
Abstract
Resistance mechanisms in a group of carbapenemase-negative Pseudomonas aeruginosa that were susceptible to all antibiotics except carbapenems (carbapenem resistance-only P. aeruginosa [CROPA]) were studied. Ten genetically nonrelated CROPA isolates and their carbapenem-susceptible counterparts were further investigated. OprD production was demonstrated by protein electrophoresis in only 1 of the 10 carbapenem-susceptible isolates, while the other 9 isolates showed hyperproduction of OprM. DNA sequencing of oprD revealed a shortened loop 7 domain (group 1A allele) in eight carbapenem-susceptible isolates. Various oprD mutations, leading to early terminations, were found in 9 of the 10 CROPA isolates. RNA analysis demonstrated hyperexpression of oprM with normal expression of mexA in eight of the carbapenem-susceptible isolates, while in seven of their CROPA counterparts, the oprM expression was significantly reduced. Deletion of oprM was performed in two pairs of representative isolates. Selection of imipenem resistant variants by a disc assay indicated that the lost-of-function mutations in oprD occurred relatively faster in the ΔoprM mutants compared with their corresponding parent strains. Under selection pressure, reduced production of OprM may promote the selection of spontaneous changes in oprD, resulting in the carbapenem resistance in a group of pan-susceptible P. aeruginosa isolates characterized by harboring an oprD-group 1A allele.
Introduction
P
Chromosomally encoded resistance in P. aeruginosa is predominantly achieved by reducing the expression of outer membrane porins, overproducing efflux pumps, and presumably increasing the production of drug-inactivating enzymes. 13 It has been shown that porin OprD in P. aeruginosa serves as the preferred means of entry for carbapenems; thus, reduced OprD expression should lead to carbapenem resistance.13,14,21 We also have demonstrated that the loss of OprD is the major contributor to carbapenem resistance among XDRPA clinical isolates. 19
The reduction of antibiotic accumulation can also be achieved through the overexpression of membrane-associated efflux pumps. Among the five efflux pump superfamilies, the resistance-nodulation-division (RND) family plays a very important role in the expression of drug resistance in P. aeruginosa. RND pumps are typically composed of a tripartite system that includes a periplasmic membrane fusion protein (MFP), an outer membrane factor (OMF), and a cytoplasmic membrane (RND) transporter. 13 In addition, the genes encoding these pump components are frequently organized as operons in the P. aeruginosa genome. For example, the resistance to the carbapenem meropenem but not imipenem is associated with the overexpression of the mexA (encoding MFP)-mexB (encoding RND)-oprM (encoding OMF) operon.14,15
At Chang Gung Memorial Hospital, Linkou (CGMH, Linkou, Taiwan), clinical physicians and infection control personnel noted that the recovery of P. aeruginosa strains resistant only to carbapenems (carbapenem resistance-only P. aeruginosa [CROPA]) has been increasing in recent years. The isolates in this group were susceptible to all antibiotics, except the carbapenems that are commonly used in treating P. aeruginosa infections. However, no solid evidence for the explanation of this carbapenem resistance among the clinical isolates has been reported to date. In this study, the mechanisms underlying this observation were investigated based on the results from a matched-pair study. We show that OprM reduction in P. aeruginosa carrying a shortened oprD loop 7 (L7) domain may promote the imipenem resistance. The DNA sequence of the shortened oprD L7 region was completely matched to the oprD-group 1A allele. 22
Materials and Methods
Ethics statement
This study aimed at characterizing P. aeruginosa isolates using molecular methods. All isolates studied were retrieved retrospectively from the Bacteria Bank, Department of Laboratory Medicine, CGMH, Linkou. The clinical information of the patients was neither available nor required in this study. The patient's informed consent was not required or collected, because all microbial cultures were ordered by physicians due to the necessity of clinical management (none were collected purposely for this study). The design and procedure of the study had been approved by the Institutional Review Board of the CGMH, Linkou.
Settings and bacterial strains
CGMH at Linkou is a university-affiliated medical center with 3,715 beds and is located in northern Taiwan. From 1999 to 2011, a total of 46,281 P. aeruginosa clinical isolates were identified by standard methods at this medical center, and their susceptibilities to antimicrobial agents were determined by a routine disc diffusion method according to CLSI guidelines. 2 Isolates used for the further analyses are summarized in Table 1. The CROPA strains were defined as isolates that were susceptible to all antimicrobial agents tested (amikacin, gentamicin, ceftazidime, aztreonam, ciprofloxacin, cefepime, piperacillin, and piperacillin-tazobactam) but not carbapenems (meropenem and imipenem). Susceptibilities to different carbapenems (imipenem, meropenem, and doripenem) in paired isolates were interpreted by determining the minimum inhibitory concentrations (MICs) by E-test according to CLSI guidelines. 2
CROPA, carbapenem resistance-only P. aeruginosa; XDRPA, extensive drug-resistant P. aeruginosa.
Genomic fingerprinting
Genomic fingerprinting was performed by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). A PFGE analysis of SpeI-digested DNA fragments was performed as described elsewhere. 20 The 10 pair isolates were further subjected to an MLST assay by sequencing seven housekeeping genes according to methods previously described. 4 The allelic profiles for each gene and the subsequent sequence types were determined by referring to the MLST database (http://pubmlst.org/paeruginosa/).
Polymerase chain reaction amplification and DNA sequencing
DNA extraction was performed using the QIAamp DNA Mini Kit (Qiagen) according to the manufacturer's instructions. Polymerase chain reaction (PCR) was used to detect the existence of known acquired genes encoding carbapenem resistance-associated β-lactamases: Ambler class A carbapenemases, such as KPC; class D OXA-type extended spectrum β-lactamases, such as OXA-23, -24, and -48; and class D metallo-β-lactamases, such as VIM, IMP, NDM-1, SIM, GIM, and SPM.3,17 To investigate the existence or integrity of oprD, the primer set OprD-F (5′-CGCCGACAAGAAGAACTAGC-3′) and OprD-R (5′-GTCGATTACAGGATCGACAG-3′) was employed for the PCR analyses. An additional primer, OprD-F2 (5′-GCCGACCACCGTCAAATCG-3′), combined with OprD-R was specifically used for checking a shortening of the L7 region within oprD.
Quantitative real-time reverse transcriptase-PCR
The expression levels of the efflux gene mexA, oprM, and porin oprD were determined by quantitative real-time reverse transcriptase-PCR (qRT-PCR) according to previously described methods.5,19 All amplifications were performed in triplicate. The level of mRNA was normalized to that of the housekeeping gene rpsL using the Pfaffl equation. 16 The results are presented as fold-change expression levels relative to the wild-type strain PAO1. An effect on gene expression was considered significant when the ratio was ≥3-fold for mexA and oprM or <0.4-fold for oprD.
Analysis of outer membrane proteins
The extraction of outer membrane proteins was performed according to methods described elsewhere, with minor modifications. 11 Briefly, cells from an overnight culture in LB broth were harvested by centrifugation at 1,500 g for 30 min at 4°C. The cell pellets were suspended in suspension buffer (30 mM Tris/HCl [pH 8.0]) supplied with protease inhibitor phenylmethylsulfonyl fluoride (PMSF), followed by sonication for 2 min on ice to break the cells. The unbroken cells were centrifugated at 4°C, and the membranes in the supernatant were further pelleted at 100,000 g for 1 hr at 4°C and suspended in the same buffer. The inner membrane was solubilized by the addition of sodium N-lauroylsarcosinate to a final concentration of 1% and incubation for 30 min at 30°C. The remaining outer membrane was pelleted at 40,000 g for 40 min at 4°C and suspended in suspension buffer before 0% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis. The Coomassie blue-stained bands of interest were further identified by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) at the Clinical Proteomics Core Laboratory, CGMH, Linkou.
Deletion of oprM in P. aeruginosa
The deletion of oprM was performed by introducing deletion-carrying pEX18Tc into P. aeruginosa strains of interest according to methods described elsewhere and approved by the bio-safety committee, Chang Gung University.
7
The plasmid construct was prepared by cloning PCR-amplified DNA fragments corresponding to the region upstream of oprM and the downstream portion of this gene using P. aeruginosa PAO1 genomic DNA as the template. Briefly, the 935-bp upstream arm terminating at the 63rd nucleotide of oprM was amplified using the primer pair OprM-U-F (5′-CGTCAAC
Selection of imipenem resistance in P. aeruginosa by the disc diffusion method
The selection of inducible imipenem resistance was performed by placing an imipenem disc on a Mueller–Hinton agar plate inoculated with the indicated P. aeruginosa isolates. The bacterial cells growing near the edge of the inhibitory zone or the colonies grown within the zone were collected for further passaging and genetic analyses.
Results
Molecular epidemiology and mechanisms of carbapenem resistance in CROPA isolates
A total of 1,704 (3.7%) of 46,281 P. aeruginosa clinical isolates were found to be CROPA strains. The yearly prevalence of CROPA is shown in Figure 1. A significant increase in CROPA was noted from 1.8% in 1999 to 3.5% by 2003 (p<0.00005), which was followed by an obvious but not significant drop to 2.7% by 2004 after an outbreak of SARS in Taiwan. However, the prevalence had significantly increased again to 4.9% by 2008 (p<0.000001). Of the 1,704 CROPA isolates, 80 nonrepeated clinical isolates were further selected for the investigation of their epidemiological correlation by PFGE. Seventy-four pulsotypes were identified, indicating that there was no CROPA clonal spread.
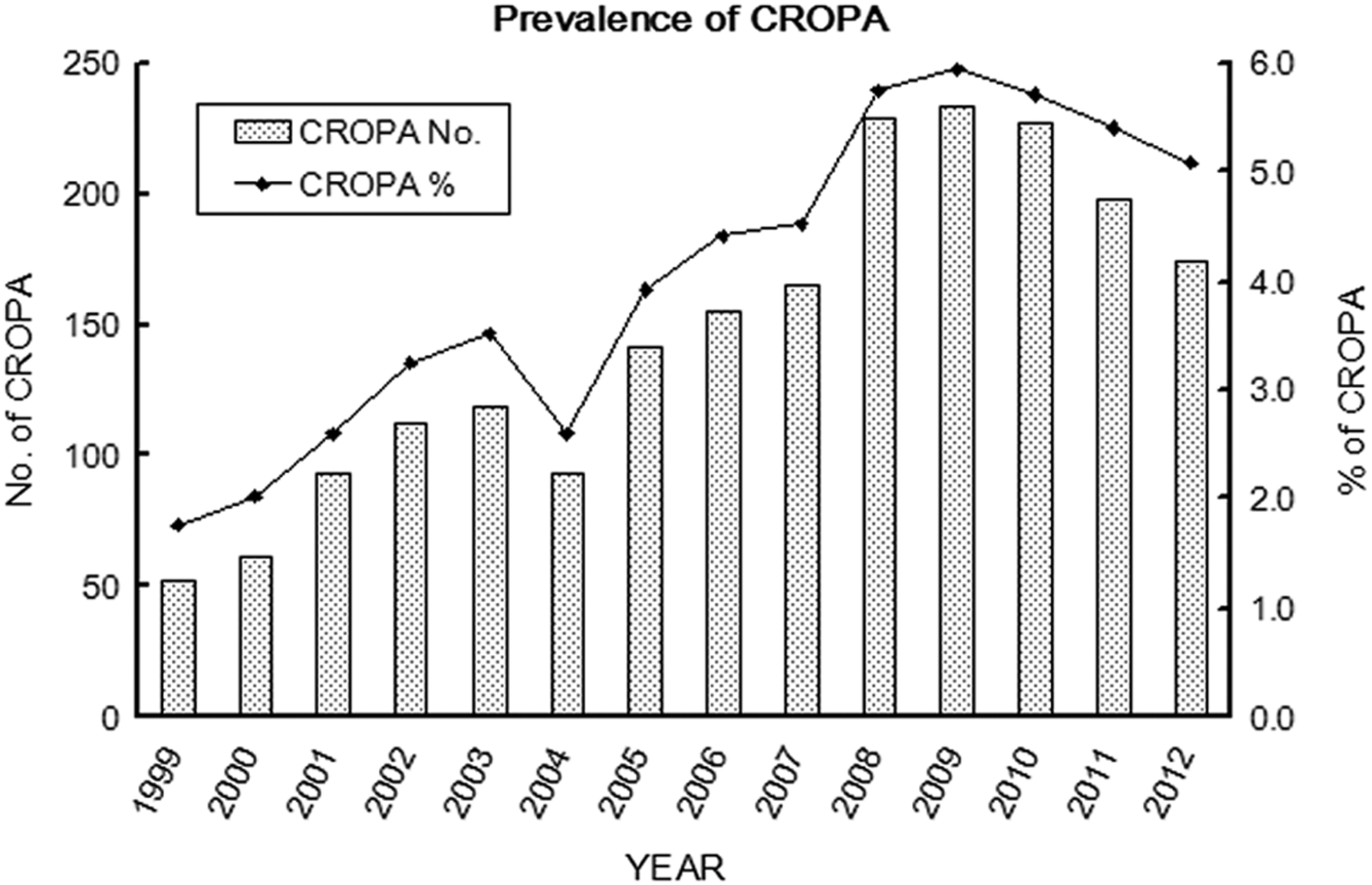
Prevalence of carbapenem resistance-only Pseudomonas aeruginosa (CROPA) isolates. Secular trends in the annual number (histograms) and prevalence (lines) of CROPA isolates identified between 1999 and 2012 at CGMH, Linkou.
Of the 80 isolates, 75 were imipenem nonsusceptible, and the remaining 5 were both meropenem and doripenem nonsusceptible. No acquired resistance determinants were found in any of the isolates. The expression level of oprD was then investigated, with the qRT-PCR results showing reduced oprD expression in only 33 isolates (data not shown). This finding suggested that the reduced expression of oprD may not be the major contributor to carbapenem resistance among the 80 isolates. Other possible alternative mechanisms may include impaired assembly of the OprD porin or increased expression of efflux pumps responsible for carbapenem resistance. Both possibilities were further subsequently studied.
Investigation of the mechanisms of carbapenem resistance through matched-pair study
Our investigation of the genetic mechanisms of carbapenem resistance was then limited to analyses of single pairs of isolates collected from individual patients. The results shown in Table 2 indicate that all of the resistant strains showed a high level of resistance to imipenem but a low level of resistance or susceptibility to meropenem and doripenem, except for strains 23R and 33R. The isolates in each single pair showed the same MLST (Table 2) and PFGE patterns (data not shown), suggesting that they are isogenic strains. However, a great diversity of MLSTs and PFGE patterns was found pair by pair, indicating that there was no genetic relatedness of the isolates in different pairs.
ET/FS, early termination/frame shift; L7, gene shortage in the oprD loop 7 domain; Y, intact loop 7 domain with various polymorphisms in other domains of oprD.
Relative to expression in PAO1.
MIC, minimum inhibitory concentration; MLST, multilocus sequence typing.
Since the expression level of oprD was discordant among the 20 paired isolates (Table 2), oprD was sequenced in each isolate. In comparison to the strain PAO1 genome, one point mutation and replacement of 12 amino acids by a different sequence of 10 amino-acid residues were found in the oprD L7 domain in all of the carbapenem-susceptible isolates, except for strains 24S and 34S (Table 2 and Fig. 2). The DNA sequence of the unique oprD L7 region completely resembled the oprD-group 1A allele previously reported by Turton et al. (GenBank accession No. GQ228105.1). 22 Several common amino-acid polymorphisms unique to these oprD-group 1A allele-carrying isolates were also found in other loop domains (Fig. 2). With regard to CROPA isolates, all but strain 9R had further mutational changes leading to early termination and/or frame shifts due to insertion or deletion in the middle region of the oprD gene. Such loss-of-function changes were also found in strains 23R and 33R even though they both carried an intact L7 domain (Table 2). Although both strains 24S and 34S, the susceptible counterparts of strains 23R and 33R, respectively, had an intact L7 domain, they demonstrated common mutations in loop 2 (T103S and K115T) and loop 3 (F170L) domains; such mutations were unique to PAO1 or other eight susceptible P. aeruginosa isolates that carried the oprD-group 1A allele (Fig. 2).

Multiple sequence alignment of OprD protein variants from PAO1 and 10 susceptible isolates in matched-pair study. Locations of putative loops are highlighted by gray shading. Positions of the amino-acid substitution only found in both strains 24S and 34S are shown in bold. Star marks indicate changes in amino acid only found in isolates carrying oprD-group 1A allele. Sporadic amino-acid substitutions differing from PAO1 are shown in black shading.
The results of the outer membrane protein assay showed only trace or invisible OprD on the gels after Coomassie blue staining for all the susceptible isolates, except for strain 34S (Fig. 3). OprD expression was not observed among any of the carbapenem-resistant isolates. Interestingly, hyperexpression of an outer membrane protein was found in all of the susceptible strains, except for strain 34S, whereas the expression of this protein was either lost or weak in all of the resistant strains (Fig. 3). This SDS gel protein band was further analyzed by MALDI-TOF MS, and OprM was identified. The expression levels of mexA and oprM were then evaluated by qRT-PCR. However, no significantly increased mexA expression (>3-fold change relative to PAO1) was found in any strain, except 23R. In addition, a stronger oprM expression was found in the susceptible versus resistant strains among 8 of the 10 pairs (ranging from 1.6- to 491-fold), except for the pair strains 15R/16S and 27R/28S (Table 2). Nonetheless, referring to the SDS-PAGE results shown in Figure 3, both the 15R and 27R strains revealed a markedly weaker OprM expression than their susceptible counterparts.
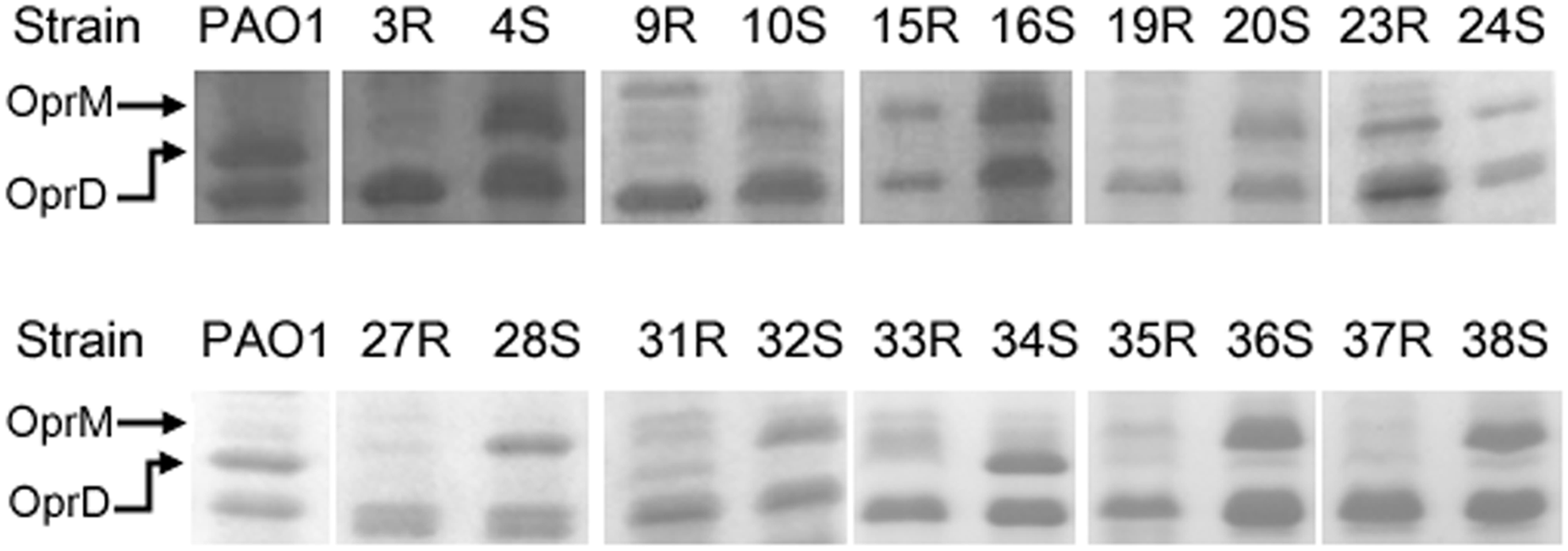
Outer membrane protein analysis. Expression of the outer membrane protein of strains used in matched-pair study was analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and PAO1 was used as a control strain. The protein bands in the position corresponding to OprD and OprM were confirmed by matrix-assisted laser desorption ionization-time of flight mass spectrometry.
Investigation of the role of OprM in carbapenem resistance among CROPA isolates
The hyperexpression of OprM in the susceptible isolates but reduced to trace level expression in the CROPA isolates suggested a nonefflux porin function of OprM in this case. Therefore, oprM in strains 19R, 20S, 23R, and 24S, along with PAO1, was knocked out, and their susceptibility to different carbapenems was determined by the E-test. The results shown in Table 3 indicate that the deletion of oprM further increased the susceptibility of all of the mutant strains to both meropenem and doripenem, implying an efflux porin function associated with the MexAB efflux pump system. 15 Susceptibility testing of the 10 antimicrobial agents (see Materials and Methods section) by disc diffusion was also investigated. The deletion of oprM increased the susceptibility to almost all of the antibiotics tested in the susceptible strains, confirming its efflux porin role associated with different RND pumps. In addition, DNA sequencing of the oprM gene and its upstream region was performed, and no mutation was found in any of the 20 isolates used in the matched-pair study (data not shown).
S, susceptible; R, resistant; AN, amikacin; GM, gentamicin; CAZ, ceftazidime; ATM, aztreonam; CIP, ciprofloxacin; FEP, cefepime; PIP, piperacillin; TZP, piperacillin-tazobactam; MEM, meropenem; IPM, imipenem.
To determine whether the reduced OprM expression in the susceptible strains contributed to the development of imipenem resistance, strains PAO1, 20S (oprD-group 1A allele), 24S (OprD polymorphisms), and their oprM deletion counterparts were plated onto agar plates under imipenem disc antibiotic pressure to select for resistant mutants. Amazingly, the deletion of oprM in strains 20S and 24S rapidly promoted the selection of spontaneous mutations to imipenem resistance after three passages under the selection pressure (Table 4). Continuous imipenem selection pressure did not drive the intact oprM-carrying carbapenem-susceptible isolate 24S to become imipenem resistant until the fifth passage, while none of the strains PAO1, PAO1ΔoprM, and 20S developed carbapenem resistance during the period of this study (11 passages). Moreover, a decreasing expression of oprM to a lower level concomitant with the development of imipenem resistance was observed in strain 24S, whereas the expression of oprM stood in a relatively higher level in strain 20S. Early termination due to a premature stop codon and/or frame shift mutations in oprD were also observed in the tested strains showing induced carbapenem resistance (Table 4). Two additional susceptible isolates carrying oprD-group 1A allele were employed to investigate whether the original oprM expression (relative to PAO1) affects the rate in developing imipenem resistance. The higher oprM expression in strain 4S (13-fold) displayed a slower rate of resistant development than that in strain 10S (5-fold; Table 4).
Zone diameters were the same during the passages between 9 and 11.
In mm.
ET, early termination.
Proportion of CROPA isolates carrying oprD-group 1A allele
Our results suggested that a shortening of the L7 region, followed by loss-of-function mutations in the oprD gene, was the major contributor of carbapenem resistance among the CROPA isolates. Therefore, the proportion of isolates carrying oprD-group 1A allele was determined by DNA sequencing among the collected 102 CROPA clinical isolates. Results shown in Figure 4 indicate that 75 (73.5%) of the 102 CROPA isolates carried oprD-group 1A allele. An obvious but nonsignificant increase in the proportion of oprD-group 1A allele-carrying CROPA isolates was observed from 2006 (67%) to 2008 (84%; Fig. 4), which was correlated with an increase in the abundance of total CROPA isolates (Fig. 1). In addition, only 5 of 48 (10.4%) carbapenemase-positive XDRPA clinical isolates carried the oprD-group 1A allele, whereas such a gene shortening was found in 35 of 66 (53.0%) carbapenemase-negative XDRPA clinical isolates. In order to understand the proportion of isolates carrying the oprD-group 1A allele in the whole P. aeruginosa population, 178 clinical isolates collected from blood cultures were investigated and 73 (41.0%) of them carried the oprD-group 1A allele.
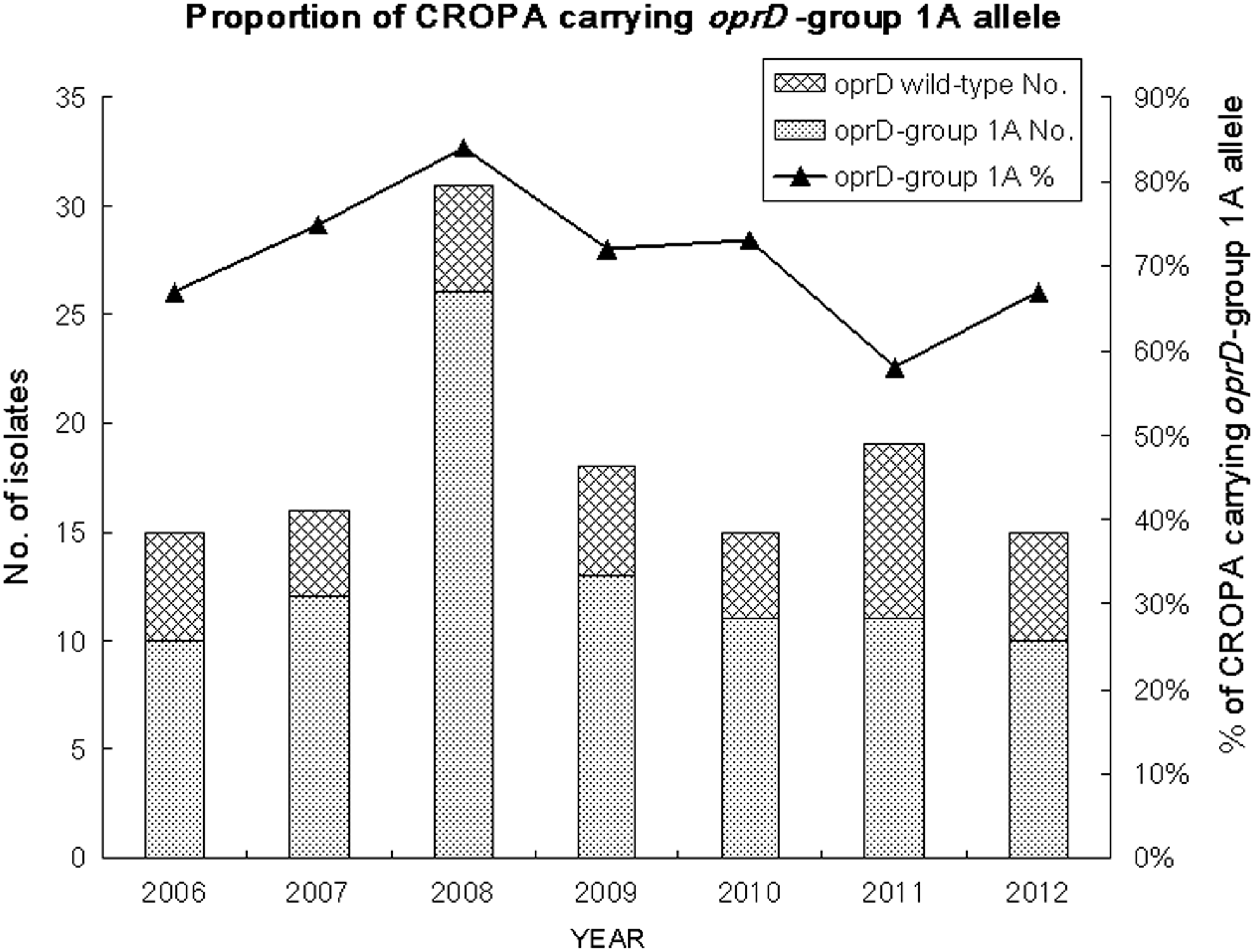
Proportion of CROPA isolates carrying oprD-group 1A allele. Secular trends in the annual proportion (lines) of CROPA isolates carrying the oprD-group 1A allele identified between 2006 and 2012 at CGMH, Linkou. The annual number of CROPA isolates with an intact oprD and oprD-group 1A allele is also shown in meshed and dot bars, respectively.
Discussion
Through the comparison with carbapenem-susceptible counterparts, the CROPA isolates were found to be characterized by harboring a deficient oprD with a unique shortening in the L7 domain (oprD-group 1A allele). We also demonstrated that, probably under carbapenem selection pressure, such isolates may further develop mutational changes, such as early termination and/or frame shift, in oprD, leading to function loss of the porin and carbapenem resistance, particularly imipenem. In addition, for the first time, reduced production of OprM was found to play a role in promoting the evolution of carbapenem resistance.
Eight of the 10 carbapenem-susceptible isolates in the matched-pair study carried a unique shortened oprD L7 domain, identical to the group 1A allele of an oprD sequence type previously reported by Turton et al. 22 It has been shown that deletion of short stretches of amino acids in L7 did not affect the OprD assembly into outer membranes, while it resulted in hypersusceptibility to both imipenem and meropenem. 10 The same construct also increased susceptibility to β-lactams, quinolones, and chloramphenicol. 9 Epp et al. detected a shortening in the oprD L7 region with concomitant reduced expression of OprD in seven P. aeruginosa clinical isolates with unusual meropenem hypersusceptibility and increased imipenem resistance. 6 Interestingly, the L7 regions in six of their isolates also belonged to the oprD-group 1A allele. In this study, 8 of the 10 carbapenem-susceptible strains that harbored the oprD-group 1A allele also demonstrated reduced expression of OprD. Although its expression was weak, such an altered porin may still allow the effective entry of carbapenems into the bacteria, as demonstrated in previous reports.6,9,10 Still, some common amino-acid polymorphisms were found in the other domains among isolates carrying the oprD-group1A allele or intact L7 regions (Fig. 2). The effect of these amino-acid substitutions on the carbapenem susceptibility remains to be further studied.
Through the analysis of carbapenem-resistant isolates, reduced expression of OprD is known to cause carbapenem resistance in P. aeruginosa.13,14,19,21 In the preliminary stage of this study, we were confused with the finding that on protein electrophoresis, most of the carbapenem-susceptible isolates did not seem to produce OprD. With the subsequent matched-pair study, we were able to trace the evolution of these oprD-group 1A allele-carrying isolates from pan-susceptible to the antibiotics tested that were resistant solely to carbapenems. Not surprisingly, the carbapenem resistance was achieved by the loss-of-function mutations, including early termination with a premature stop codon and/or frame shift mutations, in OprD, thereby totally blocked its production.
The production of OprM was high among the carbapenem-susceptible isolates carrying the oprD-group 1A allele, but was reduced in the CROPA isolates, suggesting that the reduced expression of OprM may play a role in the development of carbapenem resistance. Our observation appeared to be contradictory with the general understanding that meropenem resistance may be caused by the overexpression of membrane-associated efflux pumps, such as mexAB-oprM.15,18 It was also noted that mexA remained unchanged among the study isolates. Although mexA, mexB, and oprM are organized into an operon, a weak promoter was found upstream of oprM in the coding region of mexB, enabling a portion of the oprM expression to be independent of mexAB.13,24 However, DNA mutations were not found in the coding region and/or within 350 bp upstream of oprM (data not shown). Thus, the factors that triggered the overexpression of oprM independent of mexAB as observed in this study remain unclear. Nonetheless, Zhao et al. demonstrated that the introduction of oprM, but not mexAB, into a multidrug-susceptible ΔmexAB-oprM mutant increased the resistance to quinolones, cephalosporins, erythromycin, and tetracycline. 24 These authors also found that OprM can be expressed and function as a drug efflux independent of MexAB. 24 In agreement with their findings, the deletion of oprM in this study resulted in a further increase in susceptibility to almost all of the antibiotics tested, particularly in the susceptible strains. The results suggested that the OprM may act as an efflux porin alone or may link to another RND system.
Interestingly, deletion of oprM in the isolates carrying the oprD-group 1A allele apparently accelerated the occurrence of the loss-of-function mutations in oprD to within three to four passages under continuous imipenem selection pressure. In addition, the oprM expression reduced gradually before the occurrence of imipenem resistance in strain 24S; however, such loss-of-function mutations in OprD were not observed in strain 20S, probably due to the fact that the expression level of oprM remained relatively high. To our knowledge, this observation has never been reported earlier.
The resistance mechanism in the paired strains 9R and 10S is largely unexplainable. Both strains 9R and 10S carried the oprD-group 1A allele, but additional oprD mutations or a significant increase in mexA were not found in strain 9R. The factors contributing to the observed carbapenem resistance in strain 9R warrant a further study. Furthermore, although strains 24S and 34S carried an intact oprD L7 domain as PAO1, the hypersusceptibility to carbapenems and other antibiotics may be attributed to the amino-acid polymorphisms in the putative L2 (T103S, K115T) and L3 (F170L) domains according to the report by Epp et al. 6 Deletion of the associated regions also has been shown to increase imipenem MIC or diminish OprD expression. 12
In conclusion, using a matched-pair study, we demonstrate the evolution of resistance to carbapenems, particularly imipenem, in P. aeruginosa clinical isolates carrying the oprD-group 1A allele. We hypothesize the mechanism underlying the evolution as the following. First, isolates carrying the oprD-group 1A allele allow most of the tested antibiotic to pass efficiently into the cell through altered porins, thereby leading to a hypersusceptibility to most of the antibiotics. Second, stress due to the carbapenems may select for the variants with spontaneous reduced expression of OprD and/or with hyper-production of related efflux pumps in response to antibiotic pressure. However, these spontaneous changes do not allow the bacteria to effectively resist carbapenem attack. Third, the antibiotic pressure forces the bacteria to undergo loss-of-function mutations in oprD concomitant with suppressed OprM expression. The reason that the continued imipenem pressure diminished oprM expression in our experiments remains unclear. One possible explanation is that in the presence of hyper-susceptible strains with the oprD-group 1A allele, small inhibitory amounts of carbapenem could select OprM variants with low-level resistance. These variants, surviving in the presence of low levels of carbapemem, might give rise to effective oprD mutants with carbapenem resistance. In the presence of such an effective mechanism, OprM variants apparently have an unnecessary cost and may therefore revert to normal expression by compensatory mutations. Possible compensatory mutations may include those occurring in the promoter sequences of mexA and mexR or in the putative ribosome binding site located upstream of the mexAB oprM operon. 8 The proportion of the carriage of the oprD-group 1A allele among the CROPA clinical isolates was 73.5%, suggesting that such gene shortage plays an important role in the evolution of carbapenem resistance in response to selection pressure. This gene shortening was also found among 41.0% of the general P. aeruginosa isolates recovered from the blood. As a result, our study provides a warning of the potential risk when carbapenems are initially applied for treating infections caused by pan-susceptible P. aeruginosa, especially in isolates carrying the oprD-group 1A allele.
Footnotes
Acknowledgments
The authors thank the Clinical Proteomics Core Laboratory, Chang Gung Memorial Hospital, Linkou, for excellent technical assistance in MALDI-TOF MS. This study was supported by the Research Program of Chang Gung Memorial Hospital (grant number CMRPG3A0831∼2 and CMRPG3D0031), Taoyuan, Taiwan.
Disclosure Statement
All authors report no conflicts of interest relevant to this study.