
Abstract
In 2009 during surveillance in a Brazilian hospital, many patients were confirmed to be colonized by vancomycin-resistant enterococci (VRE) and a few infection cases occurred. Among 14 isolates of Enterococcus faecalis, most had the same pulsotype, virulence profile (agg+elrA + gelE + ), and were sequence type (ST)103, indicating dissemination of a clone. The 47 Enterococcus faecium were separated into four pulsotypes, the predominant virulence profile being esp+acm+. All of them harbored the hospital marker IS16, and three randomly chosen isolates were ST412, belonging to the Clonal Complex 17. E. faecalis were all susceptible to penicillin and ampicillin, while all E. faecium were resistant to them. All isolates were susceptible to daptomycin and tigecycline. There were no rep-family genes common to all VRE. The VanA element of all E. faecium lost its left-side inverted repeat (IRL) region and had a specific IS insertion. On the other hand, all E. faecalis presented intact Tn1546. The size of plasmids containing the vanA gene as well as its rep-families varied between and within species. The lack of a vanA plasmid common to all VRE, together with the differences among VanA elements, despite the fact that some patients were colonized by both species during their hospitalization, leads us to suggest at least two different Tn1546 origins.
Introduction
E
Operons related to vancomycin resistance described to date are VanA, -B, -C, -D, -E, -G, -L, -M, -N.9,23,38 In Brazil, the first vancomycin-resistant Enterococci (VRE) case occurred in 1996 in an E. faecium isolated in Parana state harboring the operon VanD.10,11 The first VRE outbreak was in Sao Paulo in 1998 caused by VanA harboring E. faecalis and E. faecium. 39 Resistance to vancomycin in Brazil is mostly the VanA type caused by the presence of a gene cluster on Tn1546. The operon is formed by two regulatory genes (vanR and vanS), four genes related to vancomycin resistance itself (vanA, vanH, vanX, and vanY), two genes for the transposition of the element (orf1 and orf2), and one gene related to teicoplanin resistance (vanZ). 17 There might be variations on the composition of this vancomycin resistance operon due to insertion of IS elements and the variants are described as VanA-like elements.
The aim of this study is to investigate a possible VRE dissemination at the Risoleta Tolentino Neves Hospital, Brazil, from June 2009 to January 2011 and to characterize the VRE mobile elements.
Materials and Methods
Settings and isolates
Risoleta Tolentino Neves Hospital in Belo Horizonte, Minas Gerais state, is a 333-bed hospital serving 11,000 patients/month in its first aid clinic, performing 450 surgeries/month, and providing care for an area comprising 2.4 million inhabitants. During a surveillance program from June 2009 to January 2010, 63 VRE were isolated from 52 patients, 60 from rectal swabs and three from infection sites, and all E. faecium and E. faecalis (n=61) were selected for this study. Four of 52 patients were cocolonized by both E. faecium and E. faecalis.
The isolates were cultured in a brain–heart infusion (BHI) broth and stored with 40% glycerol at −80°C until used.
Species identification, vanA detection, and Tn1546 characterization
Genomic DNA was mechanically extracted following the method described by Palazzo et al. 27
Multiplex-PCR, as described by Dutka-Malen et al., was performed for species identification, using primers for the genes ddlE. faecium (identifies E. faecium), ddlE. faecalis (identifies E. faecalis), vanC-1 (identifies Enterococcus gallinarum), and vanC-2/vanC-3 (identify Enterococcus casseliflavus/Enterococcus flavescens). 14 Quality control strains were E. faecium NCTC7171, E. faecalis NCTC775, E. gallinarum NCTC12359, and E. casseliflavus NCTC1261. When it was not possible to identify the species by this protocol, the identification was made by sequencing a fragment of the gene rpoA. 24
vanA detection was performed through amplification of a 399-bp fragment of the gene, as described by Woodford et al. 36 Once vanA was detected, the whole Tn1546 (10.8 kb) was amplified by long PCR, as described by Woodford et al. 37 When long PCR did not amplify or showed a different pattern than expected, overlapping PCR was performed with primes P1 to P19 following the protocol of Arthur et al. 2 The fragments of overlapping PCR were sequenced to identify the insertion sequences present on them.
The presence of vanB was excluded using a modified multiplex PCR protocol with the primers described by Depardieu et al. 12 : vanA [EA1(+) and EA2(−)], vanB [EB3(+) and EB3(−)], ddl E. faecalis [DD13(+) and DD3-2(−)], and ddl E. faecium [FAC1-1(+) and FAC2-1(−)] genes were searched. The DNA of E. faecalis V583 was used as a positive control for vanB amplification.
Search for virulence factors
The search for the virulence factors cytolysin, hyaluronidase, aggregation substance, collagen adhesin, enterococcal surface protein, gelatinase, and leucine-rich protein A was performed by PCR using the primers described in Table 1. The hospital marker IS16 was searched in the same way. PCR conditions were as follows: 3 minutes at 94°C followed by 30 cycles of 45 seconds at 94°C, 1 minute at the specific annealing temperature (Table 1) and 1 minute at 72°C, and a final step at 72°C for 10 minutes.
The gelatinase production phenotype was verified by inoculating the isolates on trypticase soy agar plates containing 1.5% skim milk and incubating them for 18 hours at 37°C. 18 A clear halo around the colonies indicated proteolysis and thus gelatinase production.
A phenotypic assay for cytolysin production was performed by growing the isolates in BHI agar supplemented with 5% human blood and incubating them for 18 hours at 37°C. Clear zones around colonies indicated positivity. 15
Antimicrobial susceptibility testing
The minimal inhibitory concentration (MIC) was determined for vancomycin, teicoplanin, penicillin, ampicillin, tetracycline, erythromycin, daptomycin, quinupristin–dalfopristin, linezolid, and tigecycline by Etest® (BioMérieux).
Criteria for susceptibility were those recommended by CLSI, 8 except for tigecycline, which followed EUCAST. 34
Screening tests were performed to detect high-level aminoglycoside resistance for both gentamicin and streptomycin according to the CLSI protocol. 8
Molecular typing
Pulsed-field gel electrophoresis (PFGE) was performed after macrorestriction with SmaI, according to the procedure of Tenover et al. 33 and analyzed with Bionumerics v6.5 (Applied Maths) using the unweighted-pair group method with arithmetic mean based on Dice coefficients, where optimization and tolerance were set to 0.5% and 1.25%, respectively. Isolates were considered to be of the same pulsotype if they shared at least 80% similarity on the band pattern.
Sequence type (ST) was determined using the multilocus sequence typing (MLST) scheme,20,29 in which the sequences of the seven housekeeping genes were compared to those available on the MLST database (www.mlst.net/).
PCR for repA genes
Families of plasmids present on the isolates were determined using the primers described by Jensen et al. targeting the repA gene, which is specific for each family. 21
Location of vanA genes on plasmid DNA
The plasmid content of all isolates was determined as described by Rosvoll et al. 28 Briefly, total DNA in agarose plugs was treated with S1 nuclease to linearize the plasmids and then were separated by PFGE. The gel was capillary blotted for 24 hours on a positively charged nylon membrane, fixed by UV irradiation, and then hybridized overnight at 42°C with a vanA probe generated by PCR and labeled with the ECL Direct Labeling and Detection System (GE Healthcare). The analysis was performed based on the size of plasmids containing vanA.
Determination of the rep-family of vanA plasmids
Some of the membranes used for location of vanA on plasmids were used to determine which rep-family contained the vanA plasmid. Isolates tested for rep hybridization were selected trying to cover all pulsotypes and sizes of vanA plasmids. E. faecalis isolates chosen were VRE15, VRE18, and VRE57. E. faecium isolates chosen were VRE13, VRE14, VRE16, VRE17, VRE44, VRE45, VRE46, VRE47, VRE49, VRE50, VRE58, VRE59, VRE60, VRE61, and VRE62. They were hybridized overnight at 55°C with different rep probes (rep1, rep2, rep3, rep9, rep14, and rep17) generated by PCR and labeled with Alkphos Direct Labeling Reagents (GE Healthcare). The detection was made using the CDP-Star Detection Reagent (GE Healthcare). The rep-family of the vanA-containing plasmid was determined visualizing where the hybridization occurred on the same linearized plasmid for vanA and a rep probe.
Results
Species identification and Tn1546 characterization
Forty-seven VRE isolates were identified as E. faecium (74.6%), 14 as E. faecalis (22.2%), 1 as E. gallinarum (1.6%), and 1 as E. raffinosus (1.6%). E. faecalis and E. faecium were selected for the experiments, amounting to 61 samples.
All of the isolates presented the vanA gene, confirming Tn1546 as the transposon involved in vancomycin resistance.
All E. faecalis, but VRE15, had an intact transposon. The overlapping PCR showed an atypical pattern within primers P17 and P18, from upstream of vanY to part of vanZ on VRE15 (Fig. 1). In this region, there was amplification of two fragments; one of them was 1.5 kb larger and the other about 1 kb smaller than expected.

Variations of Tn1546 found on vancomycin-resistant enterococci (VRE) isolates of the study.
E. faecium isolates did not show amplification of any fragment during the long PCR protocol. Overlapping PCR showed a deletion in the P1-P2 region in all E. faecium isolates, interfering with the whole transposon amplification. An insertion of ∼1.5 kb was also present within the vanSH region (P11-P12); IS1251 was the insertion within it. In addition, 46 of 47 E. faecium isolates showed a fragment 1,000 bp larger than expected within the primers P15-P16 region running from vanA to upstream of vanY genes (Fig. 1). IS1216E was the insertion within this region, which starts after the nucleotide number 8649 of the Tn1546 (Genbank M97297.1). E. faecium VRE46 showed no amplification in the regions of primers P15-P16, P17-P18, and P19-P1, covering part of vanA and the entire vanXYZ (Fig. 1). E. faecium VRE31 also amplified a fragment of ∼2,300 bp in the region of primers P9-P10. Sequencing of the P9 and P10 amplicon of VRE31 showed an insertion of 1,061 bp at position 4661 of Tn1546 (Genbank M97297.1), which refers to the beginning of the vanS gene. After the insertion, the last 4 bases of vanS (AAAT) were repeated and the sequence remained as a regular vanS gene. This four-base sequence is a direct repeat, characteristic of insertion elements. Although no specific insertion sequence was assigned to this region, an open reading frame was found from the nucleotide 88–1053 encoding an integrase of 322 amino acids identified as the IS30 family member. The amino acid sequence of this integrase had 76% identity with IS1062 and 72% identity with IS1252 on the insertion sequences database (www-is.biotoul.fr/).
Virulence factors and IS16
The profile agg+elrA + gelE + was present in 13 E. faecalis isolates. VRE38 was the only isolate with a different profile, cyl+agg+elrA+gelE+. The phenotypic test of VRE38 revealed beta-hemolysis on blood agar, confirming the presence of active cytolysin.
The predominant virulence profile among E. faecium was esp+acm+, in 38 isolates. Eight isolates had the profile acm+ and one had esp+acm+agg+.
The phenotypic test for gelatinase confirmed that all E. faecalis and no E. faecium isolates produced this virulence factor.
All E. faecium isolates were positive for the hospital marker IS16.
Antimicrobial susceptibility testing
All E. faecalis isolates had high levels of vancomycin and erythromycin resistance, with MICs >256 μg/ml. They were also resistant to quinupristin–dalfopristin and tetracycline, and susceptible to ampicillin, penicillin, daptomycin, linezolid, and tigecycline. The MIC of teicoplanin for VRE15 was 16 μg/ml, classifying it as intermediate-level resistant according to CLSI. 8
All E. faecium isolates had high levels of erythromycin, ampicillin, and penicillin resistance, with MIC >256 μg/ml. They were susceptible to daptomycin, tigecycline, linezolid, and tetracycline. VRE46 was susceptible to vancomycin and teicoplanin, with MIC=1.5 μg/ml, while all other E. faecium were highly resistant to these antibiotics. The 47 E. faecium had MICs between 1.5 and 2 μg/ml to quinupristin–dalfopristin, which classified them as intermediate-level resistant to this antimicrobial agent according to CLSI. 8
All E. faecalis isolates showed high-level gentamicin resistance and were susceptible to high levels of streptomycin. All E. faecium isolates also showed high-level streptomycin resistance and were susceptible to high levels of gentamicin.
For each drug, MIC50, MIC90, and the range of the MICs were calculated and all the isolates and the results are presented in Table 2.
MIC, minimal inhibitory concentration.
Molecular typing and repA genes
Thirteen of 14 E. faecalis isolates demonstrated a clonal pattern on PFGE referred to as pulsotype A1 (Fig. 2). Only VRE15, isolated from an infection site, showed a slightly different profile that was still related (six bands difference) to that clone. Five E. faecalis isolates (VRE2, 4, 15, 29, 53) randomly selected for an MLST experiment were of ST103.
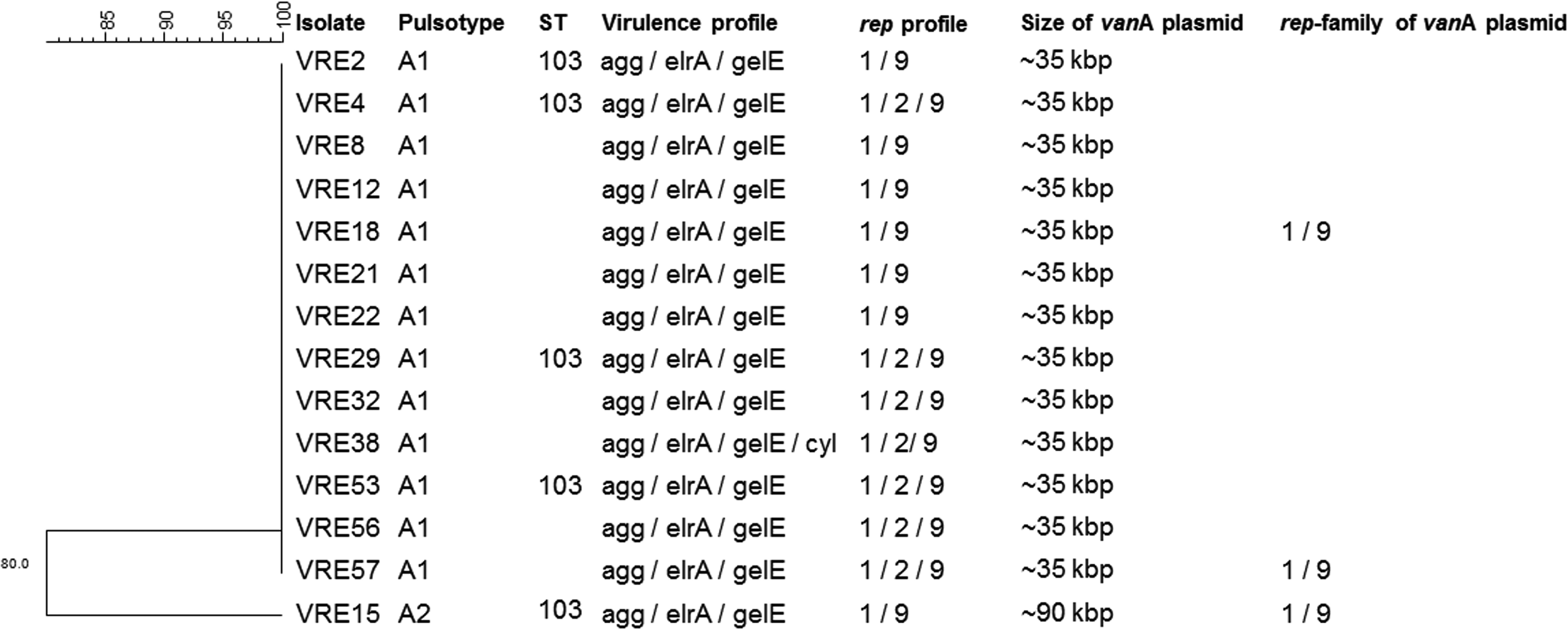
Dendrogram of Enterococcus faecalis isolates showing their sequence type (ST), pulsotype, virulence profile, rep-family profile and size, and rep-family of the vanA-containing plasmids.
E. faecium isolates were separated into four pulsotypes (A–D), as seen in Fig. 3, with predominance of pulsotype A and its subtypes (A1–A14), which harbored 35 isolates (74.47%). Pulsotype B and its four subtypes had nine isolates, pulsotype C had one isolate, and pulsotype D had two isolates. MLST of strains belonging to the four different pulsotypes was performed and they were of ST412, belonging to the Clonal Complex 17 (CC17).

Dendrogram of Enterococcus faecium isolates showing their ST, pulsotype, virulence profile, rep-family profile and size, and rep-family of the vanA-containing plasmids.
All E. faecalis isolates were positive for rep-families rep1 and rep9 and some isolates for rep2 (VRE4, 29, 32, 38, 53, 56, 57, and 63), while all E. faecium amplified for rep-families rep2, rep3, rep17, and only some isolates for rep14 (VRE5, 6, 7, 9, 10, 11, 14, 20, and 24).
Location of vanA genes in plasmid DNA and determination of the rep-family of the vanA plasmids
Figures 2 and 3 show the size of the plasmid containing vanA in each isolate as well as its rep-family, when tested.
E. faecium VRE37 is the only isolate that did not hybridize any plasmid with the vanA probe. Tn1546 might either be on the chromosome or on a plasmid greater than 250 kb.
E. faecium VRE61 has two plasmids containing vanA—one of ∼35 kb and another of ∼60 kb.
Figures 2 and 3 illustrate the peculiarities of each isolate selected for rep hybridization.
All E. faecium tested hybridized with rep17 probe, all but VRE45 and VRE47 (the only isolates tested had vanA plasmid of 40 kb; the others were of 35, 60, or 75 kb) hybridized with rep2 probe, and none hybridized with rep14 or rep3 probes.
All E. faecalis tested hybridized with both rep1 and rep9 probes.
Discussion
Most of the VRE in this surveillance program were identified as E. faecium. The 47 E. faecium were isolated from 42 different patients, while 14 E. faecalis were isolated from 13 patients. Four patients showed colonization by both species during hospitalization. A dissemination of an E. faecalis clone was detected by PFGE and MLST.
All E. faecalis isolates have elrA, a gene that codifies an internaline-like protein important for infection of macrophages. 3 They also present gelE and aggregation substance genes. Gelatinase is a metalloprotein described as the virulence factor for being also important for infections on a mouse peritonitis model. 32 Aggregation substances promote cell aggregation for subsequent occurrence of plasmid conjugation and also contribute to biofilm formation.7,13
The genes esp and acm were present in most E. faecium isolates. They are known to contribute to biofilm formation and colonization of the host tissue, respectively. A recent study of VRE in Brazil detected acm, esp, and hyl genes in E. faecium. 26 In the present study, the hyl gene was not found.
It is important to highlight here that the 47 E. faecium were clustered in only four pulsotypes and most of them (35 isolates) were of pulsotype A all sharing the ST412, which belongs to the hospital-adapted CC17. The presence of IS16, a marker of hospital clone and CC17, was found in all E. faecium. 21 All isolates were susceptible to the most recent antimicrobials available in Brazil: daptomycin, linezolid, and tigecycline, despite all E. faecalis being resistant to tetracycline.
The structure of Tn1546 found on all E. faecium isolates of this study had specific markers, including IS elements and deletion of the left-side inverted repeat (IRL). For example, IS1251 was the insertion within the vanS-vanH region at the same position as already observed in other Brazilian isolates.4,26,31 On the other hand, the transposon found in E. faecalis isolates is mostly intact, except for one isolate.
As E. faecium Tn1546 was lacking a repeated region, which is important for its mobility, it is likely the transposon needs a plasmid to move from one lineage to another.
E. faecalis VRE15 showed amplification of a nonexpected fragment and also a deletion near vanY, which culminated in the decreased MIC of teicoplanin. This rearrangement will be explored by our group in a future study.
E. faecium VRE46 showed a deletion on the transposon right-side inverted repeat (IRR) affecting the amplification of the P15 downstream region, including the vanA gene. Therefore, although it was selected by the hospital as VRE because it had MIC in the resistance level, it probably suffered reorganization in Tn1546 while stored at the laboratory and became susceptible to vancomycin.
S1 analysis revealed that all strains harbored one to six plasmids ranging in size from <9 to >200 kb. Positive vanA hybridization (Fig. 4) was detected on plasmid bands ranging from ∼25 to ∼75 kb in size for E. faecium and on plasmids of 35 kb in all E. faecalis, but one. VRE15 harbored a vanA plasmid of 90 kb. In previous studies, vanA plasmid of 70 kb was found in both species of VRE from hospitals of several states in Brazil, suggesting horizontal transfer, what differs from the results of this study.25,35

Pulsed-field gel electrophoresis (PFGE) of S1 nuclease-digested DNA of some VR E. faecium isolates visualized after staining with SYBR® Safe (Life Technologies)
Regarding the plasmid family content, we checked whether each species had different rep genes or not. This was the first report of rep-genes of VRE plasmids from Brazil. The rep2 family was the dominant rep-family in an enterococci collection analyzed by Jensen et al. when standardizing the PCR for repA. 21 We found this family in all E. faecium isolates of Risoleta Tolentino Neves Hospital and in eight E. faecalis. Although rep3 has never been found on any enterococci up to now, we found it on all E. faecium. The plasmids belonging to rep14 have only been described in the literature as present in E. faecium, which was confirmed on this study. 21 The rep-family that was found only on E. faecium is rep17, the same family that Jensen et al. observed in tetracycline-resistant E. faecium strains 21 ; they noted that pRUM, a member of this family, encodes multiresistance to antimicrobials, but needs further investigation. On our study, only the E. faecalis isolates were resistant to tetracycline, but they did not harbor rep17 plasmids. Among E. faecalis isolates, we found rep1, known as Inc18 plasmids and are important in the dissemination of resistance, and rep9, consistent with the results of Jensen et al., who did not find this family in E. faecium. These sex-pheromone response plasmids are known to be present in E. faecalis and to play an important role in transmitting antimicrobial resistance. 21 Another study has demonstrated that variants of Inc18, pAD1, and pRUM can influence the selection of specific clones within human adapted lineages. 16
By means of cohybridization experiments, we found that most E. faecium isolates tested carried a plasmid of the rep2 and rep17 family containing the vanA gene; these plasmids were ∼35, 60, or 75 kb. Only the plasmids of ∼40 kb carrying vanA were of the rep17 family exclusively. The isolate VRE61 carried two plasmids containing vanA, one of ∼35 kb and another of ∼60 kb, but both belonging to the rep2 family. VRE37 did not hybridize with the vanA probe, suggesting that the vanA-containing transposon might be in the chromosome (Fig. 3).
Even though only VRE15 had a different pulsotype and size of plasmid containing vanA, all E. faecalis isolates hybridized with both rep1 and rep9 probes. It seems that on both E. faecalis and E. faecium, the vanA-containing plasmids are mosaic, since most times they hybridized with more than one rep-family probe.
A study with 104 VRE from Portugal found that the plasmids carrying vanA in E. faecium could be grouped as pRUM derivatives (rep17, rep2, rep1), Inc18 (rep1), or mosaics of Inc18-pRUM-like (rep2, rep1, rep17), while for E. faecalis, vanA plasmids were Inc-18-pheromone-responsive mosaics. This is an important fact to keep in mind that besides the promiscuity of plasmids carrying Tn1546, they also show the ability to shuffle. Thus, it is very difficult to classify them. 16
Despite the fact that there was simultaneous colonization by more than one species in some patients during hospitalization, which would have facilitated the transmission of resistance genes and virulence factors by horizontally transferred plasmids, 19 putting all our results together, it is unlikely that there was an exchange of VanA elements between species. Therefore, we suggest there were at least two different Tn1546 origins in this hospital: one in E. faecalis and another in E. faecium.
Footnotes
Acknowledgments
The authors thank Edna Leite, Hoberdan Pereira, and Hyllo Marcello Jr. from RTNH for providing the isolates, Lars Bogø Jensen for providing the rep-family plasmids controls, Ana Lúcia Darini for allowing the use of LEBEM and providing controls for species identification, Van element typing, and detection of virulence factors, and Hernan Fazzio, Vicente Mattos, and Caio Pontes for contributing with PCRs, IS sequencing, and MLST. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (472907/2010-7) and Pfizer Laboratories LTDA and by TPM and ANGD scholarships (Fundação de Amparo à Pesquisa do Estado de São Paulo 2011/14592-1 and 2010/11890-9, respectively).
Disclosure Statement
No competing financial interests exist.