
Abstract
The usefulness of carbapenems for gram-negative infections is becoming compromised by organisms harboring carbapenemases, enzymes which can hydrolyze the drug. Currently KPC (class A), NDM (class B), and OXA-48 types (class D) are the most globally widespread carbapenemases. However, among the GES-type class A extended-spectrum β-lactamases (ESBLs) there are variants that hydrolyze carbapenems, with blaGES-5 being the most common. Two Escherichia coli and two Serratia marcescens harboring blaGES-5 on plasmids were isolated by the Canadian Nosocomial Infection Surveillance Program (CNISP) from four different patients in a single hospital over a 2-year period. Complete sequencing of the blaGES-5 plasmids indicated that all four had nearly identical backbones consisting of genes for replication, partitioning, and stability, but contained variant accessory regions consisting of mobile elements and antimicrobial resistance genes. The plasmids were of a novel replicon type, but belonged to the MOBQ1 group based on relaxase sequences, and appeared to be mobilizable, but not self-transmissible. Considering the time periods of bacterial isolation, it would appear the blaGES-5 plasmid has persisted in an environmental niche for at least 2 years in the hospital. This has implications for infection control and clinical care when it is transferred to clinically relevant gram-negative organisms.
Introduction
W
The Canadian Nosocomial Infection Surveillance Program (CNISP) has conducted surveillance of carbapenem-resistant gram-negative organisms (CRGN) from a set of acute care hospitals in Canada (CRGN study) and data from the first year, which included Enterobacteriaceae, Pseudomonas, and Acinetobacter, have been published. 11 Carbapenemases were detected in 11% of eligible isolates (30/274) and included blaKPC-3 (n=7), blaNDM-1 (n=2), and blaSME-2 (n=1) in Enterobacteriaceae, blaOXA-23 in Acinetobacter baumannii (n=9), and blaVIM-2 (n=1), blaVIM-4 (n=7), and blaGES-5 (n=3) in Pseudomonas aeruginosa. In subsequent study years, which excluded Pseudomonas, only five organisms harboring a blaGES gene were identified, an A. baumannii harboring blaGES-11 (an ESBL), two Escherichia coli, and two Serratia marcescens harboring blaGES-5. Intriguingly, the four Enterobacteriaceae harboring blaGES-5 were isolated over a 2-year period from a single hospital, which prompted us to further explore the molecular epidemiology of the cases, including the complete sequence of the blaGES-5 plasmids.
Materials and Methods
Bacteria isolates and susceptibility testing
E. coli A4Y217 and A4Y413 and S. marcescens A4Y201 and A4Y426 along with associated patient information were isolated as part of the CNISP CRGN study. 11 Antimicrobial susceptibilities were determined by Vitek 2 (AST–N219) and Etest (bioMérieux) and interpreted using CLSI guidelines. 4
Plasmid isolation and transfer experiments
Plasmid DNA was purified using Plasmid Mini or Midi Kits (Qiagen). To determine if the blaGES-5 plasmid was transmissible, transfers were attempted by mixing donor culture (50 μl) and rifampin-resistant E. coli RG12 (200 μl) from overnight drug-free cultures in Luria-Bertani (LB) medium into 5 ml fresh media with growth overnight at 37°C without shaking. Selection for transconjugants was conducted by plating on the LB medium containing rifampin (150 mg/L), meropenem (0.1 mg/L), and ampicillin (50 mg/L). Plasmids harboring blaGES-5 were isolated by electrotransformation of whole plasmid DNA from clinical isolates into electrocompetent E. coli DH10B (Life Technologies, Inc.) with selection on meropenem and ampicillin.
Molecular typing and PCR
Pulsed-field gel electrophoresis (PFGE), after restriction with XbaI, was done using the PulseNet protocol for E. coli (www.cdc.gov/pulsenet/PDF/ecoli-shigella-salmonell-pfge-protocol-508c.pdf; last accessed July 8, 2014) and the fingerprints analyzed using BioNumerics v5.1 (Applied Maths, Inc., Sint-Martens-Latem, Belgium). Plasmid fingerprinting analysis was done after digestion with EcoRV, which was found to give suitable patterns (5–10 bands) in initial enzyme screening experiments. E. coli multilocus sequence typing (MLST) was carried out as described. 21 Multiplex PCR for blaTEM, blaSHV, blaCTX-M, blaOXA-1, and blaCMY-2 groups and multiplex PCR for blaKPC, blaOXA-48, blaNDM, blaIMP, blaVIM, and blaGES groups 11 and replicon typing PCR 3 were carried out as previously described. PCR to detect the repA gene of the blaGES-5 plasmids was conducted with repG5F, 5′-ACACAATCCGACCTCAAGCT, and repG5R, 5′-ATCAGACAACACCAACAGGC (534 bp product).
DNA sequencing and bioinformatics
Sequencing was conducted on purified plasmid DNA using 454 pyrosequencing (454-Genome Sequencer FLX; Roche Diagnostics) and the reads assembled de novo using Newbler v2.5.3. Contigs were joined by standard PCR/sequencing. Annotation was carried out using the Prokka prokaryotic genome annotation system (www.vicbioinformatics.com/software.prokka.shtml) and manual curation using the BLAST suite of programs (http://blast.ncbi.nlm.nih.gov/BLAST.cgi).
Accession numbers
Plasmids pG5A4Y427, pG5A4Y201, pG5A4Y413, pG5A4Y217, and blaCMY-111 have been assigned accession nos. KJ541068, KJ541069, KJ541070, KJ541071, and KJ155695, respectively, in the GenBank database.
Results
Initial bacterial characterization and case information
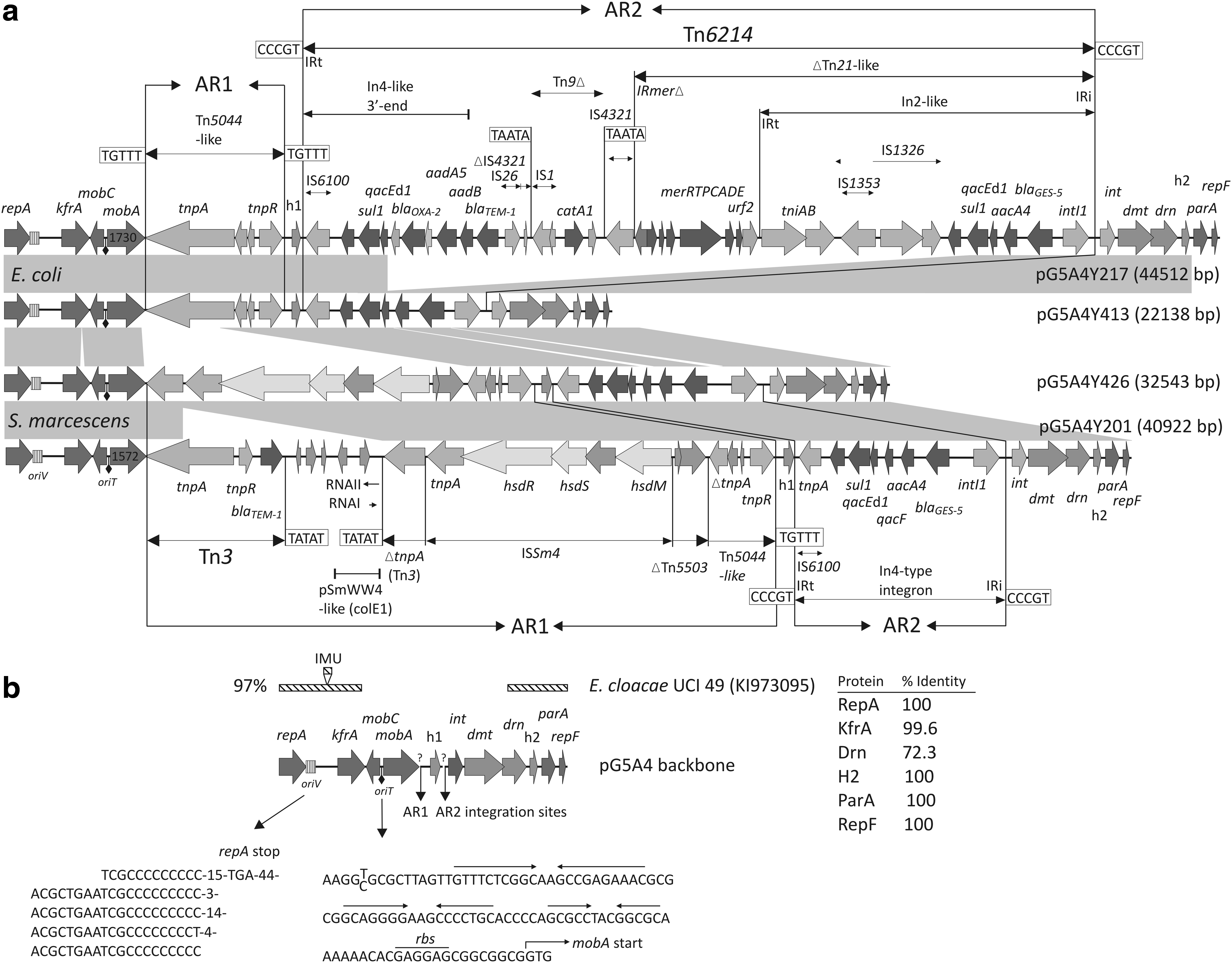
S. marcescens A4Y201 and A4Y426 and E. coli A4Y217 and A4Y413 were collected as part of the CNISP CRGN study after being assessed as having intermediate or full resistance to a carbapenem, as determined by the submitting hospital's standard methodology, and were subsequently sent to the National Microbiology Laboratory for further characterization. 11 Initial PCR analysis showed the two E. coli harbored blaGES, blaTEM, and blaCTX-M group genes and the two S. marcescens harbored blaGES, blaTEM, and blaCMY-2 group genes. Sequence analysis revealed the genes to be blaGES-5, blaTEM-1, blaCTX-M-3, and blaCMY-111, with the latter a new blaCMY-2 variant (Ala238→Val). Whole plasmid analysis revealed that each isolate harbored multiple plasmids (data not shown) and replicon typing showed that all isolates harbored IncA/C and ColE1 replicons. Details of the blaGES-5-containing isolate's case information were collected for each patient (Table 1). Importantly, bacterial isolation dates and length of time in the hospital of the patients before blaGES-5 isolation were as follows: S. marcescens A4Y201 (October 2010, 101 days), E. coli A4Y2017 (April 2011, 76 days), E. coli A4Y413 (June 2012, 25 days), and S. marcescens A4Y426 (January 2013, 39 days). Thus, the two S. marcescens were isolated 795 days apart from the general systems intensive care unit (GSICU) and the two E. coli 454 days apart from the cardiovascular ICU (CVICU). All isolates were nosocomially acquired as based on physician judgment, and there was no intervening identification in any other clinical samples. The two ICUs were found on different floors of the hospital. The blaGES-5 plasmids named pG5A4Y201, pG5A4Y217, pG5A4Y413, and pG5A4Y426 (collectively pG5A4 plasmids) were successfully transformed from each clinical isolate to E. coli DH10B to allow further analysis. PCR analysis showed that the blaCTX-M-3 and blaCMY-111 genes were not cotransformed with blaGES-5, nor was blaTEM-1, except in the case of pG5A4Y217.
CRGN, carbapenem-resistant gram-negative; CVICU, cardiovascular intensive care unit; GSICU, general systems intensive care unit; n.a., not applicable.
Antimicrobial susceptibilities
Antimicrobial susceptibility results are shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr). As is common for organisms harboring ESBLs and/or carbapenemases, the clinical isolates were multidrug resistant (resistant to three or more drug classes) and exhibited resistance to third-generation cephalosporins, β-lactam and inhibitor combinations, ertapenem, and ciprofloxacin, and were intermediate or resistant to gentamicin and tobramycin, but susceptible to amikacin. Three of the four isolates were resistant to meropenem and imipenem with E. coli A4Y413 the exception. However, E. coli AY413 was resistant to ertapenem (minimum inhibitory concentration [MIC]=2 μg/ml), and the MICs for meropenem and imipenem (≥1 μg/ml) also met the CLSI screening criteria for a potential carbapenemase-producing organism. MICs were lower in the transformants harboring the pG5A4 plasmids compared with the clinical isolates, but were still fourfold or greater than in the host E. coli DH10B (Supplementary Table S1). For the clinical isolates, higher MICs could be due to the production of the CTX-M or CMY-111 enzymes with concomitant porin mutations.
Molecular typing
The two S. marcescens and the two E. coli were closely related to each other, respectively, as determined by PFGE analysis (>90% similarity; Supplementary Fig. S1a). MLST analysis showed both E. coli isolates to be the sequence type ST205. Although restriction fragment length polymorphism (RFLP) analysis of the pG5A4 plasmids revealed variant patterns and evident size differences, three bands were shared between all four plasmids (5.5, 0.7, 0.6 kb) and indicated a possible relationship of at least part of each plasmid (Supplementary Fig. S1b).
Plasmid sequences
The molecular epidemiology of the clinical isolates and four plasmids prompted us to determine the complete sequence of the pG5A4 plasmids. Complete listings of the reading frames for the four plasmids are shown in Supplementary Tables S2 and S3. Sequencing revealed that the plasmids shared a nearly identical backbone of ∼10.6 kb with the remaining sequences being partly related accessory regions (AR1 and AR2), which consisted of mobile elements and contained the antimicrobial resistance genes (Fig. 1). The AR1 was integrated in the identical position upstream of a hypothetical protein gene (h1, Fig. 1), but was associated with apparent deletions of different length in the mobA relaxase gene 3′ end as no mobA stop codon was encountered before the beginning of the AR1 sequence. Thus, each mobA gene can be considered a hybrid with the reading frames ending in the AR1. If translated pG5A4Y217 and pG5A4Y413 would produce a 616 amino acid MobA protein, of which only the first 578 residues can be considered native, and pG5A4Y201 and pG5A4Y426 would produce a 526 amino acid MobA protein with the first 524 residues being considered native. In pG5A4Y217 and pG5A4Y413, the AR1 is a transposition module of a Tn5044-like transposon flanked by 5 bp direct repeats (TGTTT), indicating acquisition by transposition. In the AR1 of pG5A4Y201 and pG5A4Y426 much of the Tn5044-like transposon is truncated by a partial Tn5503 end, which itself is truncated by a novel large insertion sequence, ISSm4 that harbors a type 1 restriction–modification system. The ISSm4 tnpA end is linked to the last 1,687 bp of the Tn3 tnpA end, which in pG5A4Y426 interrupts the mobA gene. In the pG5A4Y201 AR1, a ColE1 plasmid with partial homology to the S. marcescens plasmid pSmWW4 (acc. no. CP003960) that had likely acquired a complete Tn3 has probably integrated through recombination between duplicated Tn3 tnpA regions. The AR2 was integrated in the same position in all the plasmids downstream of h1 and upstream of an integrase gene (int, Fig. 1). In the pG5A4Y217 AR2 is a ∼29 kb complex transposon, Tn6214, flanked by 5 bp direct repeats (CCCGT), which is an accretion of parts of other transposons, ISs, and class 1 integrons with the blaGES-5-aacA4 genes as cassettes in an integron at the IRi end. The AR2 in pG5A4Y413 is an In4-like integron with the blaGES-5-aacA4 gene cassettes. Recombination between the duplicated sul1 regions in Tn6214 could lead to a looping out/deletion of the intervening sequence to form the AR2 of pG5A4Y413. The AR2s of pG5A4Y201 and pG5A4Y426 are identical In4-like integrons harboring the blaGES-5-aacA4 gene cassettes, but with some differences from the cassette regions in pG5A4Y217 and pG5A4Y413. The pG5A4Y201 and pG5A4Y426 cassettes also contain a qacF gene downstream of aacA4 and an additional 297 bp of novel DNA between the blaGES-5 and intI1 genes. Outside of the AR1 and AR2, the putative backbone of the pG5A4 plasmids includes genes involved in plasmid replication (repA, repF), segregation/stability (kfrA, parA), transfer (mobA, mobC), two hypothetical protein genes (h1, h2), a DNA invertase/recombinase (int), and a deoxymethyltransferase (dmt) and deoxyribonuclease (drn) of a type II restriction module (Fig. 1). The int-dmt-drn region may be postulated to be an acquired region in the plasmid backbone, although there were no obvious sequence signatures such as repeated DNA associated with this region. In the pG5A4Y201 and pG5A4Y426 kfrA gene, there was an in-frame 63 bp internal tandem duplication (positions 613–675 and 676–738), which resulted in KfrA protein 21 residues larger than that in pG5A4Y217 and pG5A4Y413. The RepA protein shares ≥70% identity with RepA proteins identified in a number of environmental bacteria, including gram-negative nonfermenters, and Enterobacteriaceae, and a partial phylogenetic tree and alignment are shown in Figs. 2 and 3, respectively. In a plasmid classification scheme based on relaxase domains of relaxase proteins, 6 the pG5A4 plasmids belonged to the MOBQ1 group whose prototype is the broad-host-range mobilizable IncQ1 plasmid RFS1010. An alignment of the relaxase domains of five MOBQ1 relaxases with the pG5A4 relaxase domain is shown in Fig. 4. A contig from Enterobacter cloacae UCI 49 (acc. no. KI973095, unpublished), which appears to be a complete plasmid, contains a 5.4 kb region of near identity (∼97%) to the pG5A4 backbone (Fig. 1). A 288 bp integron mobilization unit (IMU) with 90% identity to those found in pCHE-A, an IncQ-type plasmid that harbors blaGES-5, 16 is found between repA and kfrA in E. cloacae UCI 49, but is absent in the pG5A4 backbone. Coincidently, plasmid pCHE-A isolated from a E. cloacae represents the first incidence of blaGES-5 in Canada. The E. cloacae UCI 49 plasmid, which has a large MobA protein belonging to the MOBP14 group, 6 was isolated from a human urine isolate in 2013 in Irvine, California (BioSample no. SAMN02356629), and the patient has no known link to the ones in this study.
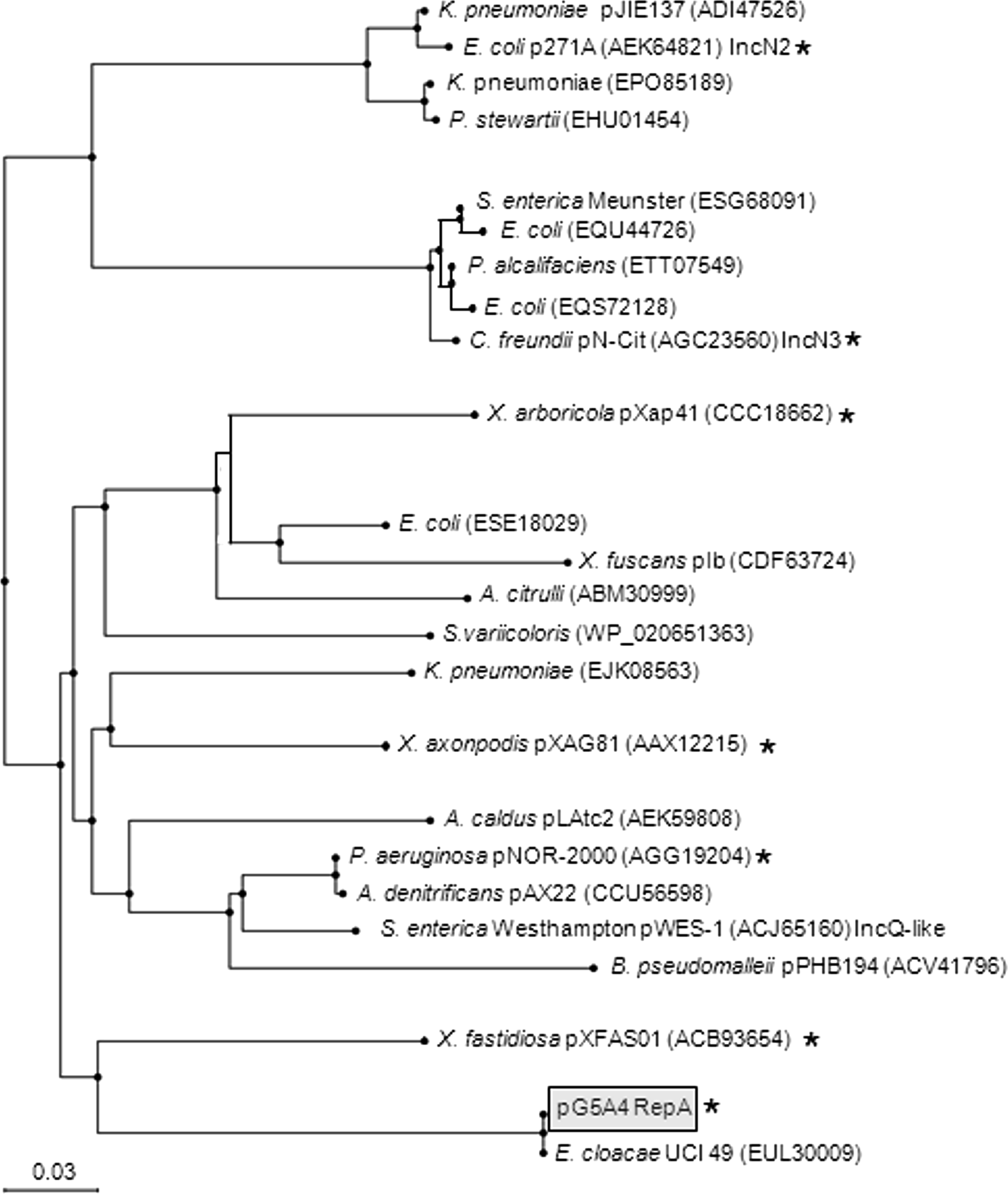
Phylogenetic tree showing the pG5A4 RepA and closely related proteins. The tree was produced on the BLAST Web site using the Blast Tree View Widget after using the pG5A4 RepA protein as the query in a DELTA-BLAST search, followed by a second PSI-BLAST iteration. The parameters for tree construction were Fast Minimum Evolution (Tree method), 0.85 sequence distance (Max Seq Difference), and Grishin-protein (Distance). Accession nos. of the proteins are shown in brackets. When the RepA protein is from a known plasmid, the plasmid name is shown as is the Inc-type if known. Asterisks indicate proteins shown aligned in Fig. 3.

Alignment of the pG5A4 RepA protein with six other related RepA proteins as indicated by asterisks in Fig. 2. Positions where at least three of the proteins share an identical amino acid are boxed.

Alignment of the relaxase domains of MOBQ1 relaxases related to the pG5A4 relaxase. The three motifs that define this group are indicated. Black triangles indicate residues involved in the catalytic center. Positions where at least three of the proteins share an identical amino acid are boxed. Proteins (accession nos.) are from Nitrosospira multiformis (ABB76106), Sphingomonas sp. PAMC26617 (WP_010165461), Acinetobacter sp. NIPH713 (ENW89148), Thiothrix flexilis (WP_020560601), and plasmid RSF1010 (AAA26445).
Plasmid transfer experiments
An oriV and a putative oriT have been identified in the pG5A4 plasmids (Fig. 1). Given the size and gene content of the pG5A4 backbone region, they might be expected to be mobilizable, but not necessarily self-transmissible despite containing truncated mobA genes (see Plasmid sequences section). Using E. coli A4Y217 (clinical isolate, 616 amino acid MobA), S. marcescens A4Y201 (clinical isolate, 526 amino acid MobA), and E. coli A4Y217T (transformant, no other coresident plasmids) as donors, transconjugants harboring blaGES-5 and the pG5A4 repA gene were only obtained with the clinical isolate E. coli A4Y217. We lacked a selectable S. marcescens strain to act as a recipient in mating experiments. We note that the transconjugants, which did not harbor an IncA/C replicon, appeared to contain at least two plasmids of sizes differing from pG5A4Y217 (data not shown). Thus, blaGES-5 transfer may have been achieved by mobilization with accompanying plasmid recombination or, alternatively, a self-transmissible cointegrate harboring blaGES-5 and the pG5A4 repA gene may have been transferred.
Discussion
Remarkably, two closely related E. coli isolated >1 year apart and two closely related S. marcescens isolated >2 years apart, all harboring a novel transferable MOBQ1 multidrug-resistant plasmid containing the blaGES-5 carbapenemase were isolated from four different patients. No other clinical isolate containing the plasmid was identified during this time period in patients from this hospital. The four isolates were identified from patients in two physically separated ICUs, although patients are often moved between the units. It would be extremely unlikely that undetected patient-to-patient transmission occurred during this time period; therefore, a persistent environmental source is postulated. Although it is credible that S. marcescens may exist in the hospital inanimate environment for extended periods, this seems less likely in the case of E. coli. It seems most plausible that the pG5A4-type plasmids are harbored by environmental bacteria and occasionally are transferred to the clinically relevant gram-negative organisms. Indeed, blaGES-5 has been found on untyped plasmids harbored by Kluyvera intermedia that were recovered from sinks and taps in a Brazilian hospital. 18 Small pG5A4 parental plasmids could have acquired the accessory regions or parts of, after transfer into gram-negative organisms harboring drug resistance genes on other plasmids. It has been previously found that MOBP-type plasmids harboring blaGES-5 have been isolated from unknown bacteria from a wastewater treatment plant in Germany. 7 In addition, Tn5044 associated with mercury resistance operons have commonly been found in gram-negative environmental bacteria, 12 and Tn5503 has been found in the IncP-6 plasmid Rms149 found in P. aeruginosa. 9 Indeed, Rms149 (57 kb) is similar to the pG5A4 plasmids, in that it consists of a small mobilizable backbone comprising only 21% of its sequence, with the rest consisting of multiple mobile elements. 9 Like Rms149, the pG5A4 plasmid backbone may consist of a combination of gene modules from other plasmid systems, hence their association with a MOBQ1 relaxase, whereas the near identical E. cloacae UCI 49 plasmid backbone contains a MOBP14 relaxase. Whether the hybrid MobA proteins are produced and contribute to the plasmids transferability is unknown. Relaxase proteins are generally large proteins with multiple domains, including the N-terminal relaxase (nickase) domain, and others with helicase, primase, or unknown functions.1,8 Thus, the pG5A4 plasmids MobA do contain the relaxase domain (∼225 amino acids) and may also harbor additional functionality if in fact active. The majority of IncA/C plasmids are self-transmissible17,20 and many have been shown to mobilize the Salmonella genomic island 1. 5 Thus, it was still possible that the IncA/C plasmid in the donor may have contributed transfer functions in trans to allow the horizontal transfer of blaGES-5 and the pG5A4 replicon.
The long-term persistence of a transferable plasmid harboring the blaGES-5 carbapenemase, potentially arising from an environmental source, has implications for infection control and clinical care when it is transferred to clinically relevant gram-negative organisms.
While this manuscript was in proof a carbapenem resistant S. marcescens was isolated from a patient in the hospital where the four isolates in this study were obtained. PCR, sequence analysis, and transfer experiments revealed that this isolate harbored blaGES-5 on a pG5A4-type plasmid. Subsequent environmental screening lead to the isolation a Citrobacter freundii that harbored blaGES-5 on a pG5A4-type plasmid. This strain was isolated from a sink located on the same ward as the above patient. Thus, the pG5A4-type plasmid has continued to persist for the 23 months after its last isolation and an environmental reservoir has been identified.
Footnotes
Acknowledgments
The authors gratefully acknowledge the Infection Control Practitioners who collected epidemiological data for the CNISP, and the expert technical assistance of Romeo Hizon and Ken Fakharuddin.
The authors would like to thank the following members of the Canadian Nosocomial Infection Surveillance Program (CNISP): David Boyd, National Microbiology Laboratory, Public Health Agency of Canada; Elizabeth Bryce, Vancouver General Hospital, Vancouver, BC; John Conly, Foothills Medical Centre, Calgary, AB; Janice Deheer, Kelowna General Hospital, Kelowna, BC; John Embil, Health Sciences Centre, Winnipeg, MB; Joanne Embree, Health Sciences Centre, Winnipeg, MB; Gerard Evans, Kingston General Hospital, Kingston, ON; Sarah Forgie, Stollery Children's Hospital, Edmonton, AB; Charles Frenette, McGill University Health Centre, Montreal, QC; Camille Lemieux, University Health Network, Toronto, ON; George Golding, National Microbiology Laboratory, Public Health Agency of Canada; Denise Gravel, Centre for Communicable Diseases and Infection Control, Public Health Agency of Canada; Elizabeth Henderson, Peter Lougheed Centre, Calgary, AB; James Hutchinson, Health Sciences Centre, St. John's, NL; Michael John, London Health Sciences Centre, London, ON; Lynn Johnston, Queen Elizabeth II Health Sciences Centre, Halifax, NS; Kevin Katz, North York General Hospital, Toronto, ON; Pamela Kibsey, Victoria General Hospital, Victoria, BC; Magdalena Kuhn, South East Regional Health Authority, Moncton, NB; Joanne Langley, IWK Health Centre, Halifax, NS; Nicole Lesaux, Children's Hospital of Eastern Ontario, Ottawa, ON; Mark Loeb, Hamilton Health Sciences Corporation, Hamilton, ON; Anne Matlow, Hospital for Sick Children, Toronto, ON; Allison McGeer, Mount Sinai Hospital, Toronto, ON; Robyn Mitchell, Centre for Communicable Diseases and Infection Control, Public Health Agency of Canada; Dorothy Moore, Montreal Children's Hospital, McGill University Health Centre, Montreal, QC; Michael Mulvey, National Microbiology Laboratory, Public Health Agency of Canada; Linda Pelude, Centre for Communicable Diseases and Infection Control, Public Health Agency of Canada; Virginia Roth, The Ottawa Hospital, Ottawa, ON; Andrew Simor, Sunnybrook Health Sciences Centre, Toronto, ON; Kathryn Suh, The Ottawa Hospital, Ottawa, ON; Geoffrey Taylor, University of Alberta Hospital, Edmonton, AB; Eva Thomas, Children's and Women's Health Center, Vancouver, BC; Nathalie Turgeon, Hôtel-Dieu de Québec du CHUQ, QC; Mary Vearncombe, Sunnybrook Health Sciences Centre, Toronto, ON; Joseph Vayalumkal, Alberta Children's Hospital, Calgary, AB; Karl Weiss, Maisonneuve-Rosemont Hospital, Montreal, QC; and Alice Wong, Royal University Hospital, Saskatoon, SK.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.