
Abstract
Acinetobacter baumannii is emerging as a pathogen that is commonly involved in nosocomial infections. A. baumannii has exhibited the ability to develop multidrug resistance (MDR), including resistance to carbapenems, the last-line class of antibiotics to treat these infections. In particular, MDR A. baumannii International Clone (IC) 2 has disseminated worldwide causing substantial problems in hospitals, including in Asia and Oceania. The global spread of this clonal lineage emphasizes the importance of tracking molecular epidemiology to obtain greater understanding of the population dynamics of A. baumannii. Carbapenem resistance in A. baumannii occurs mainly as a result of acquisition of OXA-type carbapenemase genes, and to some extent by acquisition of metallo-β-lactamase genes. The acquisition of carbapenemase genes, particularly the blaOXA-23, blaOXA-40, and blaOXA-58, by specific clonal lineages may be one of the attributes responsible for the relative homogeneity of the MDR A. baumannii population.
Introduction
A
Due to high similarities of phenotypic and genotypic characteristics, it is difficult to identify Acinetobacter spp. into their species level. 6 Available automated systems used in routine diagnostic laboratories, such as API-20NE, Vitek 2®, and Phoenix, are not able to distinguish the species among the ACB complex. Furthermore, phenotypic and genotypic methods, such as DNA-DNA hybridization and amplified rRNA gene restriction analysis are time-consuming.7,99 Sequencing methods of rpoB and 16S–23S rRNA gene spacer region are reliable for such species identification.11,55 A rapid PCR-based method for gyrB is another reliable tool for species differentiation among the ACB complex. 33 In this review, we verified that publications within the literature had applied proper methods for species identification. The term “Acinetobacter spp.” in this review was used where the methods could not identify species level of Acinetobacter spp.
Within the last few decades, A. baumannii has alarmingly emerged as one of the most important nosocomial pathogens. Infections caused by A. baumannii as a nosocomial pathogen includes ventilator-associated pneumonia, bloodstream infection, wound infection, and meningitis. There are few therapeutic options that can overcome this organism. 77 Carbapenems are one such remaining option as the last-line drugs for treatment of infections caused by A. baumannii. Unfortunately, resistance to carbapenems has become common among A. baumannii worldwide. It is worth noting that A. baumannii is naturally resistant to several antibiotics such as ampicillin, narrow-spectrum cephalosporins, trimethoprim, and ertapenem. 56 A high prevalence of carbapenem-resistant A. baumannii is observed in several geographic areas such as Asia-Pacific (APAC), Latin America, and the Mediterranean.15,35,44,96 The SENTRY antimicrobial surveillance program has shown a continuing decrease in the imipenem susceptibility rates among Acinetobacter spp. collected from different regions (Fig. 1).

The susceptibility rate to imipenem in the APAC region declined from 73.7% in 2001–2004 to 37.4% in 2009 (Fig. 1).26,27 This high rate of carbapenem resistance was observed in most countries from this region. Moreover, pandrug-resistant A. baumannii isolates (resistant to all available class of antibiotics) have also been increasingly identified in Asia.4,16 This highlights the increasing trend of antibiotic resistance in A. baumannii in the APAC region, especially resistance to carbapenems.
Molecular Typing Methods for A. baumannii
A number of molecular typing methods have been used to understand the molecular epidemiology of A. baumannii. The most commonly used typing techniques include (1) DNA-based fingerprinting methods, pulsed-field gel electrophoresis (PFGE), 85 amplified fragment length polymorphism analysis, 21 and multiple-locus variable-number tandem-repeat analysis 96 and (2) PCR-based and sequencing methods, repetitive sequence-based PCR (rep-PCR) analysis, 32 multilocus sequence typing (MLST),5,20 and sequence-based typing and their allele-specific multiplex PCRs. 95 Whole-genome sequencing has also been used for this purpose.1,87
PFGE or restriction analysis of chromosomal bacterial DNA is still currently considered as the gold standard for epidemiologic typing and used in numerous A. baumannii outbreak studies showing high discriminatory power. 85 However, PFGE is laborious, and thus, it is not a suitable technique for rapidly resolving the molecular epidemiology of outbreaks and is not suitable for comparing typing results between laboratories. Amplified fragment length polymorphism (AFLP) analysis has been used as a reference method to identify outbreak clones in Europe, initially named European clones I, II, and III, which had later spread worldwide21,98 and are now known as worldwide clone, global clone, or international clone (IC) 1, 2, and 3.22,32 Up to now, there is no consensus regarding the terminology. In recent literature, A. baumannii outbreak strains have been designated as international clones over others, such as worldwide clone or European clone.20,113 Therefore, “international clone” (IC) will be used in this review.
MLST is an objective method of typing microorganisms that are suitable for population-based studies and global epidemiologic analysis as it allows one to place tested isolates among global isolates registered in centralized A. baumannii MLST databases.5,20 This method is based on comparison of the sequences of internal fragments of seven housekeeping genes. 5 Two MLST schemes are available for Acinetobacter. The first MLST scheme was established by Bartual et al. and is maintained at PubMLST (http://pubmlst.org/abaumannii/). This scheme uses gltA, gyrB, gdhB, recA, cpn60, gpi, and rpoD genes.5,106 The second MLST scheme is maintained at the Pasteur Institute's MLST database (www.pasteur.fr/recherche/genopole/PF8/mlst/Abaumannii.html). The method uses cpn60, fusA, gltA, pyrG, recA, rplB, and rpoB genes, sharing three of them with the PubMLST scheme. 20
In Asia and Australia, PubMLST is used more frequently, while groups in Europe recently prefer to use the Pasteur Institute's scheme.19,28,39 Both MLST schemes were applicable to non-baumannii Acinetobacter spp.104,106 However, the Pasteur MLST database has more designated STs for non-baumannii species and strains than PubMLST. This suggests that Pasteur MLST may be suitable for global epidemiology study of non-baumannii Acinetobacter spp. We have performed a population study on multidrug resistance (MDR) A. baumannii on a global scale using the PubMLST scheme. Our results showed that clonal complex 92 (CC92) was the largest and most geographically diverse CC, which corresponded to IC 2 based on previous typing methods, that is, AFLP and rep-PCR. 83
Multiplex PCR is an alternative method to differentiate clonal lineages of A. baumannii. This method is a sequence-based typing method for three specific genes: ompA, csuE, and blaOXA-51-like which was developed by Turton et al. 95 Owing to higher numbers of polymorphism found in these three genes, this approach has the possibility for greater discrimination than schemes based on housekeeping genes such as MLST.94,95 Through this method, 96 carbapenem-resistant A. baumannii isolates from hospitals in 17 European countries were characterized. Seven different sequence groups (SGs) were identified, and three of these (SG 1, 2, and 3) corresponded to IC 2, 1, and 3, respectively. The majority of isolates belonged to IC 2 and 1, and the remainder belonged to four novel groups. 94 A group in Australia also used this method to characterize Australian isolates, where the majority of isolates belonged to IC 2 and 1. 28
rep-PCR is another useful typing method, one of which is a semiautomated and commercially available method called DiversiLab™ (BioMérieux). This method utilizes a microfluidic LabChip on a bioanalyzer to analyze the amplicons and uses built-in software to build the dendrograms. This method offers the advantages of rapid turnaround time, ease of use, and the ability to maintain libraries of all typed isolates that can be valuable for a future epidemiologic study. Higgins et al. investigated a large global cohort of imipenem-nonsusceptible A. baumannii isolates from 32 countries using the DiversiLab system. 32 The result showed the presence of at least eight distinct clonal clusters, which were assigned as worldwide (WW) lineages 1–8. Three of these clusters represented IC 1, 2, and 3 (WW 1, 2, and 3, respectively) with a predominance of IC 2. Higgins et al. also showed results comparable to those generated by the DiversiLab system and those from PFGE and MLST.83,84 This suggests that the DiversiLab system is one of the more reliable techniques for the global epidemiologic study of A. baumannii.
Molecular Epidemiology of A. baumannii in Asia and Oceania
The molecular epidemiology of A. baumannii in Asia and Oceania has been studied with increasing intensity in recent years using various typing methods. MLST has been most frequently used. To better understand the molecular epidemiology of A. baumannii in Asia and Oceania, here, we have generated a snapshot of the A. baumannii population structure from Asia and Oceania available in the PubMLST database. This snapshot was generated using eBURSTv3 analysis (http://eburst.mlst.net/v3/enter_data/single/) (Fig. 2). We included A. baumannii isolates from nine countries: Australia (n=51), China (n=125), India (n=22), Japan (n=33), South Korea (n=71), Malaysia (n=2), Singapore (n=4), Thailand (n=12), and Vietnam (n=11).
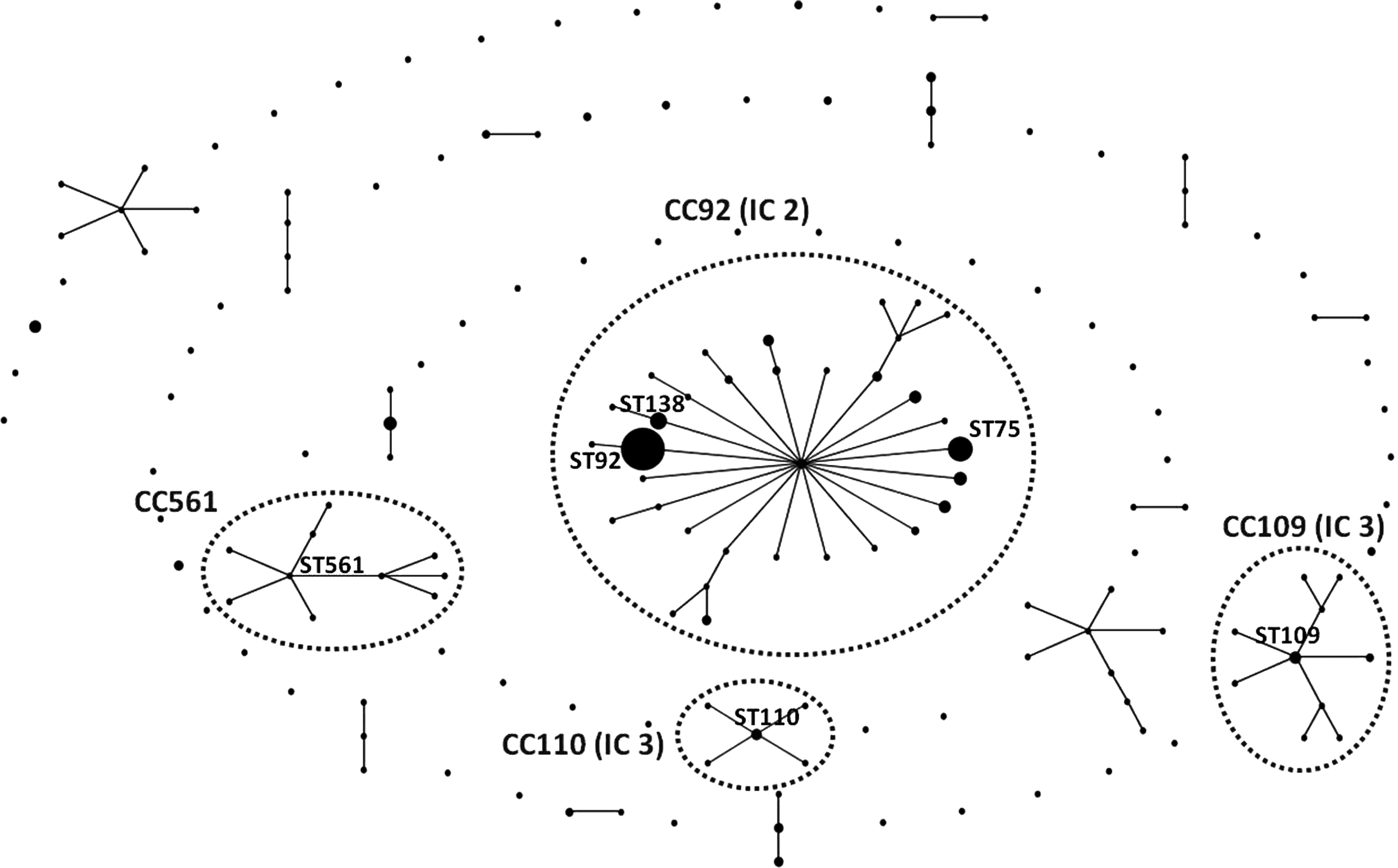
A population snapshot of Acinetobacter baumannii isolates from Asia and Oceania (data from PubMLST). The number of epidemic and clonal complexes (CCs) with corresponded international clones (ICs) is indicated (dash line). The circle represents an ST. The relative size of the circles indicates their prevalence in the PubMLST database. The lines connecting each circle represent single locus variant STs that differ in only one of the seven housekeeping genes.
A total of 331 A. baumannii isolates from 170 different STs were available in the PubMLST database from Asia and Oceania. These were grouped in 17 CCs and 68 singletons (Fig. 2). The most predominant CC was CC92 (n=161, 48.6%), which included isolates from all countries except India. CC92 corresponds to IC 2. Among CC92, ST92 (n=72), ST75 (n=23), and ST138 (n=10) were the most identified STs in the PubMLST database. ST92 was recovered from Australia, China, Japan, South Korea, and Thailand from 2000 to 2010.
Additionally, we have sought additional ST information from publications from the region available in PubMed. ST92 was also found in India, the Philippines, and Taiwan, however, these data were not available through the PubMLST database. 50 ST75 (also part of CC92) was reported as a major ST that caused outbreaks in hospitals in China and South Korea.17,60,74,114 A study from South Korea demonstrated that ST75 and ST138 have increased in 2008–2010. 74 The isolates of these two STs showed higher carbapenem resistance rates than isolates of ST92, which was the most prevalent ST in 2003–2007.
In Asia and Oceania, IC 1 and IC 3 are relatively uncommon compared to IC 2. CC109 (n=15), which corresponds to IC 1, was the second most common CC in Asia and Oceania. More than 50% of the isolates clustered in this CC were recovered from Australia. ST109 was the predicted founder, which was first reported from Australia in 1997. This ST was also found in Japan and South Korea. 89 CC110, which corresponded to IC 3, consisted of eight isolates from South Korea (three), India (two), Australia, Japan, and Vietnam. CC561, a newly described CC has been found in bloodstream isolates in South Korea (2010–2011) and not associated with IC 1, IC 2, or IC 3. Furthermore, A. baumannii can be isolated from feces of pigs and cattle. However, none of these strains was associated with IC 1, IC 2, or IC 3, which are the common clones of A. baumannii strains in humans. 30 This shows that lineages of A. baumannii from animals and humans are distinct.
In conclusion, a few major clonal lineages, including IC 1, 2, and 3, were responsible for the spread of A. baumannii in Asia and Oceania. CC92 (corresponding to IC 2) was the most prevalent clonal lineage recovered from this region (48.6%). There was an emergence of a single CC, CC561, which was found only in South Korea (Fig. 2). Even though CC92 is the most commonly identified clone over a decade, a shift of STs into more carbapenem-resistant ones was observed within CC92. This suggests that carbapenem resistance may be responsible for an evolution within successful clonal lineages, which may lead to a decrease in the heterogeneity of the A. baumannii population.
Mechanisms of Carbapenem Resistance in A. baumannii
Several molecular mechanisms are responsible for conferring carbapenem resistance in A. baumannii. The most common mechanism of carbapenem resistance is the production of carbapenem-hydrolyzing β-lactamases; oxacillinases (OXA-type carbapenemases) and metallo-β-lactamases (MBLs). Alteration of penicillin-binding proteins and loss of outer membrane proteins,18,42,69,100 efflux pump mechanisms, and other β-lactamases are also commonly implicated in carbapenem resistance of A. baumannii. 100
Oxacillinases
The name oxacillinase refers to the ability to hydrolyze the isoxazolyl penicillin, oxacillin, much better than the classical penicillins. 10 The group of OXA-type carbapenemases exhibits carbapenem-hydrolysing activities, in contrast to most OXA-type β-lactamases which do not hydrolyse carbapenems. Most of these enzymes are weak carbapenemases. However, when overexpressed they are able to confer carbapenem resistance. 24
To date, several variants of acquired type OXAs found in A. baumannii have been identified (Fig. 3A). Four major subgroups of OXA-type carbapenemases include both (1) acquired types (OXA-23, OXA-40, and OXA-58 like), where their genes have been found either in the chromosome or plasmid of some but not all A. baumannii isolates and (2) the naturally occurring chromosomal OXA-51-like carbapenemase. 97 Additional groups of acquired OXA-type carbapenemases such as OXA-143 and OXA-235 like have also recently been identified in A. baumannii.34,35

The acquisition mechanisms for OXA-encoding genes in A. baumannii are not thoroughly understood. The blaOXA-23-like and the blaOXA-40-like genes have been reported to be encoded on both the chromosome and plasmids.24,68,77,102 The blaOXA-58-like genes were frequently identified as plasmid mediated. Multiple genomic studies have revealed the pivotal role of insertion sequence (IS) elements. The presence of ISAba1 upstream of blaOXA-23-like, blaOXA-235-like as well as intrinsic chromosomal blaOXA-51-like provides promoter sequences leading to overexpression of these downstream genes conferring resistance to carbapenems.34,81,97 ISAba1, ISAba2, ISAba3, and IS8 have been shown to enhance the expression of blaOXA-58. 82 The overexpression of blaOXA-40-like and blaOXA-143-like is not associated with IS elements and their native promoters may be sufficient for overexpression.92,111
Metallo-β-lactamases
MBLs, Ambler class B enzymes, bear two zinc ions in their active sites and are capable of hydrolyzing all β-lactam antibiotics except aztreonam. IMP, VIM, SIM, and NDM, a recently described MBL, have been identified in Acinetobacter spp. from Asia and Oceania.76,81,101 Among these MBLs, IMP was the most commonly reported in A. baumannii until 2007. However, NDM has often been reported in various Acinetobacter species since its emergence in 2009.101,110 Generally, MBLs are less commonly found in A. baumannii than the OXA-type carbapenemases, most of which, with the exception of NDM, are captured by integrons as gene cassettes. NDM in comparison is found to mobilize through a composite transposon, Tn125, among Acinetobacter spp. 79 Furthermore, A. baumannii has also been theorized to be the platform for the origin of NDM. 93
A rarely reported MBL, the GIM-1 enzyme, has been recently reported in A. pittii isolates in Germany, but has not yet been recovered in Asia and Oceania. 44
Distribution of OXAs and MBLs in Asia and Oceania
The distribution of acquired type OXA and MBL genes is very diverse (Table 1). The acquisition of the blaOXA-23-like has become the most common cause of carbapenem resistance in A. baumannii in Asia and Oceania. 50 OXA-23 was initially described in 1985 as ARI-1 75 and is the most widely spread acquired OXA carbapenemase in this region and worldwide. Other variants of OXA-23, such as OXA-27 (Singapore), OXA-49, OXA-146, OXA-422, and OXA-423 (China), were identified in carbapenem-resistant A. baumannii isolates (Table 1).2,103 OXA-165 to OXA-171 were reported from Thailand (accession no. HM488986 to HM488992). OXA-23-like enzymes have also been found in A. pittii, A. nosocomialis, and A. calcoaceticus.51,52 Several variants of OXA-23 like (OXA-23, OXA-102, OXA-103, OXA-105, OXA-133, and OXA-134) were found on the chromosome of Acinetobacter radioresistens suggesting that A. radioresistens is the progenitor of the blaOXA-23-like genes.66,80
AU, Australia; BA, Bangladesh; CH, China; JP, Japan; HK, Hong Kong; IN, India; KO, South Korea; NP, Nepal; PK, Pakistan; SP, Singapore; TH, Thailand; TW, Taiwan; VT, Vietnam.
MBL, metallo-β-lactamases.
The prevalence of the OXA-40-like cluster causing carbapenem resistance in Acinetobacter spp. from Asia and Oceania is low. OXA-72 was first identified in A. baumannii from Thailand in 2004 (GenBank accession no. AY739646). This enzyme was recovered from A. baumannii isolates from India and southern Taiwan, where OXA-72-producing A. baumannii has been reported to cause hospital outbreaks.54,63,65 OXA-40-like carbapenemases have been found in non-baumannii species such as A. pittii and A. nosocomialis.15,103
OXA-58-like enzymes comprise OXA-58, OXA-96, OXA-97, OXA-164, and OXA-420. OXA-58 in A. baumannii was detected across Asia and Oceania, including Australia, China, India, Singapore, South Korea, and Taiwan (Table 1). Interestingly, the ratio of OXA-58-like enzymes among carbapenem-resistant Acinetobacter was above 40% from certain hospitals in Taiwan.40,61 OXA-96, which differs from OXA-58 by one amino acid substitution, was only described in an A. baumannii isolate from Singapore in 2006. 51 OXA-420 has been recently detected in A. baumannii from Nepal (GenBank accession no. AB983359). Of note, OXA-58-like enzymes were frequently identified in A. pittii isolates from Singapore and Taiwan.40,51,52
OXA-182, a member of OXA-143-like enzymes, has emerged in South Korea. 49 This enzyme shares 93% identity with OXA-143, which is reported to be highly prevalent in Brazil.35,67 The study from South Korea showed that OXA-182 was identified in imipenem-nonsusceptible A. baumannii isolates recovered as early as 2002 and in A. nosocomialis from year 2004 to 2007. 49
OXA-51-like enzymes are intrinsic for A. baumannii and have also been recovered in Taiwan from A. nosocomialis.9,62 OXA-51 like is the most diverse group of oxacillinase with more than 100 variants identified.24,78 Furthermore, the sequences of OXA-51-like genes showed a correlation with their epidemiologic grouping, that is, OXA-69, OXA-66, OXA-71, OXA-51, OXA-65, OXA-64, and OXA-68 correspond to IC 1, 2, 3, 4, 5, 7, and 8, respectively.25,112 OXA-51-like enzymes that were identified in Asia and Oceania are shown in Fig. 3B. OXA-66 was found in Australia, China, Hong Kong, India, Japan, Singapore, South Korea, and Taiwan.23,25,51,64,72,103 OXA-138 and OXA-194 to OXA-197 recovered from A. nosocomialis in Taiwan were also clustered with OXA-66. OXA-69 identified in Australia, India, and Pakistan was grouped with OXA-371 from Nepal. 25 OXA-51 was detected in Japan and India. 23 OXA-68 (Hong Kong and Singapore) 8 was clustered with OXA-144 (Pakistan) and OXA-426 (China). OXA-64 was only reported from Singapore. 29 There was no report of OXA-65 and OXA-71 in Asia and Oceania.
Even though MBLs are less frequently identified in A. baumannii in Asia and Oceania, a relatively high prevalence of MBL-producing Acinetobacter spp. isolates was observed in some countries, such as India and South Korea.58,73,86 Several variants of IMP-type enzymes were identified in Japan, including IMP-1, IMP-11, and IMP-19 (Table 1). IMP-2 was identified in A. baumannii isolates from a hospital in India. 73 This enzyme shares 84.9% amino acid identity with IMP-1. IMP-4, which had 95.6% amino acid identity with IMP-1, was first described in Acinetobacter spp. isolates from Hong Kong 14 and was subsequently reported from Australia and Singapore as well.51,76 IMP-8 and IMP-14 were only identified in Taiwan and Thailand, respectively.46,59
VIM-like enzymes were sporadically reported from India, South Korea, and Taiwan.49,59,73 SIM-1 was identified in China and South Korea.58,115 NDM-1 was recently recovered from Acinetobacter spp. isolates from India, Bangladesh, Pakistan, Japan, and China. 101 These isolates from China included nonclinical samples recovered from hospital sewage and meat-producing animals.105,117
MBLs were identified in several Acinetobacter species, including A. baumannii, A. baylyi, A. bereziniae, A. calcoaceticus, A. haemolyticus, A. johnsonii, and A. junii, A. lwoffii, A. nosocomialis, and A. pittii.53,59,76,88,101,107,108,115 MBLs identified in Acinetobacter spp. are listed in Table 1.
Conclusion
Carbapenem resistance in A. baumannii has had a drastic increase in Asia and Oceania within the last decade. The increase of carbapenem resistance in non-baumannii Acinetobacter spp. has also been observed. Production of OXA carbapenemases and MBLs is the main mechanism of carbapenem resistance in A. baumannii. OXA-23-like enzymes are the most commonly identified carbapenemases in A. baumannii from Asia and Oceania. Other groups of OXA carbapenemases and MBLs are sporadically reported from different countries. It is worth noting that NDM can also be isolated from meat-producing animals in China. Thus, the spread of NDM-producing Acinetobacter may be hastened by its appearance in the food chain and subsequently become a real threat in Asia and Oceania.
CC92, which corresponds to IC 2, is the most successful and widely disseminated clone of A. baumannii in Asia and Oceania. Carbapenem resistance may be one of the adaptive mechanisms responsible for a decrease in heterogeneity among the A. baumannii population. To address this increase of carbapenem resistance Acinetobacter spp., proper species identification and typing methods are essential for early detection and monitoring in epidemiology investigations. Factors that have contributed to the ability of Acinetobacter spp. to spread and persist in the hospital environment have yet to be determined. The genome-wide approach may provide such insight for prevention and control of further Acinetobacter spp. transmission.
Footnotes
Acknowledgment
W.K. is a Research Higher Degree candidate funded by Siriraj Hospital, Mahidol University.
Disclosure Statement
D.L.P. has been on the advisory boards for Merck, AstraZeneca, Cubist, Bayer, and Pfizer.