
Abstract
CTX-M-15 is a chief contributor for expanded-spectrum cephalosporin and monobactam resistance in India, complicating treatment options. In this study, we have investigated genetic context of CTX-M-15 in Escherichia coli and their transmission dynamics in a tertiary referral hospital of India. A total of 198 isolates were collected, of which 66 were harboring blaCTXM-15. Among them, 14 isolates were carrying a single CTX-M-15 gene and 52 were harboring multiple extended-spectrum β-lactamase genes along with blaCTX-M-15. The resistance gene was flanked by tnpA, ISEcp1, IS26, and ORF477 in 10 different arrangements. The resistance determinant was horizontally transferable through F, W, I1, and P Inc types of plasmids. Restriction mapping of plasmids showed a variable band pattern even within the same Inc types. Minimum inhibitory concentration was found above the breakpoint level against expanded-spectrum cephalosporins and monobactam while susceptible against carbapenems. blaCTX-M-15 was highly stable and sustained in the cell after 115 serial passages. In pulse-field gel electrophoresis, eight pulsotypes of E. coli were found to be responsible for the spread of blaCTX-M-15 in the tertiary referral center. We conclude that the presence of CTX-M-15 in the heterogeneous group of E. coli is highly alarming in terms of infection control and it may require regular monitoring, so as to formulate appropriate antibiotic policy to stop the spread of this resistance determinant.
Introduction
C
This study was undertaken to investigate the genetic context of CTX-M-15 within E. coli, their horizontal and vertical transferability in a tertiary referral hospital of India.
Methodology
Bacterial strain
A total number of 198 consecutive nonduplicate isolates of E. coli were collected from different wards/clinics of Silchar medical college and hospital, India, for a period of 1 year (i.e., May 2012–April 2013).
Phenotypic detection and molecular characterization of ESBLs
All isolates were subjected to an initial screening using cefotaxime and ceftazidime (Hi-Media) at a 1 mg/L concentration and were confirmed by the combined disc diffusion method through cefotaxime and ceftazidime alone and in combination with clavulanic acid (Hi-Media) as per Clinical Laboratory Standard Institute (CLSI) criteria. 7 Multiplex polymerase chain reaction (PCR) was carried out for detection and characterization of blaESBL genes using five pairs of primers targeting blaCTX-M, blaTEM, blaPER, blaSHV, and blaOXA-2.3,15
Analysis of clonal variation and sequencing
Denaturing gradient gel electrophoresis (DGGE) was performed with amplified PCR products of blaCTX-M with DCode™ Universal Mutation Detection System (Bio-Rad). PCR products (5 μL) of blaCTX-M were loaded onto a polyacrylamide gel with 40–60% denaturant gradients. After electrophoresis, the gel was stained for 10 minutes within ethidium bromide and then washed in distilled water for 20 minutes. Gel image was acquired using a Gel Doc™ EZ imager (Bio-Rad). The representative of each band pattern was sequenced. The nucleotide sequences were analyzed using the software available on the National Center of Biotechnology Information website (www.ncbi.nlm.nih.gov).
Investigation of genetic environment of blaCTX-M
To determine linkage with insertion sequences, PCR was performed with IS26, tnpISEcp1, ISEcp1, tnpIS26, and CTX-M primers using the following reaction condition: initial denaturation 95°C for 2 minutes, 35 cycles of 95°C for 20 seconds, 50°C for 45 seconds, 72°C for 2 minutes, and a final extension at 72°C for 7 minutes. The primers used as described earlier. 10 Amplified products were purified and cloned on pGEM-T vector (Promega) and sequenced as well. To screen the presence of integron in the ESBL-producing organism, integrase gene PCR was carried out. 13 Furthermore, association of blaCTX-M with integron was assessed by performing two PCRs: in one reaction 5′-CS and CTX-M-R′ and in another reaction 3′-CS and CTX-M-F′ were used.13,15
Plasmid preparation, genetic transferability, and incompatibility typing
Bacterial isolates were grown overnight in Luria Bertani broth (Hi-Media) containing 0.5 μg/ml of cefotaxime, and plasmids were extracted using the QIAGEN® Plasmid Mini Kit (Germany). Transformation was carried out using E. coli JM107 and selected on LB agar plates containing cefotaxime 0.5 mg/L. The conjugation experiment was performed between the clinical isolates and streptomycin-resistant E. coli recipient strain B (Genei). Overnight, culture of the bacteria was diluted in Luria Bertani Broth (Hi-Media) and was grown at 37°C till the OD of the recipient and donor culture reached 0.8–0.9 at A600. Donor and recipient cells were mixed at 1:5 donor-to-recipient ratios, and transconjugants were selected on cefotaxime (0.5 μg/ml) and streptomycin (800 μg/ml) agar plates. The conjugation frequency was determined as the number of exoconjugants per input donor cell, and results were the average of three independent experiments per strain. 9 Incompatibility groups were determined by PCR-based replicon typing 6 among all the transformants and transconjugants.
Cloning of CTX-M-15 whole gene including promoter regions
For amplification of whole blaCTX-M gene, including promoter regions, primer pairs were designed and PCR was carried out with the designed primers of CTX-M-F′ 5′-CATTGCAGCAAAGATGAAATC-3′ and CTX-M-R′ 5′-CAGCGCTTTTGCCGTCTAAGG-3′gene, respectively. Amplified products were purified and cloned on the pGEM-T vector (Promega) and sequenced further.
Restriction mapping of plasmids and carriage of blaCTX-M
Restriction mapping of plasmids harboring blaCTX-M was carried out. Plasmids were digested using HindIII, XbaI, and EcoRI (Promega) and were separated by pulse-field gel electrophoresis (PFGE) (CHEF DR III system; BIO-RAD). Furthermore, Southern blotting was performed on agarose gel by in-gel hybridization with the blaCTX-M probe labeled with the Digoxygenin High Prime Labeling Mix (Roche) Detection Kit. Plasmid DNA was separated on agarose gel and subjected to transfer to the nylon membrane (Hybond N) and hybridized with a prepared blaCTX-M-specific probe. Detection was performed using an NBT color detection kit (Roche).
Susceptibility testing of donor strains, transformants, transconjugants, and CTX-M clones
Antimicrobial susceptibility was determined by Kirby–Bauer disc diffusion method on Muller-Hinton Agar (Hi-Media) plates. 7 The following antibiotics were used: amikacin (30 μg), ciprofloxacin (30 μg), gentamicin (10 μg), tigecycline (15 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg), and cefoxitin (30 μg) (Hi-Media). Susceptibility of tigecycline was interpreted as per FDA breakpoints. 12 Minimum inhibitory concentration (MIC) with cefotaxime, ceftazidime, ceftriazone, cefepime, imipenem, meropenem, ertapenem, and aztreonam (Hi-Media) was also determined by the agar dilution method. 7 The results were interpreted as per CLSI recommendation. However for tigecycline, the organism showing an inhibition zone less than 20 mm was considered resistant. 7
Plasmid stability testing
Plasmid stability analysis of a total of 66 numbers of blaCTX-M producers in donor strains and transformants was performed by the serial passages method for consecutive 110 days at 1:1,000 dilutions in LB broth without antibiotic pressure. After each passage, 1 ml of the culture was diluted into 10−3 dilution with normal saline, and 50 μl of the diluted sample was spread on to the LB agar plate. After overnight incubation, 50 colonies from plates were randomly picked and subjected to PCR assay for blaCTX-M. The template DNA was prepared from bacterial suspension by the boiling centrifugation method. Approximately 105 CFU were taken for preparation of template DNA, and the PCR assay was carried out for the presence of the blaCTX-M. 17
Strain typing
Genotyping of all the E. coli isolates harboring blaCTX-M was done by digestion of agarose plugs with XbaI and analyzed by PFGE (CHEF DRIII system; BIO-RAD).
Results
A total of 66 isolates were observed carrying blaCTX-M. Of which, 14 were carrying blaCTX-M only (Table 1), whereas in the rest of the 52 isolates, cocarriage of multiple ESBLs was observed along with blaCTX-M (Table 2). Most of the isolates obtained from the surgery unit followed by medicine, gynecology, and pediatrics wards were isolated from pus, urine, throat swab, as well as sputum, and recovered from different age groups of patients (Table 1). DGGE showed an identical banding pattern of blaCTX-M in all isolates, on sequencing it was confirmed as a blaCTX-M-15 variant. Sequencing results also confirmed that blaCTX-M-15 was associated with mobile genetic elements with 10 different types of genetic arrangements (Fig. 1, Table 1). In all the arrangements, blaCTX-M-15 was flanked by tnpA, ISEcp1, IS26, and ORF477. Additionally, tnpR was in the upstream region of blaCTX-M-15 in arrangement 4. In arrangements 1, 4, and 10, blaCTX-M-15 was in reverse orientation (Fig. 1). All the study isolates were found to carry class I integron although blaCTX-M-15 could not be found to be associated with the gene capture mechanism. blaCTX-M-15 was found to be plasmid mediated and horizontally transferable in all the test isolates. Conjugation frequency of each transconjugant is mentioned in Table 1. Incompatibility typing of plasmids of transformants and transconjugants suggested that they were originated through I1, F (FIA = 6; FIC = 3; FrepB = 3), W, and P Inc groups (Table 1). Hybridization results further supported their transferability through plasmids as all the plasmids were hybridized with a blaCTX-M-15-specific probe. Restriction mapping of plasmids fetched a variable band pattern even within the same incompatibility type. On observing susceptibility results, all the test isolates were found to be susceptible to tigecycline and showed a multidrug-resistant phenotype. MIC was found above the break point against extended-spectrum cephalosporins and monobactams ranging from 64 to 256 mg/L, whereas in the case of carbapenems, MIC was found within the susceptible range (Table 3). Isolates harboring multiple ESBLs in combination with blaCTX-M-15 too showed the same MIC range (Table 2). To predict any other coexisting mechanisms in cephalosporin resistance, MIC of transformants and transconjugants was assessed and compared with the parent strain. The observation was the same MIC value toward all the cephalosporins and monobactam (Table 3). Vertical transfer of blaCTX-M-15 was investigated and found that the resistance determinant was stable till 115 consecutive serial passages without any antibiotic pressure. The proportion of plasmid loss is summarized in Table 4. PFGE results showed eight different pulsotypes in the study (Fig. 2). Pulsotype 5 was predominant as 35 isolates showed this banding pattern. None of the pulsotype could be specifically linked with any of the wards in our clinical setting.
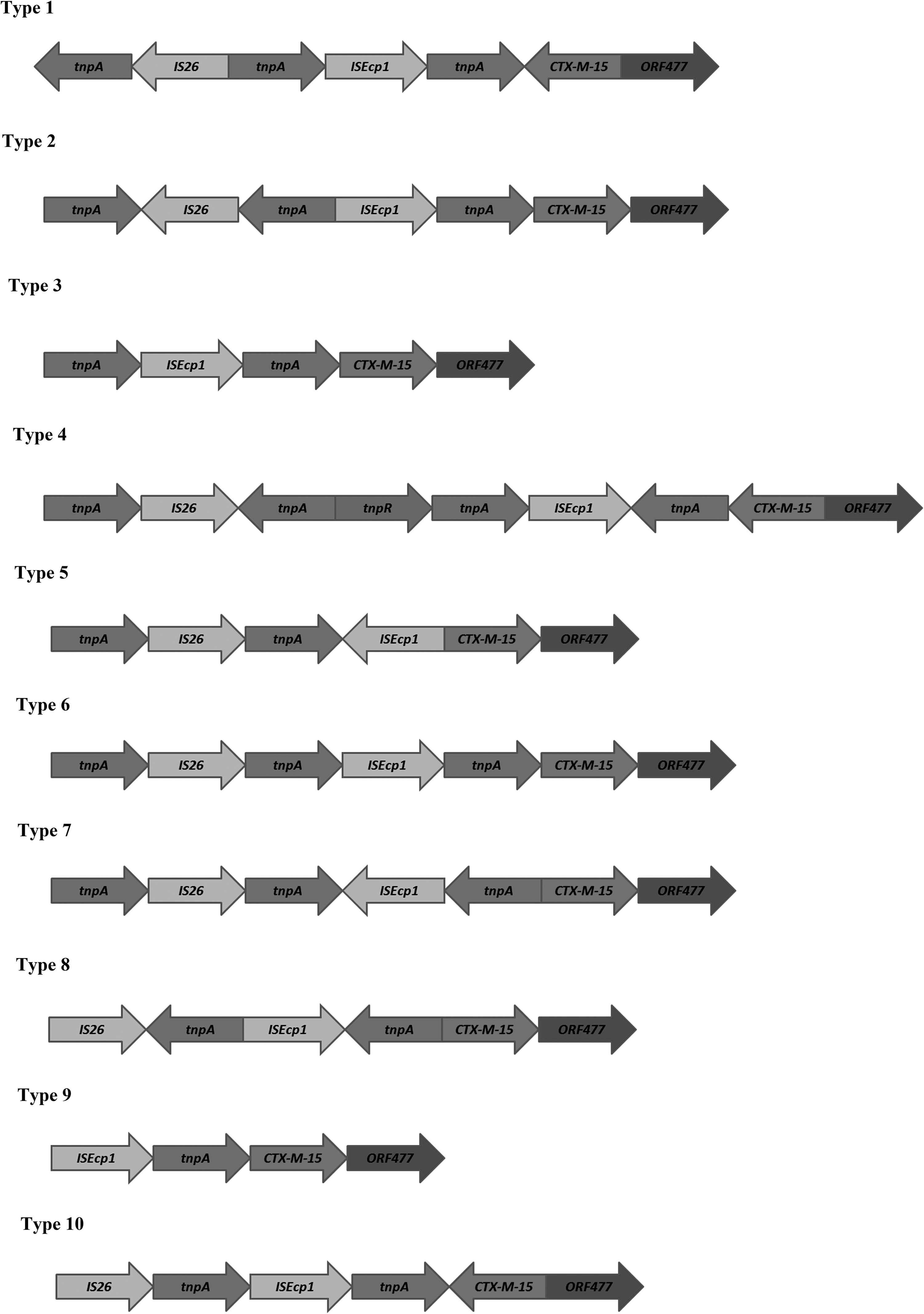
Schematic representation of diverse genetic arrangements of CTX-M-15 genes with mobile genetic elements.
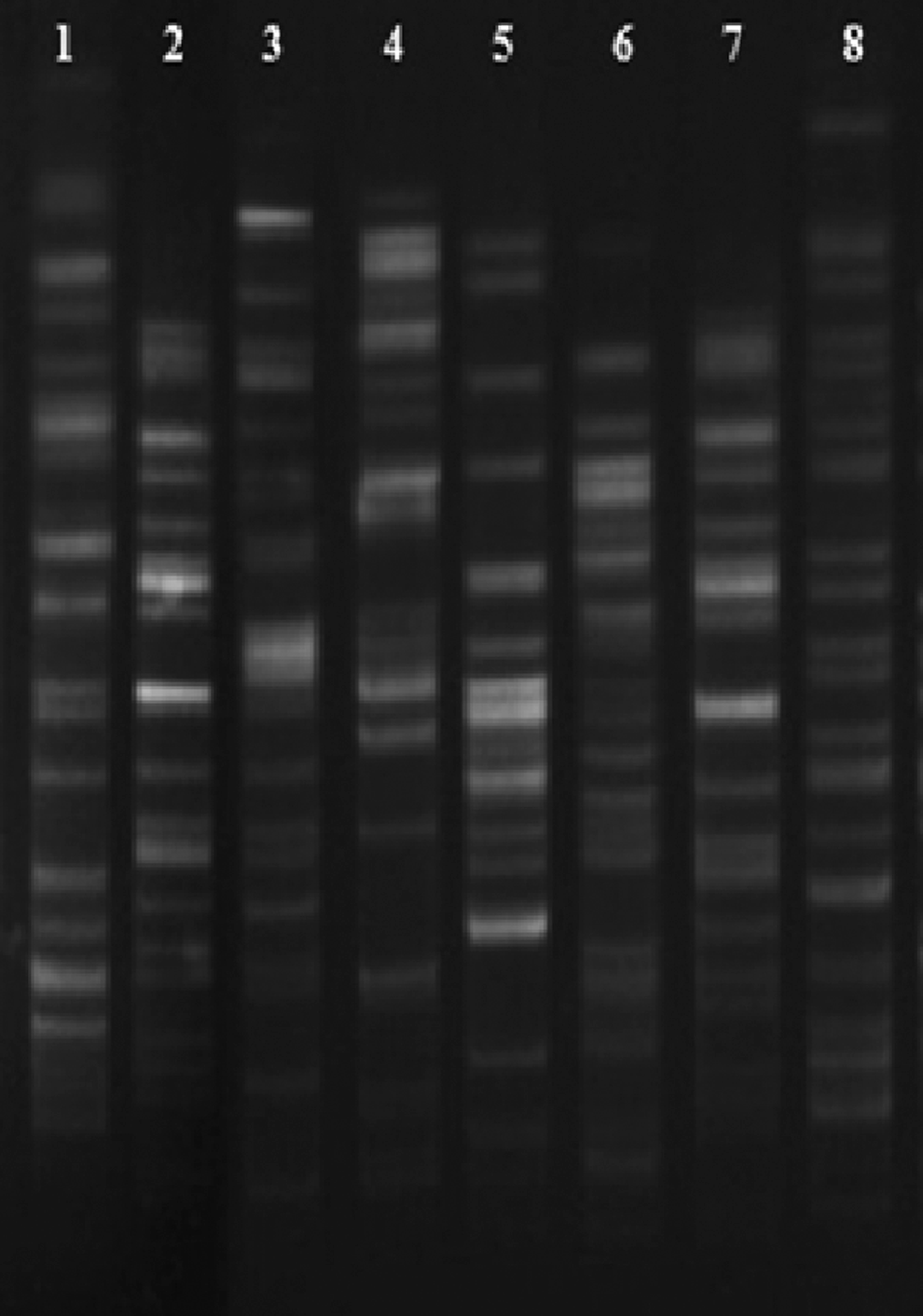
Pulse-field gel electrophoresis (PFGE) pattern of Escherichia coli isolates harboring CTX-M-15 gene digested by XbaI, lanes 1–8: E. coli PFGE pulsotypes 1–8.
Antibiotics: AMK, amikacin; CIP, ATM, aztreonam; ciprofloxacin; FOX, cefoxitin; GEN, gentamicin; SXT, cotrimoxazole.
CAZ, ceftazidime; CPD, cefpodoxime; CRO, ceftriaxone; CTX, cefotaxime; ESBL, extended-spectrum β-lactamase; ETP, ertapenem; FEP, cefepime; IPM, imipenem; MEM, meropenem; MIC, minimum inhibitory concentration.
DS, donor strain; TF, transformants; TC, transconjugants.
PCR, polymerase chain reaction.
Discussion
This study was carried out to determine flanking regions of blaCTX-M-15, the predominant ESBL type in India. The investigation was performed in a single species to understand their clonal lineage. An understanding of their genetic elements gives better insight into mobilization units of resistance gene. A high MIC value against cefotaxime was observed in tranformants and clones, although the value was same against other third generation cephalosporins and monobactam. Amikacin resistance in study isolates could be due to the presence of acquired 16SrRNA methyl transferases or any other mechanisms, which is also noticed in recent studies.23,24 Ten different organizations of blaCTX-M-15 indicate parallel mobilization of the resistance determinant within this tertiary referral hospital. It was observed that blaCTX-M-15 was in reverse orientation in three arrangements (types 1, 4, and 10). However, it did not interfere with cephalosporinase activity as observed with MIC data. Among the 10 different arrangement types, type 4 was most common in study isolates (n = 4). However, all the arrangement types were previously reported elsewhere from the world. In a previous study conducted in France, the arrangement type 4 was reported in Enterobacteriaceae, 10 whereas type 10 was observed in a report from France. 14 Thus, this kind of study of ours put a glimpse on the global distribution of these genetic arrangements and their movement, adaptability, and propagation under antibiotic exposure in different clinical environmental conditions across the globe. Interestingly, in this study it was observed that blaCTX-M-15 was horizontally transferred through multiple incompatible types of plasmids. This proves their diverse source of origin and adaptation in our hospital settings. CTX-M-15 is known to be pandemic and is often reported to be carried within Inc FII plasmid, 8 although there were some reports of their carriage through Inc I1, A/C, L/M, and N types as well. 19 In India, no such data are available and this study is the first of such kind. These plasmids with blaCTX-M-15 were highly stable and maintained within cell without any antibiotic pressure. Thus, they have the potential for expansion in community if dispersed outside the hospital environment in an antibiotic stress-free condition. In the stability analysis, the P Inc type harboring blaCTX-M-15 showed progressive plasmid loss after 40 passages. This implicates the specialized adaptation of this plasmid for survival of host under cephalosporin stress in the hospital environment. None of the earlier studies has attempted to investigate the vertical transmission dynamics of this pandemic resistance determinant and it will be of more interest to compare our data with other variants of CTX-M gene from different geographical regions having varying antibiotic pressure and treatment policy. The presence of this resistance gene in the heterogeneous group of E. coli in this hospital might be due to the extensive use of cefotaxime and ceftriaxone in this setting. Eight pulsotypes were distributed over the hospital, irrespective of their confinement in any of the wards, although pulsotype 5 was predominant of all. This information is quite helpful in infection control, and no such study has been attempted earlier from this part of the country on typing of CTX-M producers.
In conclusion, this is probably the first report from India describing multiple genetic contexts of blaCTX-M-15 and diverse genetic vehicles from a single center. This study insists vital requirement for regular monitoring of this resistant determinant and to execute the right antibiotics policy to reduce the irrational utilization of expended spectrum cephalosporins, and so as to decrease the treatment failure and antibiotic pressure in the clinical setting.
Footnotes
Acknowledgments
The authors would like to acknowledge the help of the HOD, Microbiology, Assam University, for providing infrastructure. They also acknowledge the University grants commission (UGC-MRP), Government of India, and the Department of Bio-technology (DBT-NER, Twinning), Government of India. Authors also acknowledge the help from Assam University Biotech Hub for providing laboratory facility to complete this work.
Disclosure Statement
No competing financial interests exist.