility grouping. The similarities and differences in genetic environments of bla, aac (6′)-lb-cr, and qnr genes reported previously may reflect a difference in transposition activities of such elements.13,28 We further hypothesize that differences in antibiotic-use patterns in different regions influence the transposition activity of such elements.
The present report is also unique in that it analyzes the battery of associated mobile elements and the frequencies of their occurrence, specifically, the occurrence of various combinations. ISEcp1 plus ISCR1 plus sul-1 integrons was the most common combination. The genetic environment of the 16S rRNA methylase genes was also assessed. The armA genes were systematically found downstream of ISCR1 and ISEcp1 elements, as described previously. 24 The gene rmtB was always associated with the ISCR1 element. This may reflect the environmental niche of ISCR1 elements or may simply indicate that there are insufficient data concerning the genetic environments of mobile 16S rRNA methylase genes. Genes blaNDM-1 and blaCMY-2 were found downstream of ISAba125 element, as previously described. 8 This result further indicated that part of the ISAba125 sequence is constantly present upstream of the blaNDM-1 and blaCMY-2 genes in K. pneumoniae. It is therefore likely that this insertion sequence element may have played a role in the mobilization of blaNDM-1 and blaCMY-2, at least in the early processes of its acquisition. It also suggests that the original dissemination of the blaNDM-1 and blaCMY-2 genes occurred in A. baumannii and that its spread among K. pneumoniae might correspond to a secondary event. Majority of PMQR genes were physically linked to either integron-associated ISCR1 or the ISEcp1. Most bla genes are distantly linked to elements that are in turn linked to other resistance genes, such as aac(6′)-Ib-cr and qnr. In all cases, the ISEcp1 was the most common IS element associated with blaCT X-M-14, blaCTX-M-15, and blaCMY-2. Similar reports have been published in Tunisian. 2 In addition, we assessed the clonal relatedness of the 19 amrA or rmtB/CTX-M-producing K. pneumoniae isolates via ERIC-PCR profiles and MLST method, which showed that the isolates were largely not clonal. Our results are concordant with the observation that ST15 was the dominant clone of amrA or rmtB/CTX-M-producing KP in mainland China. K. pneumoniae ST15 has previously been described as a host lineage for the NDM-1 enzyme and is also a frequent host of CTX-M enzymes. 33
In a nutshell, this study reports carriage of 16S rRNA methylase genes in MDR K. pneumoniae strains and their association with multiple genetic elements. Additionally, our results further demonstrated that an IncA/C-type plasmid possesses interesting features, since it contains genes encoding the MBL NDM-1 and the 16S RNA methylase armA. Similar reports have been published in an IncHI1 plasmid in France. 7 Plasmids carrying the armA gene are very diverse in terms of genetic structure. Nevertheless, they accumulate a series of antibiotic-resistance determinants in most cases. The existence of coresistance markers, located on single plasmids, suggests that the process of the selection of armA-carrying plasmids may be associated with many unrelated antibiotic molecules or toxic compounds.
Footnotes
Acknowledgments
This study was supported by grants from the Education Department of Jiangxi Province, China (GJJ14178 and GJJ13200), and the Health and Family Planning Commission of Jiangxi Province (2013A035).
Disclosure Statement
None of the authors have any conflicts of interest to declare concerning the article.