
Abstract
Introduction
B
During the last decade, livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) has emerged throughout Europe and North America, and food production animals are increasingly recognized as a potential reservoir of MRSA. 21 The predominant clonal lineage of LA-MRSA belongs to clonal complex (CC) 398 and pigs are particularly regarded as a reservoir of this clone.21,20 Even though MRSA CC398 rarely causes infections in animals, this clonal lineage has attracted much attention due to its potential to colonize and cause infections in humans. 21 Human acquisition of MRSA CC398 is regarded as an occupational health risk; people in close contact with colonized animals, such as farmers and veterinarians, are at greater risk of getting infected/colonized. 21
In the period 2012 until May 2014, four people with no contact to pig farming have died due to bacteremia caused by MRSA CC398 in Denmark.56,57 Denmark has an annual pig production of ∼30 million pigs and is one of the world's largest exporters of pork.16,17 The most prevalent methicillin-sensitive S. aureus (MSSA) among Danish pigs belongs to CC398 (primarily spa type t034). 25 Until recently, pig-associated MRSA had only been identified in CC398 in Denmark. However, a study by Agersø et al. detected the presence of MRSA sequence type (ST)433 (spa type t1333) belonging to CC30 in pigs at slaughter, and it was suggested that MRSA CC30 may have the potential to spread among Danish pigs as MSSA CC30 (primarily ST433) is the second most common S. aureus genotype in Danish pigs. 6
In a recent study, one human case of bacteremia caused by ST433 MSSA was detected. The isolates of ST433 from pigs and humans were clonally related, indicating the potential of pig to human transfer. 5 MRSA CC30, causing infections in humans globally, belongs to four clonal lineages. Although ST433 belongs to the same CC (CC30), it groups distinctly from the four clonal lineages based on pulsed-field gel electrophoresis and it is unknown whether ST433 can adapt to humans and spread clonally. 37
The emergence of LA-MRSA and its potential to colonize and infect humans poses an important and increasing risk for public health. The discovery of yet another clonal lineage of LA-MRSA (CC30) in Danish pigs, which is also carried by farmers, is worrying. Effective use of biocides may be important to reduce/eradicate MRSA in pig herds. Knowledge on the emergence of bacterial isolates from livestock animals with reduced susceptibility to biocides used in their environment is limited. Therefore, we investigated porcine MRSA and MSSA isolates belonging to either CC398 or CC30 for their susceptibility to active compounds of commonly used disinfectants in pig production. These disinfectants comprise benzalkonium chloride (QAC), hydrogen peroxide and sodium hypochlorite (highly oxidizing compounds), formaldehyde (aldehyde compound), and caustic soda (strong alkaline compound).55,62 Furthermore, the antimicrobial resistance profile, hemolysis properties, and the presence of toxic shock syndrome toxin-1 (TSST-1) and Panton-Valentine Leukocidin (PVL)-encoding virulence genes were investigated.
Materials and Methods
Chemicals and reagents
Active disinfectant compounds included in this study were benzalkonium chloride, 50% (w/w) aqueous solution (VWR); sodium hypochlorite, available chlorine 10–15% (Sigma-Aldrich); formaldehyde solution, ≥37% (Merck); hydrogen peroxide, ≥30% (Fluka), and sodium hydroxide, ≥98% (Sigma-Aldrich). Premade tryptone soya agar plates with 5% sheep blood (blood agar plates) were purchased from Oxoid. BBL™ Mueller–Hinton II broth (MHB) and agar (MHA) were from Becton, Dickinson and Company. Tryptic soy broth (TSB) and agar (TSA) were from Oxoid. Sheep blood was from Statens Serum Institute.
Strain collection
A total of 79 porcine S. aureus isolates of CC398 (61) and CC30 (18) were included in this study (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr) and comprise 21 MSSA and 40 MRSA isolates belonging to CC398 and 13 MSSA and 5 MRSA isolates belonging to CC30. The isolates were chosen based on spa type, sampling date, and geographic origin to represent different types of CC398 and CC30 isolated from pigs in Denmark from 2000 to 2011.
Twenty-five of the isolates were from diagnostic submissions to The National Veterinary Institute or The National Food Institute, Technical University of Denmark.6,18 Fifty-four colonizing S. aureus isolates were sampled from healthy pigs (nasal swabs) at 12 different farms or from healthy pigs at seven different slaughter plants (nasal swabs before scalding) as part of the DANMAP surveillance program in 2009.6,25,37 The origin of two of the colonizing pig isolates was unknown. For isolates collected at slaughter plants, only one sample per month was collected per Central Husbandry Register (CHR) number. 6 Isolates originating from the same farm were collected on the same day, but from different animals. In one case, two isolates were collected from the same pig, however, they belonged to CC30 and CC398, respectively, and were therefore both included in the study.
Species isolation and identification of S. aureus of MRSA were performed as previously described.1,6,25,35 The presence of the mecA gene encoding methicillin resistance was confirmed by multiplex polymerase chain reaction (PCR) as previously described. 35 The spa type of isolates was determined following the recommendations at SeqNet 50 using recommended primers25,18 or primer pair 1794 (5′-AGACGATCCWTCAGTGAGC-3′) and 1827 (5′-TAATCCACCAAATACAGTTGTACC-3′). 6 Based on the amplified sequence, spa types were assigned to each isolate using the spa plugin in the BioNumerics v4.6 software (Applied Maths).6,25,18
Determination of the minimal inhibitory concentration and minimal bactericidal concentration to biocides
Determination of minimal inhibitory concentrations (MICs) to benzalkonium chloride (0.250–128 μg/ml), hydrogen peroxide (0.00025–0.128%), sodium hypochlorite (0.003125–1.6%), formaldehyde (0.000781–0.4%), and caustic soda (0.003125–1.6 M) were determined according to the CLSI document M31-A7 for susceptibility testing of bacteria isolated from animals. 14 Selected isolates were also tested for their susceptibility to chlorhexidine (0.0625–32 μg/ml). Briefly, bacteria from overnight (ON) blood agar plates were resuspended in 0.9% NaCl to a McFarland standard of 0.5 (∼108 CFU per ml) and further diluted 100-fold in MHB. The size of the inoculums was determined by plating MHB cell suspensions on MHA plates and incubated ON at 37°C.
Working solutions of biocides were prepared in MHB, and twofold dilution series were prepared in 96-well round-bottom microtiter plates (50 μl), where after 50 μl of MHB, cell suspensions were transferred to the microtiter plates (Nunc) (∼5×105 CFU per ml in wells). Wells without biocides were included as positive growth controls and wells without bacteria, but containing a biocide concentration twofold lower than the lowest concentration of the test interval, served as sterility controls. Plates were incubated at 37°C for 16–20 hr in an aerobic atmosphere, after which cell growth was inspected by the eye.
The MIC value was defined as the lowest concentration of the compounds giving rise to no visible growth. The MBC was determined according to the CLSI (formerly NCCLS) guideline M26-A 41 for determining the bactericidal activity of antimicrobial agents, with minor adjustments. After the MIC values were read, taking ∼15 min, microtiter plates were further incubated until a total incubation time of 24 hr was reached. Then, pellets were resuspended and 10 μl was spotted on MHA plates from wells with no visible growth, wells containing the highest concentration of biocides and where cell growth was visible, and from wells, including the sterility control. Plates were incubated at 37°C and colonies were counted after 48 hr of incubation. Biocide concentrations leading to a 99.9% reduction of viable cells were regarded as the MBC values.
The MIC and MBC determinations were performed in duplicates and only results giving rise to similar values or that deviated by one dilution step were included. In the case of variation within the duplicate determination, the highest value was selected. S. aureus ATCC 29213 was included as the control strain for reproducibility of the susceptibility testing procedure.
Ethidium bromide accumulation assay
Isolates were screened for the presence of active QAC efflux pumps using an ethidium bromide accumulation assay as previously described. 59 Isolates were grown for 24 hr at 37°C on MHA containing ethidium bromide (0.5 μg/ml), followed by inspection for fluorescence under UV light. Cells accumulating ethidium bromide had red fluorescence and cells that did not accumulate fluorescence were white and defined to include active efflux pumps. The strain, JCM 16556 (qacB), that overexpresses efflux pumps, was included as a positive control strain.
Detection of qac genes
The four porcine S. aureus isolates (75, 76, 78, and 79) found not to accumulate ethidium bromide had previously been whole-genome sequenced. 48 Identification of QAC resistance genes in these isolates was performed by BLASTing nucleotide sequences of all qac genes described to date (qacA/B, smr, qacE, qacEΔ1, qacF, qacG, qacH, qacI, qacJ, qacZ) from the NCBI homepage (www.ncbi.nlm.nih.gov/) with sequences of the sequenced isolates.
Determination of the MIC to antimicrobial agents
Susceptibility to ciprofloxacin (0.12–8 μg/ml), erythromycin (0.12–16 μg/ml), florfenicol (1–64 μg/ml), chloramphenicol (2–64 μg/ml), penicillin (0.06–16 μg/ml), spectinomycin (8–256 μg/ml), streptomycin (2–128 μg/ml), sulfamethoxazole (8–512 μg/ml), tetracycline (0.5–32 μg/ml), tiamulin (0.25–32 μg/ml), and trimethoprim (1–32 μg/ml) had earlier been or were determined using the semiautomated Sensititre broth microdilution system (Trek Diagnostic Systems) as previously described. 2 Selected isolates were also tested for their susceptibility to gentamicin (0.25–16 μg/ml). Plates were inoculated and incubated according to the CLSI document M07-A8. 14 S. aureus ATCC 29213 was used as the quality control strain.
The European Committee on Antimicrobial Susceptibility Testing (EUCAST) epidemiological cutoff values (ECOFF) 22 were used to interpret MIC results as recommended by the EU Reference Laboratory for Antimicrobial Resistance (EURL-AR). 23
Virulence of isolates
lukPV and tst detection
All isolates were screened for the virulence genes, lukPV and tst, using primers previously described (see list below). DNA extraction and the PCR program were the same as described above in the Detection of qac genes section, except for the annealing temperature for tst that was set to 52°C. The lukPV-positive S. aureus, USA300 LAC, 19 and tst-positive S. aureus, Sa564, 24 were used as positive controls.
lukPV: Luk-PV-1 ATCATTAGGTAAAATGTCTGGACATGATCCA and
Luk-PV-2 GCATCAAGTGTATTGGATAGCAAAAGC. 32
tst: TST-1: TST-1: AGCATCTACAAACGATAATATAAAGG and
TST-2: CATTGTTATTTTCCAATAACCACCCG. 9
glp dn TGG TAA AAT CGC ATG TCC AAT TC and
glp up CTA GGA ACT GCA ATC TTA ATC C. 39
Hemolysis assay
Delta-hemolytic activities of individual bacterial isolates were assessed by cross-streaking test isolates perpendicular to the beta-hemolysin hyperproducing S. aureus RN4220, a strain derived by multiple mutagenesis procedures from the parent strain NCTC8325, 61 on sheep blood agar plates. Hemolysis was assayed after 24 hr at 37°C and again after additional 24 hr at 4°C. The latter step was performed to evoke the hot–cold lysis phenomenon observed with beta-hemolysis. 4 The delta-hemolytic activity was denoted by an enhanced area of hemolysis in the intersection of RN4220 and test isolate streaks.
Results
Susceptibility to biocidal compounds
The MIC and MBC distributions of the 21 MSSA and 40 MRSA isolates belonging to CC398 and the 13 MSSA and 5 MRSA isolates belonging to CC30 are shown in Fig. 1. The MIC and MBC values of the individual isolates are accessible in the supplementary material (Supplementary Table S2). The MIC and MBC distributions to each of the biocidal compounds formed one large population of isolates, respectively. All tested isolates had MICs and MBCs within a range of 1–8 and 2–16 μg/ml to benzalkonium chloride, 0.001–0.002% and 0.002–0.016% to hydrogen peroxide, 0.025–0.1% and 0.05–0.4% to sodium hypochlorite, 0.00313–0.0125% and 0.00625–0.025% to formaldehyde, and 0.025–0.2 and 0.1–0.2 M to caustic soda, respectively.
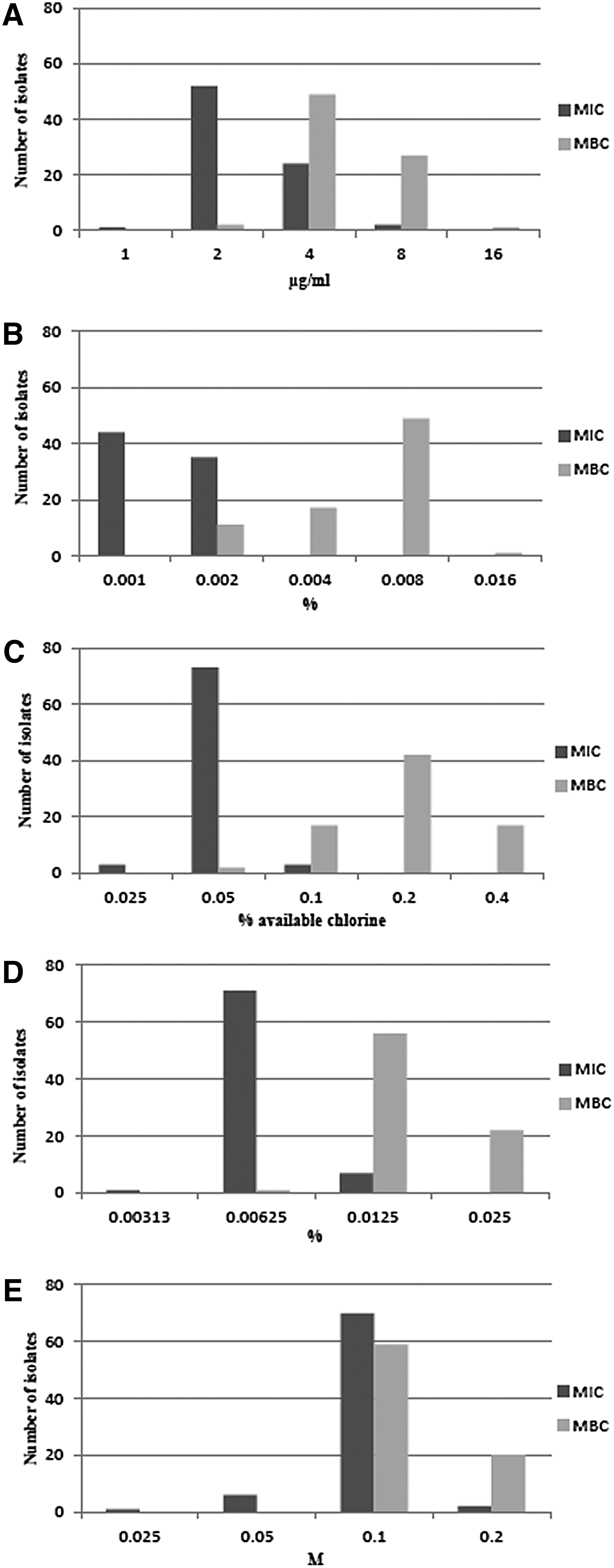
Distributions of the minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) of the 79 Staphylococcus aureus pig isolates to
The paradoxical effect, also known as the eagle effect, was in some cases observed in the MBC determination of hydrogen peroxide (Supplementary Table S2), namely that the number of surviving cells increased with increasing hydrogen peroxide concentration beyond the lowest concentration giving rise to≥99.9% kills. 41
Susceptibility testing of the control strain was reproducible within a twofold range and the MIC and MBC values were within the range of the tested isolates (Table 1). The method used for MIC determination showed good agreement within duplicate determinations; duplicates were shown to have the same values in 87–99% of the cases, depending on the biocide tested. A greater variance was observed for determination of MBCs in which duplicates varied between 66% and 81%. However, in all cases, duplicate determinations did not vary more than one dilution step.
MBC, minimal bactericidal concentration; MIC, minimal inhibitory concentration; n, number of runs.
Detection of qacG
All isolates were screened for the presence of an active QAC resistance efflux pump using an ethidium bromide accumulation assay. Four MRSA CC30 isolates (75, 76, 78, and 79) did not accumulate ethidium bromide, indicating the presence of an active efflux mechanism (Fig. 2). These isolates had previously been whole-genome sequenced and importantly the QAC resistance gene, qacG, was identified in three of these isolates, and none of the isolates contained other QAC genes (76, 78, and 79). Sequence contigs, including the qacG gene from the whole-genome sequenced isolates (isolate no. 76, 78, and 79), were analyzed by BLAST against the nucleotide database at the NCBI homepage (www.ncbi.nlm.nih.gov/) to identify the possible location of qacG in these isolates.

Ethidium bromide accumulation assay. Isolates were grown for 24 hr at 37°C on TSA containing ethidium bromide (0.5 μg/ml), followed by inspection for fluorescence under ultraviolet (UV) light. S. aureus strain, JCM 16556 (qacB), that overexpresses efflux pumps was used as a Qac efflux pump positive control strain and S. aureus strain, 8325-4, as a Qac negative control strain. Figure illustrates results for the methicillin-resistant Staphylococcus aureus (MRSA) CC30 isolates.
Two isolates (no. 76 and 78) had 99% identity to the full qacG corresponding to position 1532–1855 (GenBank Accession No. Y16944), and one isolate (no. 79) had a partial sequence on the contig corresponding to position 1560–1855, also with more than 99% homology. The size of the contigs varied from 2,093 to 2,222 bp for three of the isolates, and the average number of reads was ∼75–130 times higher than the average number of reads from the whole-genome sequence runs of the same isolates, indicating qacG being present in multiple copies. The size of the contigs is very similar to the size of pST94 and the higher number of reads indicates that qacG is likely present on a plasmid rather than in the chromosome. Moreover, on the same contigs, a putative replicon was identified with 92–93% homology to rep94 of pST94. This indicates that qacG in isolates 76, 78, and 79 resides on pST94 or a pST94-like plasmid. The qacC gene was identified in the fourth isolate (75) and was present on a 3,210 bp contig, which showed 100% identity to the plasmids, SAP82A (GQ900434 position 40798–44007), pG01 (FM207042 position 41312–38103), and pLW043 (AE017171 position 222245–19036), originating from S. aureus. These plasmids vary in size from 44 to 58 kb, and the region corresponding to the contig contained, in addition to the qacC, a putative repA replicon. The average number of reads was ∼3.5 times higher than the average number of reads from the whole-genome sequence runs of the same isolate, indicating that qacC is present in multiple copies, probably on a plasmid.
Susceptibility to antimicrobial agents
The phenotypic resistance profiles and distribution of resistance patterns of the 79 S. aureus isolates are shown in Tables 2 and 3. S. aureus belonging to CC398 represented 23 different phenotypic resistance patterns. All of the CC398 isolates were tetracycline resistant and 84% and 56% of the isolates showed resistance to trimethoprim and streptomycin, respectively. Antimicrobial resistance to erythromycin (46%), spectinomycin (38%), and tiamulin (33%) was also frequently observed, whereas resistance to ciprofloxacin (20%) was found to a lesser extent. Only one isolate showed resistance to chloramphenicol–florfenicol (2%) and sulfamethoxazole (2%), respectively. Thirty-one (78%) and 16 (76%) of the MRSA and MSSA CC398 isolates, respectively, were multiresistant (defined as resistance to three or more classes of antimicrobial agents other than the group of β-lactams). The S. aureus CC30 isolates were arranged in nine different phenotypic resistance patterns. None of the MSSA CC30, but one of the MRSA CC30 isolates, was multiresistant. The group of MRSA CC30 isolates showed resistance to penicillin (100%), tiamulin (100%), erythromycin (20%), trimethoprim (20%), and streptomycin (100%), whereas resistance to penicillin (31%), tiamulin (46%), erythromycin (8%), trimethoprim (8%), tetracycline (15%), spectinomycin (8%), and streptomycin (8%) was present in MSSA CC30 isolates. Of the total number of S. aureus CC30 isolates, 11% showed phenotypic resistance to tetracycline.
CHL, chloramphenicol; CIP, ciprofloxacin; ERY, erythromycin; FFN, florfenicol; PEN, penicillin; SPE, spectinomycin; STR, streptomycin; SMX, sulfamethoxazole; TET, tetracycline; TIA, tiamulin; TMP, trimethoprim; D, diagnostic; F, farm; S, slaughter plant; N, unknown; MRSA, methicillin-resistant S. aureus; MSSA, methicillin-sensitive S. aureus.
Toxin production of S. aureus isolates
Three of the tested isolates (38, 39, and 67) were delta-toxin producing (Fig. 3), three isolates were not producing any toxins, and the rest either produced alpha- and/or beta-hemolysins.
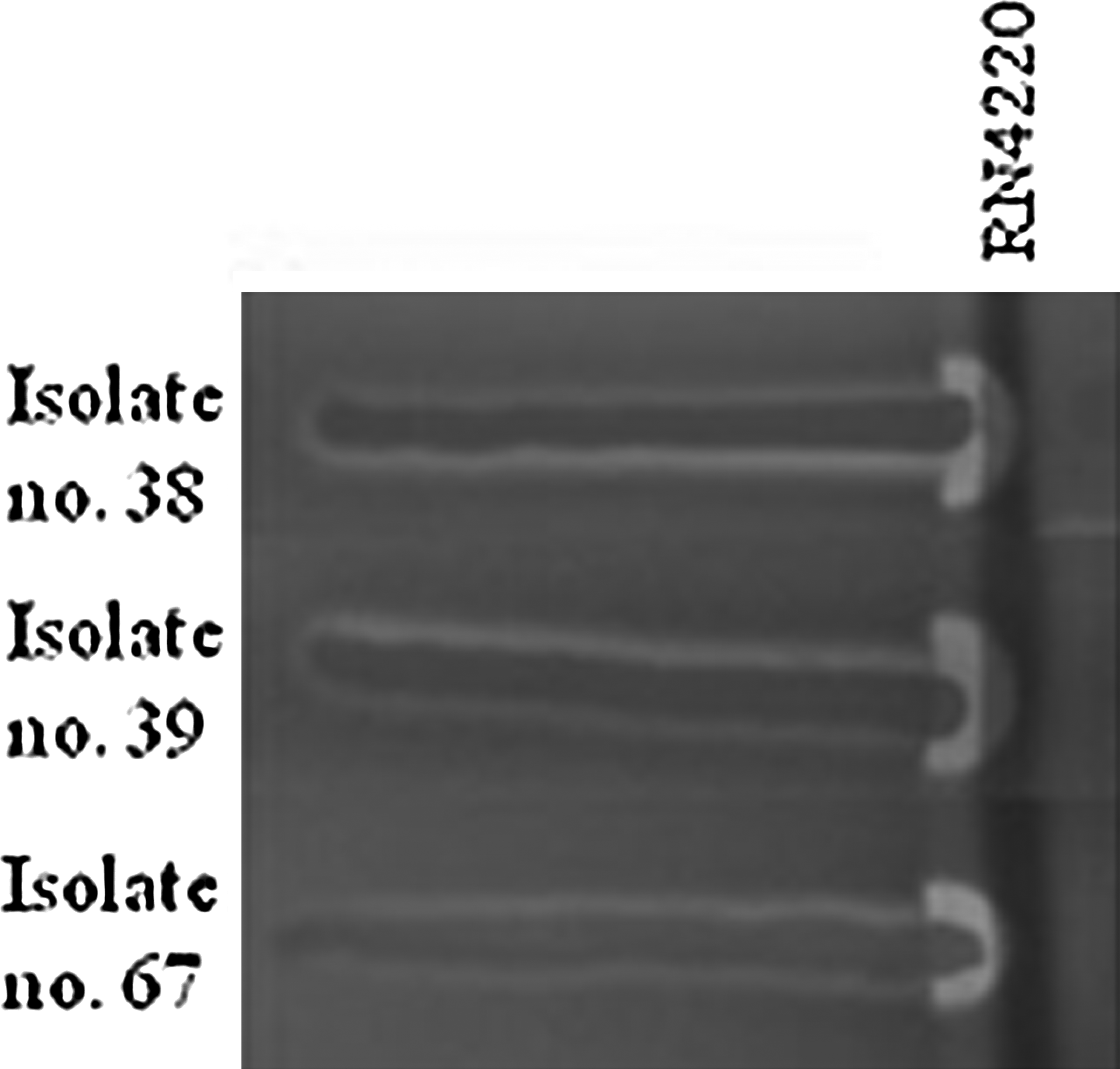
Hemolytic activity of S. aureus on sheep blood. RN4220, a strong beta-hemolysin producer, is streaked vertically and isolates horizontally. Beta-hemolysin forms a turbid zone of hemolysis surrounding the vertical streak of RN4220. Delta-hemolysis and beta-hemolysis are synergistic, producing a clear hemolysis where they intersect.
Discussion
In this study, we tested the susceptibility of porcine S. aureus CC398 and CC30 isolates to active biocidal compounds commonly used for disinfection in the pig production. In addition, the susceptibility to antimicrobial agents, the presence of QAC resistance efflux pumps, and some important virulence factors were investigated.
Susceptibility of porcine S. aureus isolates to biocidal compounds
Results for the susceptibility testing to biocides in this study indicate that there has been no development of resistance to benzalkonium chloride, hydrogen peroxide, sodium hypochlorite, formaldehyde, or caustic soda since each of the MIC and MBC values of all the isolates formed one large population in their susceptibilities to these compounds. These results are similar to the susceptibility testing of other bacterial species from pigs to benzalkonium chloride, hydrogen peroxide, and formaldehyde, where the susceptibilities to these compounds also were found to form one large population. 3 Furthermore, the MIC values of S. aureus isolates to benzalkonium chloride, hydrogen peroxide, sodium hypochlorite, and formaldehyde and MBC values to benzalkonium chloride, hydrogen peroxide, and sodium hypochlorite were very similar to previous findings.3,30,43,46,58,64
No noteworthy difference in the susceptibilities was observed between MSSA and MRSA isolates or between CC398 and CC30 isolates. The results, however, do not give information about a time-dependent change in the susceptibility to the compounds tested. In a study by Skovgaard et al., they investigated the triclosan susceptibility of Danish human clinical Staphylococcus epidermidis blood isolates from before (old) and after the introduction of (current) triclosan in Denmark. 53 They found that the collection of current isolates included significantly more triclosan-tolerant isolates (12.5%) compared with the collection of old isolates (0%). The authors suggested that the wild-type S. epidermidis population structure had changed due to adaptation to the widespread use of triclosan, resulting in a triclosan-tolerant subpopulation present in Denmark. 53 However, it was not possible to include the old isolate to examine a possible change in the population structure to biocides included in this study.
Detection of the QAC resistance genes, qacG and qacC
Disinfectants based on QACs have various applications in veterinary medicine and play an important role in the control of animal disease. Efflux-mediated resistance to QACs has been described in S. aureus and several QAC resistance genes have been identified.11,12 Staphylococci carrying qac genes do not always give rise to an increase in the MIC value toward benzalkonium chloride.29,64 The porcine S. aureus isolates were therefore screened in an ethidium bromide accumulation assay to estimate efflux pump activity. Four MRSA CC30 isolates were found not to accumulate ethidium bromide, and the presence of the QAC resistance gene, qacG, was identified in three isolates and qacC in one. This study is the first to describe the presence of qac genes in S. aureus of porcine origin in Europe and in LA-MRSA belonging to CC30. The qacG and qacC genes have previously been identified in food-related staphylococci.10,28,64 QacG and QacC have a more narrow spectrum of substrates compared with the QacA efflux pump, encoded by qacA, which is generally found to be associated with clinical Staphylococcus spp. 29 The presence of a qac gene (also qacG and qacC) has once before been reported in MRSA isolates from pigs in Hong Kong. However, these pig-associated MRSA isolates belonged to CC9 (t899), a S. aureus clone associated with colonization of pigs in Asia. 64
Two of the strains carrying the qacG gene from our study were found to have higher MIC values to benzalkonium chloride (8 μg/ml) compared with the isolates (which accumulated EtBr), but the other two (positive for qacG and qacC, respectively) isolates had similar MIC values (4 μg/ml) as some of the EtBr-accumulating isolates. The MBC values for qacG positive and some of the EtBr-accumulating isolates were the same (8 mg/ml), as found also in accordance with the study by Wong et al. 64 The susceptibility to benzalkonium chloride still remains lower than the recommended working concentration of benzalkonium chloride and may therefore not have relevant practical implications. However, a study by Otter et al. showed that the use of chlorhexidine for decolonization of MRSA gave rise to an increase in MRSA isolates carrying the qacA gene (chlorhexidine is a substrate for QacA) depending on the clonal type. This was even though the level of resistance to chlorhexidine in qacA-positive isolates was much lower than the applied concentration. 44 Furthermore, Heir et al. demonstrated that qacG-positive Staphylococcus spp. isolates could become adapted to grow at higher benzalkonium chloride concentrations after exposure to sublethal levels of this compound. 28 Incorrect use of disinfectants might give rise to bacterial exposure to lower level QAC concentrations, give strains harboring QAC resistance genes an advantage, and select for subpopulations carrying these genes. 26
qac genes have been found to coexist on genetic elements, such as plasmids and integrons, with genes encoding resistance to important antimicrobial compounds, such as penicillin, trimethoprim, and gentamicin, isolated from clinical, animal, or food-related staphylococci.10,13,34,40,45,47,51,52,63 Use of QACs has therefore been suggested to possibly select for antimicrobial resistance genes located on the same genetic element as the qac genes. 26 The qacG gene was first described by Heir et al. and found to reside on a small plasmid (pST94), containing two open reading frames encoding the QacG and a putative replication protein. 28 Sequence analysis of whole-genome sequenced isolates with qacG suggested that the gene is located on individual plasmids constituting pST94 or a pST94-like plasmid.
The qacG gene has previously been found to reside on a closely related pST94 plasmid isolated from bovine staphylococci 10 and has so far not been associated with plasmids carrying additional resistance genes.10,28 The qacC gene has previously been detected on large plasmids (more than 40 kb) also found to carry resistance genes, and the isolate (no. 75) had qacC on a contig identical to a region on a large plasmid carrying vancomycin resistance genes (plw043), 63 but although the isolate was resistant to several antimicrobials, the resistance genes were not found on the same contig as qacC.
CC30 isolates were in general more susceptible to non-beta-lactam antibiotics than the tested CC398 isolates (Tables 2 and 3). Even though the CC30 isolates, including qacG, probably carry this gene on a plasmid not associated with resistance genes, all of the isolates with qacG were MRSA isolates. One isolate was additionally resistant to gentamicin, another drug of critical importance for the treatment of human infections. Therefore, the use of QACs may contribute to spread of antimicrobial resistance by favoring these MRSA CC30 strains.
Isolates showed a general low hemolytic activity and were all found to be negative for the virulence genes, pvl and tsst, that are frequently observed in HA- and CA-MRSA isolates. However, virulence genes are commonly found on mobile genetic elements and since S. aureus is excellent at acquiring foreign DNA material, development of more virulent strains might just be a matter of time. Surveillance of the development of reduced susceptibility to biocides and antimicrobial resistance and development of more virulent strains among porcine S. aureus isolates is important to determine possible risks for the public health.
Conclusion
Detection of qac genes, qacG and qacC, in European porcine MRSA isolates was for the first time described in this study. These findings are worrying since incorrect use of or the presence of residual concentration of QACs may contribute to the spread and persistence of MRSA CC30 isolates among Danish pigs and complicate the task of MRSA eradication. Virulence factors important for human disease seem uncommon in LA isolates, but surveillance is important to detect changes in the pathogenesis or host specificity of such isolates.
Footnotes
Acknowledgments
This work was funded by a grant, 2101-08-0030, from the Program Committee for Food, Health and Welfare under the Danish Council for Strategic Research. The strains, JCM 16555 (qacA) and JCM 16556 (qacB), were kindly provided by the Japan Collection of Microorganisms, RIKEN BRC, which is participating in the National BioResource Project of the MEXT, Japan. The authors want to thank Hanne Mordhorst for excellent technical assistance and Henrik Hasman for help on the sequence analysis.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.