
Abstract
Dissemination of antimicrobial resistance is a major global public health concern. To clarify the role of flies in disseminating antimicrobial resistance between farms, we isolated and characterized tetracycline-resistant Escherichia coli strains isolated from flies and feces of livestock from four locations housing swine (abattoir, three farms) and three cattle farms. The percentages of isolates from flies resistant to tetracycline, dihydrostreptomycin, ampicillin, and chloramphenicol (80.8%, 61.5%, 53.8%, and 50.0%, respectively) and those from animal feces (80.5%, 78.0%, 41.5%, and 46.3%, respectively) in locations housing swine were significantly higher than those from cattle farms (p<0.05). The rates of resistance in E. coli derived from flies reflected those derived from livestock feces at the same locations, suggesting that antimicrobial resistance spreads between livestock and flies on the farms. The results of pulsed-field gel electrophoresis (PFGE) analysis showed that, with a few exceptions, all E. coli isolates differed. Two pairs of tetracycline-resistant strains harbored similar plasmids with the same tetracycline-resistance genes, although the origin (fly or feces), site of isolation, and PFGE patterns of these strains differed. Therefore, flies may disseminate the plasmids between farms. Our results suggest that flies may be involved not only in spreading clones of antimicrobial-resistant bacteria within a farm but also in the widespread dissemination of plasmids with antimicrobial resistance genes between farms.
Introduction
T
Antimicrobial-resistant bacteria can spread through many routes that include not only retail meat, 26 but also the environment in which humans are exposed to livestock. Recently, the role of flies in spreading antimicrobial-resistant bacteria was reported.4,25,27,29 With continuing urban expansion into areas zoned for agricultural use, the public health community has become increasingly concerned about flies associated with animal production. 29
We have shown that flies and cattle feces on a farm carry several clones of Escherichia coli that are resistant to third-generation cephalosporins. 25 Flies move freely between livestock and human residences 8 and may play an important role as vectors of antimicrobial-resistant bacteria, not only within a farm, but also over large distances. Our previous characterization of clonal isolates of E. coli present on a single cattle farm 25 did not demonstrate the role of flies in disseminating antimicrobial resistance.
Therefore, in this study aimed at clarifying the role of flies in disseminating antimicrobial resistance between farms, we focused on tetracycline-resistant bacteria isolated from several farms, which are observed more frequently than bacteria resistant to third-generation cephalosporins. We report here the identification and characterization of tetracycline-resistant E. coli isolated from flies and livestock feces from locations housing swine and from cattle farms.
Materials and Methods
Sample collection
All fecal and fly samples were collected during April 2013 on the main island of Okinawa, Japan, from four locations housing swine, including three farms and an abattoir, and three cattle farms (Fig. 1). The island is an appropriate location to conduct such an epidemiological study. Furthermore, flies are active because of the island's subtropical location. We collected fecal samples from 40 swine and 30 cows (10 samples per location), and 111 flies were collected using a sweep net (64 samples per swine location and 47 samples per cattle farm). The fly species comprised 74 houseflies (Musca domestica; 32 samples per swine locations and 42 samples per cattle farms), 31 greenbottle flies (Lucilia sericata; all from swine locations), and 6 stable flies (Stomoxys calcitrans; 1 sample per swine location and 5 samples per cattle farms), were identified according to morphology using a stereomicroscope.
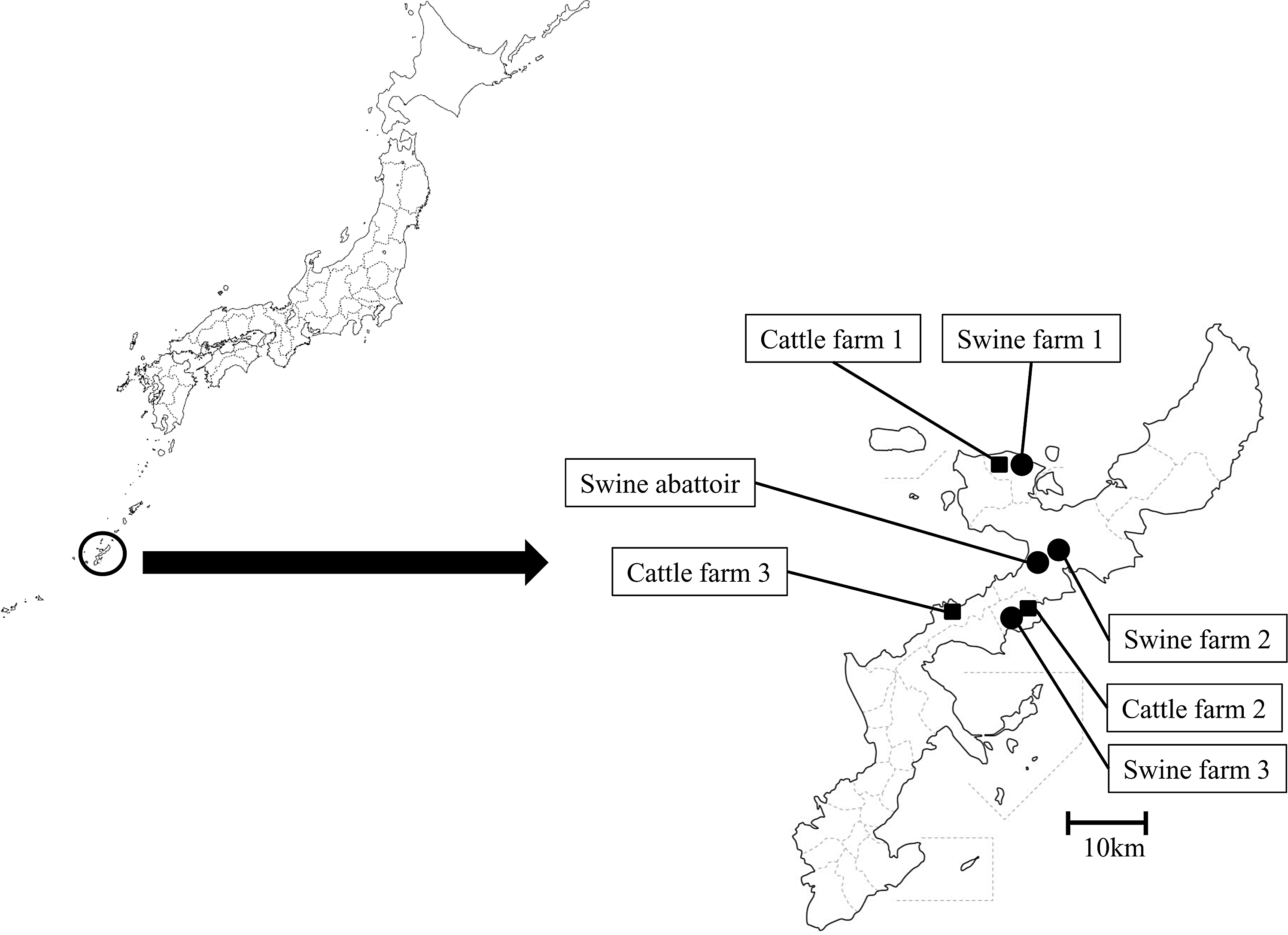
Map of the sampling sites. Sources: http://www.finemakeyuri.com/map.finemakeyuri.com/002.html; http://map.finemakeyuri.com/552.html
Isolation of bacteria
Individual flies were surface-sterilized using sodium hypochlorite and ethanol as described previously, 15 and samples were then washed thrice with sterile distilled water and homogenized in potassium phosphate-buffered saline. The homogenized flies and fecal samples were inoculated onto cotton swabs (Seedswab gamma1; Eiken Chemical, Tokyo, Japan). The swabs were then used to inoculate deoxycholate-hydrogen sulfate-lactose (DHL) agar medium (Nissui Pharmaceutical, Tokyo, Japan) that was incubated at 37°C overnight. For each sample, a maximum of three colonies were identified as E. coli according to colony morphology and analyzed further. The isolates were identified biochemically as E. coli using API20E tests (Sysmex, Kobe, Japan) and verified using polymerase chain reaction (PCR) amplification with primers specific for the E. coli malB promoter. 28 The PCR conditions were the same as previously reported. 28
When two or three isolates from any sample exhibited the same pulsed-field gel electrophoresis (PFGE) type and antimicrobial susceptibility profile, they were considered a single isolate.
Analysis of antimicrobial susceptibility
We determined the minimal inhibitory concentrations (MICs) using the agar dilution method of the Clinical Laboratory Standards Institute (CLSI) guidelines. 7 The antibiotics tested were as follows: ampicillin, cefazolin, cefotaxime, kanamycin, gentamicin, dihydrostreptomycin, tetracycline, chloramphenicol, nalidixic acid, and enrofloxacin (all from Sigma-Aldrich, St. Louis, MO). The resistance breakpoints were defined according to the CLSI guidelines. We determined breakpoints for dihydrostreptomycin, which are not defined by the CLSI guidelines, from a report on the Japanese Veterinary Antimicrobial Resistance Monitoring (JVARM) system. 17 These breakpoints were used to separate susceptible from resistant strains. The antimicrobial-resistance rate was defined as the percentage of antimicrobial-resistant strains among all isolates. E. coli ATCC25922 and Pseudomonas aeruginosa ATCC27853 were used as controls. Tetracycline-resistant isolates (MIC ≥16 μg/ml) were selected for further tests.
Characterization of resistance genes
The presence of tetA, tetB, tetC, tetD, tetE, and tetG 10 in addition to tetM and tetS 13 were determined using multiplex and standard PCR. Primer sequences are listed in Table 1. The PCR conditions were the same as previously reported,10,13 and the tetA and tetB amplicons were subsequently sequenced. The amplified product was purified with FastGene Gel/PCR Extraction Kit (Nippon Genetics, Tokyo, Japan) and sequenced in both directions using the same primers as for PCR. Nucleotide sequences were determined using the BigDye Terminator, version 3.1, Cycle Sequencing kit with an automated DNA sequencer (ABI 3130; Applied Biosystems, Foster City, CA).
TETF was designed to the conserved region of six different tet genes and used as a common sense primer in polymerase chain reaction amplifications.
Pulsed-field gel electrophoresis
The tetracycline-resistant isolates were typed using PFGE according to the Pulse Net CDC protocol. 6 Genomic DNA in each agarose plug was digested with XbaI (Takara Bio, Shiga, Japan). The PFGE procedure was performed using the CHEF-DRIII system (Bio-Rad Laboratories, Hercules, CA) under conditions as follows: switch time, 2.2–54.2 s; running time, 18 h; included angle, 120°; voltage, 6 V/cm; and temperature, 14°C. The PFGE profiles were analyzed using the BioNumerics program (Applied Maths, Sint-Martens-Latem, Belgium). Similarity and diversity were assessed using the Dice coefficient. Cluster analysis was performed using the unweighted-pair group method with arithmetic means (UPGMA).
Transfer of tetracycline-resistance genes and characterization of plasmids
Plasmid transfer was determined using previously described broth-mating assays with slight modifications. 24 Briefly, the recipients were rifampicin-resistant E. coli K12 DH5α strains, and mating was conducted at 37°C. Transconjugants were selected on Mueller–Hinton agar supplemented with 50 μg/ml rifampicin (Sigma-Aldrich) and 16 μg/ml tetracycline. The broth-mating assays were repeated three times. All plasmids harboring tet genes of transconjugants were replicon-typed using a published PCR method.5,11 Plasmid DNAs were purified from parental strains and transconjugants using a modified alkaline lysis method, 12 and plasmid sizes were estimated using the BAC-Tracker supercoiled DNA ladder (Epicentre Biotechnologies, Madison, WI). Southern hybridization was subsequently performed. The plasmids were transferred using downward capillary transfer to Hybond-N+ nylon membrane (GE Healthcare, Little Chalfont, United Kingdom), and the membrane was treated according to standard procedures. 21 Labeling of DNA probes, hybridization, and detection of hybrids were performed using digoxigenin (DIG)-PCR and DIG Nucleic Acid Detection Kits (Roche Diagnostics, Basel, Switzerland) according to the manufacturer's instructions. DNA probes were generated from purified PCR products of tetA, tetB, and the FIB, Frep, and Y gene replicons.
Statistical analysis
The chi-square test was used to compare the antimicrobial-resistance rates among isolates from fecal samples of swine and cattle and to compare the antimicrobial-resistance rates between isolates from flies derived from locations housing swine and from cattle farms.
Results
Isolation of E. coli from flies and feces
E. coli was isolated from 32.8% (21/64) of flies and 65.2% (30/40) of swine feces, respectively, taken from locations housing swine. The fly species were identified as follows: 34.4% (11/32) houseflies, 29.0% (9/31) greenbottle flies, and 100% (1/1) of stable flies. When multiple isolates from the sample exhibited the same PFGE type and antimicrobial susceptibility profile, they were considered a single isolate; thus, 26 and 41 strains, derived from flies and swine feces, respectively, were isolated.
E. coli was isolated from 27.7% (13/47) and 76.7% (23/30) of flies and cattle feces samples, respectively, from cattle farms. We obtained isolates from 31.0% (13/42) of houseflies, but were unable to isolate E. coli from the five stable flies obtained from cattle farms. Again, when multiple isolates from the same sample exhibited the same PFGE type and antimicrobial susceptibility profile, they were considered as a single isolate. Thus, 14 and 37 strains, derived from flies and cattle feces, respectively, were isolated from the cattle farms.
Antimicrobial resistance
The proportions of antimicrobial-resistant E. coli are shown in Table 2. All antimicrobial-resistance rates of isolates derived from the locations housing swine were higher than those from the cattle farms. The proportions of antimicrobial-resistant E. coli derived from flies and livestock feces were similar among farms. The percentages of tetracycline, dihydrostreptomycin, ampicillin, and chloramphenicol resistance of isolates obtained from flies and those obtained from animal feces in locations housing swine were significantly higher than those from cattle farms (p<0.05).
This value represents the CLSI breakpoints, except for dihydrostreptomycin.
The percentages of antimicrobial resistant flies isolated from locations housing swine were significantly higher compared with those in cattle farms (p<0.05).
The percentages of antimicrobial-resistant livestock feces isolated from locations housing swine were significantly higher than those isolated from cattle farms (p<0.05).
The value represents the JVARM breakpoint.
CLSI, Clinical Laboratory Standards Institute; JVARM, Japanese Veterinary Antimicrobial Resistance Monitoring.
Antimicrobial-resistance genes
In this study, 65 tetracycline-resistant strains (21 from swine facility flies, 33 from swine feces, 3 from cattle farm flies, and 8 from cattle feces) were isolated. These 65 isolates were tested for the presence of antimicrobial-resistance genes. Thirty-two (49.2%) and 28 isolates (43.1%) harbored the tetA and tetB genes, respectively. The tetG and tetS genes were detected in one isolate each (1.5%). One isolate that harbored tetA also contained tetD and tetM. Four isolates that harbored tetM also contained tetA. Five isolates that harbored tetB also contained tetM. Tetracycline-resistance genes were not detected in three tetracycline-resistant isolates. The tetC and tetE genes were not detected in any of the tetracycline-resistant isolates.
Sequence analysis revealed no polymorphisms were found in the 32 tetA and 28 tetB nucleotide sequences.
PFGE analysis of tetracycline-resistant E. coli
The genetic relationships among the 65 tetracycline-resistant E. coli isolates determined using PFGE analysis are shown in Fig. 2. The tetracycline-resistant isolates (n=65) generated 60 PFGE patterns (Fig. 2). Five pairs were observed with >80% similarity level. Each of the five pairs had the same tetracycline-resistance genes (tetA or tetB).
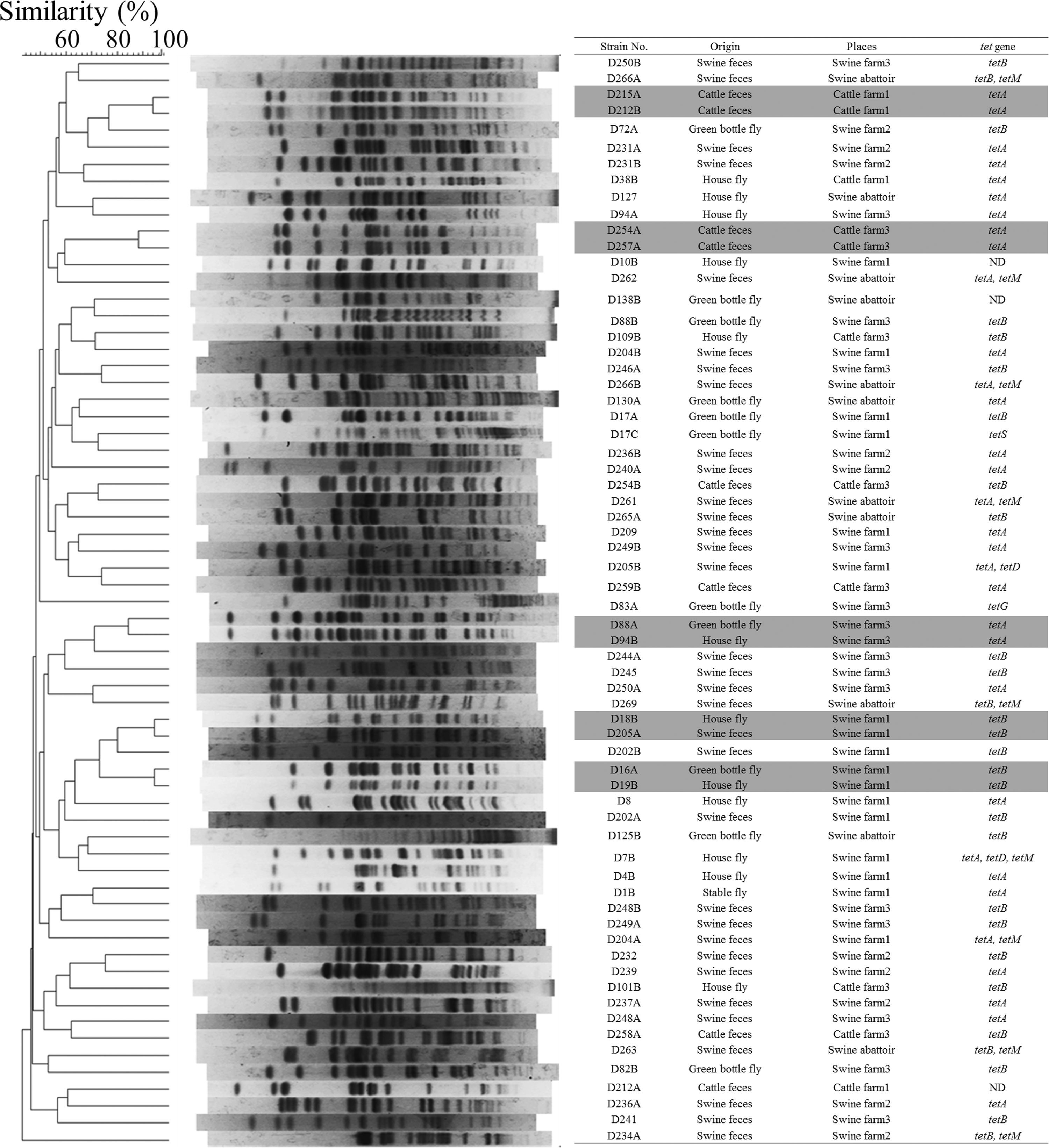
Pulsed-field gel electrophoresis (PFGE) analysis showing percent similarities of 65 tetracycline-resistant isolates. DNA samples for PFGE analysis were digested with XbaI, and isolates with >80% similarity are highlighted.
The PFGE pattern of an isolate derived from a housefly (D18B) from swine farm 1 was 95% similar to that of an isolate derived from swine feces (D205A). The antimicrobial-resistance profiles of these two isolates were the same; they were resistant to dihydrostreptomycin, tetracycline, and chloramphenicol. On swine farms 1 and 3, single isolates derived from a housefly (D19B and D94B) were 95% and 85% similar, respectively, to an isolate derived from a greenbottle fly (D16A and D88A) from each farm. The antimicrobial-resistance profiles of these two pairs of isolates were the same. D19B and D16A were resistant to ampicillin, cefazolin, gentamicin, dihydrostreptomycin, tetracycline, and chloramphenicol. D94B and D88A were resistant to ampicillin, dihydrostreptomycin, tetracycline, and chloramphenicol. In cattle farms 1 and 3, two pairs of isolates from cattle feces (D215A and D212B, D254A and D257A) indicated similar PFGE patterns (95% and 85%). The antimicrobial-resistance profiles of these two pairs of isolates were the same: D215A and D212B were resistant to ampicillin and tetracycline, while D254A and D257A were resistant to tetracycline.
Characterization of plasmids harboring tetracycline-resistance genes
We established 20 transconjugants derived from 32 isolates harboring tetA (62.5%) and 15 transconjugants derived from 28 isolates harboring tetB (53.6%; Table 3). Table 3 shows the characterization (tet genes, replicon type, plasmid length, and antimicrobial susceptibility profile) of transconjugants. Transferable tetracycline resistance genes harboring plasmids were observed in E. coli derived from flies and livestock feces in several places. Various types of replicon types were observed in plasmids. Some transconjugants exhibited resistance against multiple antimicrobials.
Bold type indicates tetA- or tetB-bearing plasmids.
ABPC, ampicillin; CP, chloramphenicol; DSM, dihydrostreptomycin; ERFX, enrofloxacin; GM, gentamicin; KM, kanamycin; TET, tetracycline; UT, undetermined type.
Strains D109B and D249A harbored similar plasmids, and strains D17A and D246A also harbored similar plasmids. These strains were isolated from different locations, from flies and livestock fecal samples, and exhibited different PFGE patterns.
The five pairs with similar PFGE patterns (D215A/D212B, D88A/D94B, D254A/D257A, D16A/D19B, and D18B/D205A) shared similar plasmids.
Discussion
This study suggests the role of flies not only in spreading clones of antimicrobial-resistant bacteria throughout a single farm but also in the widespread dissemination of plasmids carrying antimicrobial-resistance genes between farms. Although most flies do not travel a distance greater than 2 miles, certain individual flies can travel as far as 20 miles. 9 Moreover, flies have great potential for contaminating substrates, including food, with microbes during feeding and defecation. 29 These results suggest that antimicrobial-resistance genes can be transmitted to humans. Therefore, the hygiene of the animal ecosystem and prudent use of antimicrobials are required for control of the persistence and spread of antimicrobial-resistant bacteria and antimicrobial-resistance genes.
E. coli was isolated from the gut of only one stable fly (16.7%, 1/6), although E. coli was isolated from all other species of flies. Stable flies feed on blood; therefore, E. coli would be rarely isolated from the gut of a stable fly. 25 Compared with the other species of flies, the stable fly is not likely to play a critical role in disseminating antimicrobial-resistant strains of E. coli.
The resistance rates to most of the antimicrobials of isolates obtained from swine feces were much higher than those obtained from cattle feces. The same trends were observed in the national program for monitoring antimicrobial resistance (JVARM) in swine and cattle. 17 The highest antimicrobial-resistance rate of isolates obtained from swine feces was against tetracycline. A high frequency of tetracycline resistance results from the use of large amounts of tetracycline to maintain pigs. 16
The resistance rates for most antimicrobials of isolates obtained from flies in locations housing swine were much higher than those from cattle farms. The resistance rates of isolates obtained from flies were similar to those obtained from livestock feces from each location, suggesting that the resistance rate of E. coli derived from flies reflects that derived from livestock feces in the same locations.
The PFGE patterns of most of the tetracycline-resistant E. coli differed. Nevertheless, we observed similar PFGE patterns (>80% similarity) between the tetracycline-resistant isolates derived from houseflies and swine feces from the same swine farm (swine farm 1). Both of these isolates with similar PFGE patterns harbored tetB and exhibited the same antimicrobial susceptibility profile. Our previous study showed that flies carry several clones of E. coli that were also detected in cattle feces obtained from a cattle farm; these clones were also resistant to third-generation cephalosporins. 25 These results demonstrated that flies play a role in spreading clones of antimicrobial-resistant bacteria throughout a given farm.
Most of the tetracycline-resistant E. coli derived from livestock14,23 and flies harbored tetA or tetB. In line with our findings, Sawant et al. showed that tetracycline-resistant E. coli isolates were highly diverse across cattle farms. 22 Most tetracycline-resistance genes are encoded on plasmids. 20 Antimicrobial-resistance genes are horizontally transferred in the fly gut,1,18 and Binh et al. 3 suggested that swine feces serve as a reservoir of broad-host range plasmids with multiple antimicrobial-resistance genes. These results suggest that plasmids that harbor tetA, tetB, or both may be horizontally transmitted among the bacteria derived from flies and livestock feces in locations housing livestock.
In this study, similar plasmids carrying tetracycline-resistance genes (the same replicon type, antimicrobial-resistance gene, and the length of plasmid) were detected in strains isolated from different locations and generated different PFGE patterns. These results suggest that plasmids carrying tetracycline-resistance genes are widely disseminated. As vectors of antimicrobial resistance, several animals, such as wild birds and wild rodents, have been considered 19 . Our results indicate that flies may also serve as vectors of plasmids carrying antimicrobial-resistance genes and transmit them between farms and may further disseminate them over long distances to other regions, including human residences. Because flies contact habitats of humans more frequently compared with those of wild animals, flies may disseminate plasmids that carry antimicrobial-resistance genes between farms and human habitats.
Footnotes
Acknowledgments
We thank the staff of the Prefectural Livestock Hygiene Service Center (Northern Okinawa, Japan) for identifying the farms. This study was partially supported by a MEXT-supported program of the strategic research foundation at private universities and MEXT KAKENHI Grant No. 24590754.
Disclosure Statement
No competing financial interests exist.