
Abstract
The aim of this research was to statistically analyze the association between antimicrobial susceptibility/resistance to erythromycine, gentamicin, ciprofloxacin, and tetracycline and 11 virulence genes associated with adherence, invasion, and cytotoxicity in 528 isolates of Campylobacter coli and Campylobacter jejuni obtained from retail meat and fecal samples from food-producing animals and human patients. A high percentage of Campylobacter strains were resistant to antimicrobials, specifically ciprofloxacin and tetracycline. Moreover, we observed a wide distribution of virulence genes within the analyzed strains. C. jejuni strains were more susceptible to antimicrobials, and showed greater number of virulence genes than C. coli strains. Genes related to invasion capability, such as racR, ciaB, and pldA, were associated with antimicrobial-susceptible strains in both species. The genes cdtA and dnaJ, a citotoxin unit and an adherence-related gene, respectively, were associated with antimicrobial-resistant strains in both species. In conclusion, Campylobacter strains show a statistically significant association between antimicrobial susceptibility and the presence of virulence genes.
Introduction
C
Campylobacteriosis is characterized by a mild enteritis and is a self-limiting disease that does not usually require antimicrobial treatment. However, when it is associated with extra intestinal manifestations or is present in young children, pregnant women, or immunocompromised patients, it does require antimicrobial treatment. 38 For many years, macrolides and fluoroquinolones have been the first and second choices for antimicrobial treatment of Campylobacter enteritis. Since the late 1980s, however, the emergence of antimicrobial resistance (AMR) has complicated disease treatment. 26 In several countries, Campylobacter resistance to fluoroquinolones has limited its usefulness in the treatment of human infection. In addition, resistance to erythromycin is increasing, particularly in C. coli. Although the incidence of macrolide resistance in human strains is still relatively low, erythromycin should be regarded as the drug of choice in the treatment of Campylobacteriosis. 35 Furthermore, gentamicin, tetracycline, and doxycycline remain effective against Campylobacter and these agents should be considered only in serious Campylobacter infections.44,49 Campylobacter species have been shown to possess the genetic mechanisms for natural transformation and conjugation, indicating that if AMR genes were acquired, the trait would be rapidly transferred between animal and human strains. 1
Several putative virulence factors have been identified in Campylobacter species that contribute to motility, intestinal adhesion, colonization, toxin production, and invasion.4,17,27 Another important virulence factor is lipopolysaccharide, which has been related to Guillain–Barre syndrome, an autoimmune neurological disease associated with previous C. jejuni infection.28,47
Several studies have been conducted to determine the association between virulence genes and AMR in important bacterial pathogens, suggesting a link between resistance and the colonization or invasion potential of these bacteria.3,31,51,53 In addition, other studies have shown that humans infected with antimicrobial-resistant Campylobacter species have a longer duration of diarrhea than humans infected with antimicrobial-susceptible strains.19,33 Furthermore, acquired resistance generally confers substantial fitness costs on the bacteria in the absence of antimicrobial selection pressure. This situation could vary, depending on the antimicrobial class to which the bacterial strain is resistant and it could also be related to the molecular mechanism that generated its resistance. For example, in contrast to fluoroquinolone resistance, erythromycin resistance would have no observed fitness cost.18,29,52 However, a clear relationship between these two characteristics, AMR and virulence, is yet to be established.
Further investigation is required to determine whether there is indeed an association between AMR and the virulence of the pathogen. Thus, the aim of this study is to confirm such an association between antimicrobial-susceptible/resistance profiles in the presence or absence of virulence genes in several C. jejuni and Campylobacter coli isolates from retail meat, feces from food-producing animals, and in isolates from human patients.
Materials and Methods
Sampling procedures
Samples were collected between March 2012 and May 2013. All samples (chicken, turkey, swine, and bovine) were taken from healthy production animals randomly selected at a slaughterhouse located in the metropolitan region of Santiago, Chile, and its surrounding areas. Samples were also collected from supermarkets and wholesale stores by Food Control Authorities (SEREMI) in the metropolitan region of Chile. These samples were collected during the course of a routine inspection by the authorities and then transported to a laboratory in Cary Blair (Transystem®) medium (Difco®) and analyzed within 24 hours after collection.
For sample collection at animal-rearing farms and subsequent transportation to the laboratory, all procedures were consistent with those recommended by the World Organization for Animal Health (OIE) in the “Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2008”. 36 Fecal samples were collected from pigs, cattle, chicken, and turkeys; all animals were alive at the time of sampling. Animals were individually tested and all samples were collected at the same farms where these animals had been bred. Sampled chickens were between 42 and 45 days old, and sampled turkeys were between 112 and 126 days old. Sampled bovines were between 540 and 600 days old and sampled swines were between 160 and 180 days old. The samples were stored in Cary Blair (Transystem) medium.
Samples were collected randomly, taking only one sample per animal with a maximum of 30 animals per farm. The farms, therefore, are treated as independent epidemiological units. Samples from farms were tested only once during this study. In the case of chicken, turkeys, and pigs, samples were collected from each farm, or epidemiological unit, with a maximum of 30 animals of each type per farm/unit (i.e., 30 chicken, 30 cattle, etc. per farm). In this study, we considered each farm or epidemiological units as one isolated sample. For cattle, a sample was taken from each animal, and due to the lower prevalence of the pathogen in cattle, all isolates were used in the analysis.
Animal meat samples
Chicken, turkey, bovine, and pork meat samples were collected from meat that were refrigerated and ready for sale. Samples were transported to the laboratory at a temperature between 4°C and 8°C.
Human samples
Isolated human strains were collected from clinical cases at the Chilean Public Health Institute in agreement with the established collaboration between our laboratory and this Institute. The Public Health Institute isolated approximately 73 strains from clinical cases in the metropolitan region (greater Santiago) during the year 2012.
Microbiological isolation and identification
Strains obtained from humans
The clinical laboratories at various Chilean hospitals routinely send samples isolated from patients suspected of suffering Campylobacter enteritis to the Chilean Public Health Institute. In the bacteriology laboratory of the Institute of Public Health, selected fecal samples were cultured in two different mediums: Campylobacter selective agar (Preston) and blood-free Campylobacter selective agar mCCDA (Blood-Free Agar). In addition, an emulsion from the fecal sample was prepared in physiological saline and was treated using a filtration method (Milipore® 0.45 mm) with trypticase soy agar with 5% sheep's blood (Biomeriux®). Three plates were incubated at 42°C for 48 hours in an anaerobic jar with a microaerophilic environment generated by a sachet of CampyGen™ (Oxoid®).
Isolation of the presumptive colonies was carried out under a stereoscopic magnifying glass to observe the morphology and distinctive motility of samples. According to quantity and quality, cultures were transferred to trypticase soy agar with 5% sheep's blood and were then incubated for another 48 hours.
Fecal samples obtained from animals
This procedure followed the OIE recommendations for Campylobacter as given in the “Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2008”. 36
Summarizing, to isolate Campylobacter spp from animal fecal material, a blood-free selective medium mCCDA (OXOID) was used. Culture plates were put into an anaerobic jar containing an atmosphere generation system (CampyGen sachet, OXOID) and then incubated at 42°C for 48 hours. Presumptive colonies were confirmed by examination under a stereoscopic magnifying glass to observe morphology and distinctive motility. Colonies presenting characteristics compatible with the genus Campylobacter were cultured in trypticase soy agar with 5% sheep's blood (Biomeriux) and then incubated for another 48 hours, as previously described.
Samples obtained from meat
Initially we performed screening using VIDAS Campy (Biomeriux). Meat samples of 25 g were weighed and introduced into a sterile bag and then 100 ml of BOLTON broth (Oxoid) supplemented with antimicrobials was added. The bag was incubated in an anaerobic jar containing a Campygen (Oxoid) atmosphere generator sachet at 42°C for 48 hours. One to 2 ml of broth was transferred into a sterile tube, boiled to 95–100°C, and then 500 μl from the tube was placed in a VIDA campy equipment cartridge (Biomeriux) to be analyzed as positive or negative. Afterward, only positives samples were analyzed using the traditional methods described in the Food and Drugs Administration's (FDA) Bacteriological Analytical Manual (BAM). 21 Another 25 g of meat was weighed and a pre-enrichment in 250 ml of BOLTON broth was carried out, incubating the sample at 37°C for 48 hours in an anaerobic jar with a microaerophilic atmosphere. Samples were then incubated at 42°C for 48 hours in an anaerobic jar with a microaerophilic atmosphere. Then 100 ml of broth sample was cultured in a blood agar m-CCDA plate. The plate was incubated for 24 hours at 42°C in a microaerophylic atmosphere. Subsequently, presumptive colonies were confirmed by observation through a stereoscopic magnifying glass, as previously described.
Species identification
All colonies suspected to belong to the Campylobacter genus were identified to the species level through mass spectrometry using MALDI-TOF equipment (Bruker Daltonics, Inc., model Microflex LT®). Control strains C. jejuni ATCC 33560 and C. coli ATCC 43478 were used. Samples were prepared by mixing the samples with a matrix composed of alpha-Cyano-4-hydroxycinnamic acid, which has a strong optic absorption according to the wavelength of the selected laser, causing crystallization of the sample. A small amount of the bacterial strain was taken from a culture plate and placed into one of the 96 wells of the steel plate (target). Subsequently, the metal plate was introduced into the equipment and blasted with brief laser pulses. The spectrum generated was analyzed using the Byotiper 2.0 program, and compared to the Bruker 3.0 reference library, which is incorporated in the equipment used for detection.
Identification is expressed in scores. A score of ≥2.0 indicates identification at the species level, a score from ≥1.7 to 1.99 indicates reliable identification at the genus level, whereas a score under 1.7 does not allow for identification and is considered unreliable as the spectral acquisition was insufficient or protein peaks were not detected, and further analysis is required for the sample.
Determination of minimum inhibitory concentrations
Resistance was assessed using the agar plate dilution method. The minimum inhibitory concentration assay (MIC) was performed according to the method described by the Clinical and Laboratory Standards Institute M45-A2 and M100-S20. 6 C. jejuni ATCC 33560, Staphylococcus aureus ATCC, and Escherichia coli ATCC 35218 were used as quality control strains. The antimicrobials under evaluation were ciprofloxacin (Dr. Ehrenstorfer®), gentamicin (Sigma-Aldrich®), erythromycin (Sigma-Aldrich), and tetracycline (Sigma-Aldrich). Multidrug resistance was defined as the resistance to all three antimicrobial classes. One isolate per farm, per meat sample, or per patient was tested for antimicrobial susceptibility. All determinations were repeated two or more times. MIC90 values, as well as percentages of resistance, were calculated.
Interpretation of the results of the Campylobacter isolates was performed using the resistant breakpoints published by CLSI (20106) and EUCAST, (2013 14 ). Cutoff points used to classify a strain as resistant were ciprofloxacin ≥4 μg/ml, erythromycin ≥32 μg/ml, gentamicin ≥16 μg/ml, and tetracycline ≥16 μg/ml. These same points were used for isolated strains from human patients, food, and animals.
Detection of virulence genes through PCR
Virulence was determined in all isolates using PCR analysis for the presence of the following genes: flaA, dnaJ, cadF, virB11, rarR, ciaB, pdlA, cdtA, cdtB, cdtC, and wlaN, as previously described by Talukder et al. 2008. 45 In summary, bacterial DNA extraction was carried out from trypticase soy agar with 5% sheep's blood 48-hour cultures. Three colonies from each strain were boiled at 95°C for 10 minutes and then centrifuged at 10,000 g for 5 minutes. Table 1 details the primer sequences, length of the PCR products (pb), and specific alignment temperature (Tm; °C). The amplification program consisted of an initial denaturalization at 95°C for 5 minutes, 30 cycles at 95°C for 30 seconds, specific Tm for each primer for 30 seconds, and 72°C for 30 seconds, followed by a final extension at 72°C for 5 minutes. PCRs were carried out with 1 μl of DNA using RBC taq DNA Polymerase (RBC Bioscience®) and 5 μM of each primer. PCR products were then electrophoresed on a 2% (p/v) agarose gel containing GelRed™ 10,000 g (Biotium®). Two PCR positive strains were randomly chosen for sequencing and confirmed using the GenBank database of the program NCBI BLAST. Accession numbers were flaA: KF846052; cadF: KM092516; racR: CP006729; dnaJ: KJ081742; virB11: CP000550; ciaB: AF114831; wlaN: CP006729; pldA: GQ491060; cdtA: KJ15268; cdtB: KJ875956, and cdtC: JX658757.
Pulse-field gel electrophoresis
This assay followed the PulseNet protocol by Ribot et al. 2001. 40 In brief, bacterial suspensions adjusted to an optical density of 1.3 at 610 nm were embedded in 1% SeaKem Gold agarose plugs. DNA was digested with SmaI (Roche®) at a concentration of 40 U per endonuclease sample, the plugs were incubated at a temperature of 23–25°C for 2 hours. The macrorestriction fragments were separated by electrophoresis using CHEF DRIII CHILER (Bio-Rad®) equipment in 1% Pulsed Field Certified Agarose gels (Ultrapure DNA grade agarose) at 6V/cm and 14°C for 18 hours. We used a Salmonella serotype Braenderup H9812 strain, previously digested by SmaI as a base pair market and run control. Images were analyzed with the BioNumerics GelCompar II 6.0 software (Applied Maths). The similarity between fingerprints was determined using Dice's correlation coefficient with a 1% tolerance between band positions. Cluster analysis and generation of dendrograms were performed using UPGMA, and the discriminatory power was calculated using the Simpson's diversity index, as reported in a previous study. 22
Statistical analysis
AMR was considered as a binary dependent variable (0 = nonresistant; 1 = resistant). The association between the resistance profile of each antimicrobial and the presence or absence of virulence genes was assessed using multiple logistic regression models. Every model included the presence/absence of each gene as a binary explanatory variable (0 = present; 1 = absent) and associations were considered significant when p ≤ 0.05.
To determine significant differences in the number of virulence-related genes between species (C. jejuni vs. C. coli, factorial analysis of variance (ANOVA) was carried out using the number of genes as a dependent variable and the Campylobacter species as a factor. Means were considered significant when p ≤ 0.05. All of the analyses described earlier were performed using the Infostat® software (www.infostat.com.ar/).
Results
Campylobacter isolates
A total of 528 spatially and temporally related Campylobacter isolates were analyzed. Out of the total, 318 samples were identified as C. jejuni and 210 as C. coli through mass spectrometry. Data are summarized in Table 2, in which the number of analyzed samples, the isolate percentage for each source, and the number of strains isolated for each species are presented.
Antimicrobial susceptibility
The results for antimicrobial susceptibility in relation to species (C. jejuni or C. coli) and sample origin (human, animal feces, or meat), as determined by the agar plate dilution method, are shown in Tables 3 and 4. Overall, C. coli strains were significantly more resistant to erythromycin, ciprofloxacin, and tetracycline than C. jejuni strains. Also, it was observed that Campylobacter isolates from animal feces were significantly more resistant to antimicrobials than to isolates from human patients or meat samples. Resistance to one or two groups of antimicrobials was more commonly found than MDR. The most frequent MDR profile was ciprofloxacin/tetracyclin/erythromycin, which was identified in 26% of strains. Two strains of C. coli obtained from human samples presented a simultaneous resistance to ciprofloxacin and erythromycin.
The breakpoints: ciprofloxacin ≥4 mg/L, erythromycin ≥32 mg/L, gentamicin ≥16 mg/L, tetracycline ≥16 mg/L.
The breakpoints: ciprofloxacin ≥4 mg/L, erythromycin ≥32 mg/L, gentamicin ≥16 mg/L, tetracycline ≥16 mg/L.
Virulence gene detection
Campylobacter strains analyzed carried an average of six virulence genes. In general, virulence gene prevalence in strains was high, and the most frequently identified genes were as follows: cadF, flaA, cdtA, cdtB, and cdtC, whereas the least prevalent gene was wlaN. Specifically in the case of human isolates, the most prevalent genes were cadF (93%), cdtC, and cdtB (85%). For isolates obtained from animal feces, in the case of chicken, the most prevalent genes were cadF (98%), cdtA (98%), and cdtB (98%); in the case of turkeys, they were cdtA (76%), cadF (72%), and cdtB (69%). For isolates obtained from bovine fecal samples, the most prevalent genes were cdtC (71%), cadF (66%), and cdtA (63%), whereas in swine fecal samples the most prevalent genes were cdtA (90%), cadF (75%), and flaA (72%). Regarding meat samples, in chicken isolates the most prevalent genes were flaA and cdtA (63%), followed by cdctC (60%). Finally, the most prevalent genes in isolates from turkey meat were cdtA (63%), cdtC (63%), and cadF (58%) (Fig. 1).
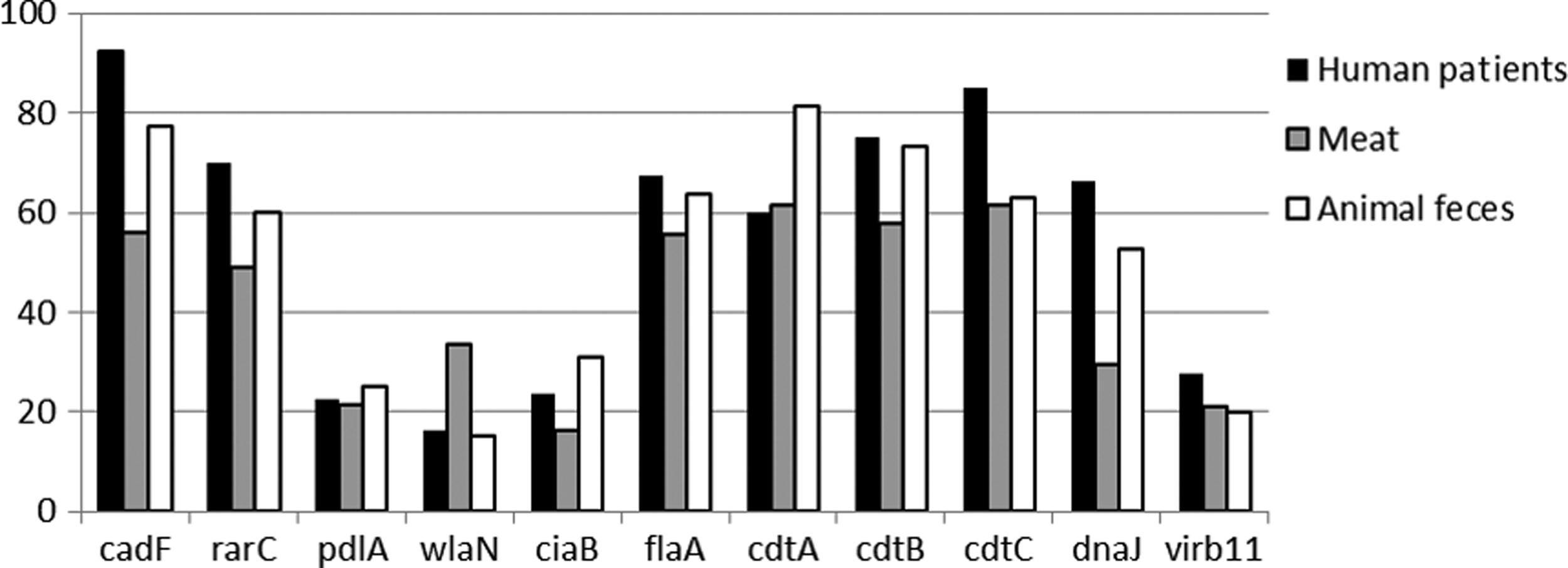
Percentage of Campylobacter jejuni and Campylobacter coli, isolated from human patients (n = 73), meat (n = 113), and food-producing animals feces (n = 342), which resulted positive to each of the 11 virulence genes under analysis.
When analyzing the results by Campylobacter species, it was observed that C. jejuni strains presented a higher percentage of virulence genes than C. coli, with a p < 0.05. The most prevalent gene for C. jejuni was pldA (82%) and the least prevalent gene was cdtA (54%), whereas in C. coli, the opposite was observed, as the most prevalent gene was cdtA (45%) and the least common gene was pldA (17.8%), as shown in Fig. 2.
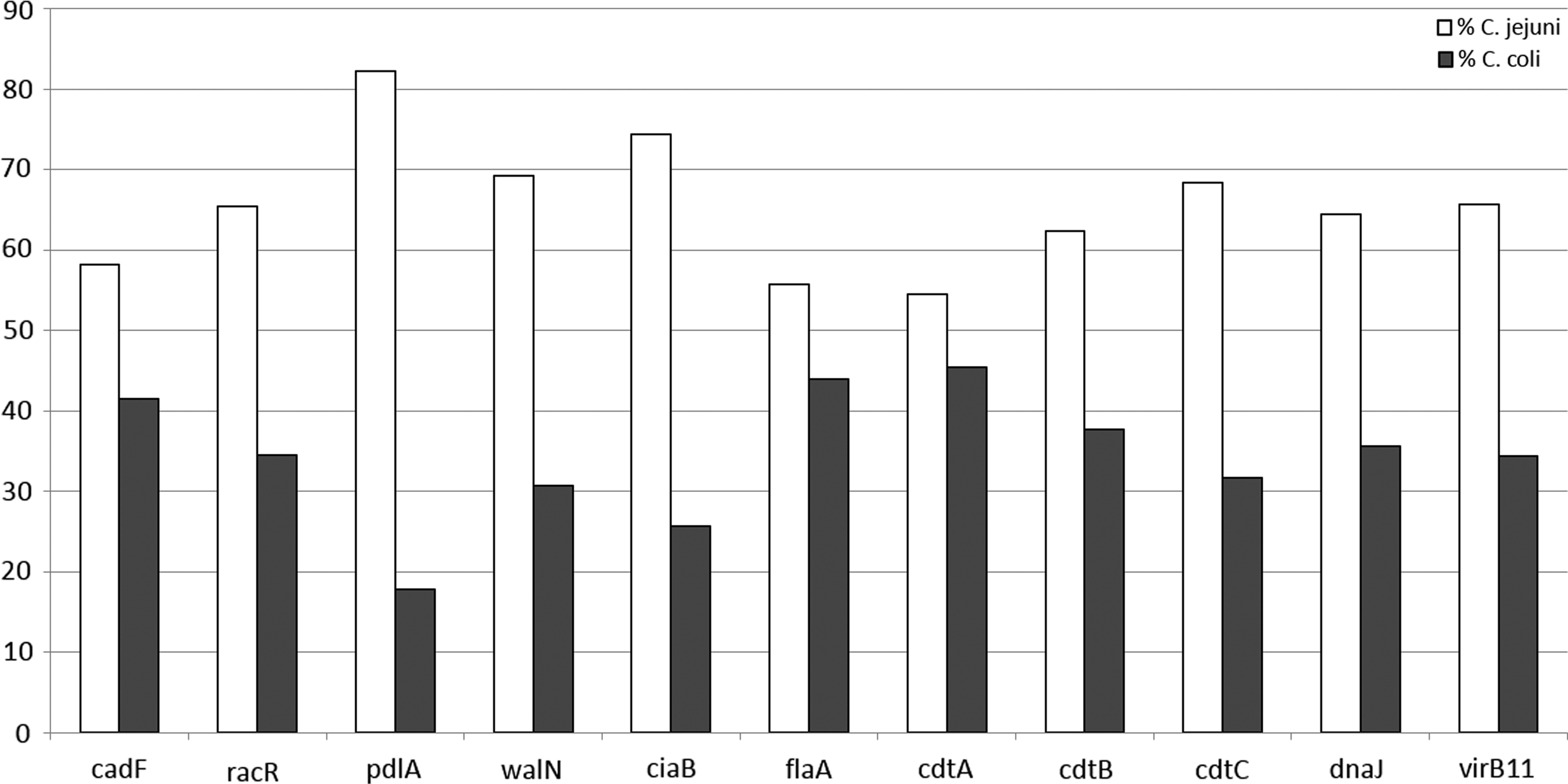
Percentage of C. jejuni strains (light bar) and C. coli strains (dark bar), which resulted positive to each of the 11 virulence genes under analysis. C. jejuni strains showed a higher number of virulence genes than C. coli with a p-value = 0.001.
Regarding the relationship between the presence of the virulence genes and the sensitivity of the strains to antimicrobials, susceptible isolates showed a greater number of virulence genes than resistant isolates. In general, for isolates from human patients and isolates obtained from meat and animal feces, gentamicin-, erythromycin-, and tetracycline-susceptible strains showed increased resistance genes as compared to ciprofloxacin-susceptible strains (Fig. 3). Ciprofloxacin- and tetracyclin-resistant strains showed a greater number of virulence genes than erythromycin- and gentamicin-resistant strains (Fig. 4).

Percentage of virulence genes present in C. jejuni and C. coli strains that were susceptible to ciprofloxacin, tetracycline, erythromycin, and gentamicin, isolated from human patients
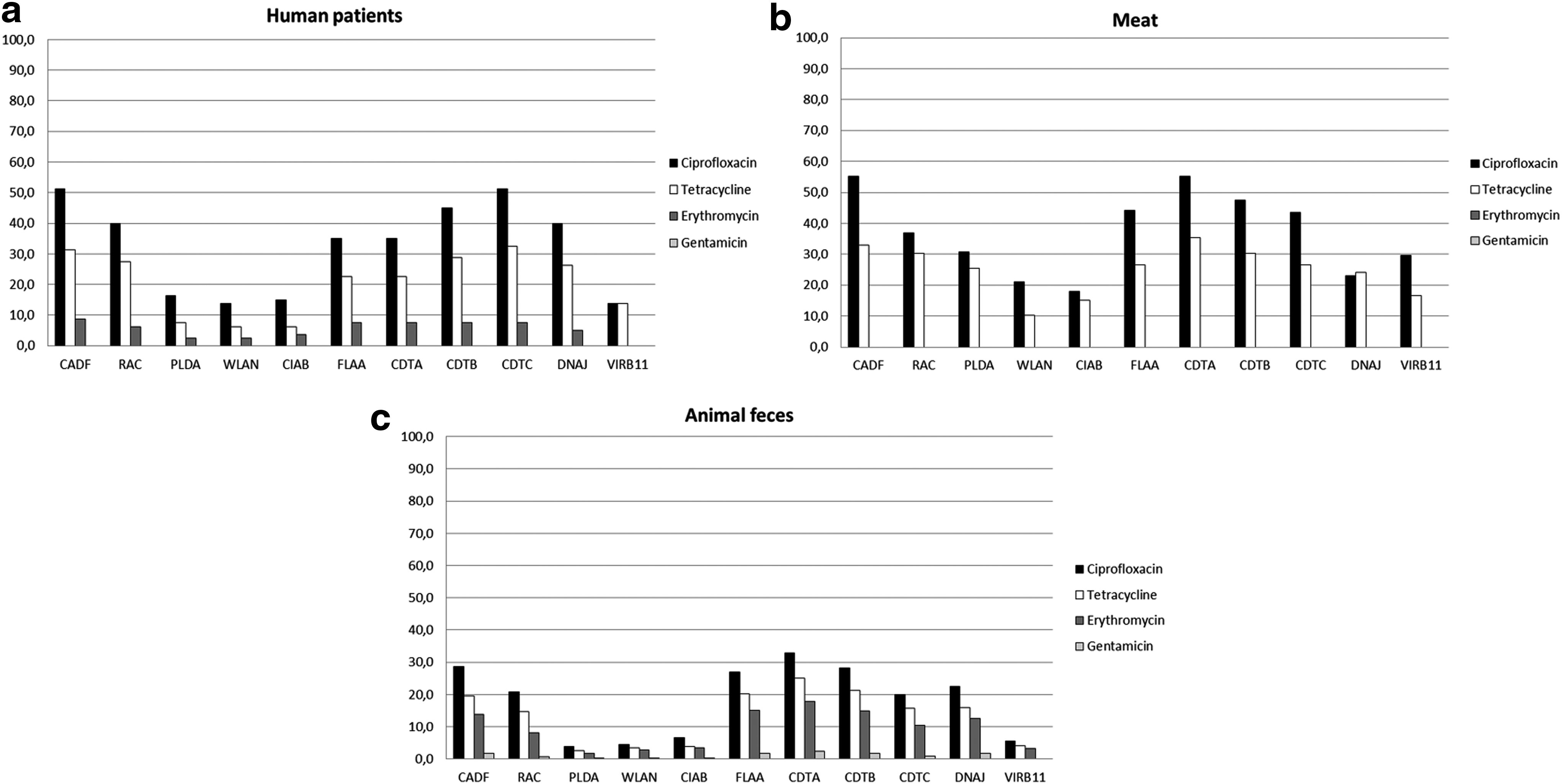
Percentage of virulence genes present in C. jejuni and C. coli strains that were resistant to ciprofloxacin, tetracycline, erythromycin, and gentamicin, isolated from human patients
Association between virulence genes and antimicrobial susceptibility
Multiple logistic regression models were performed for every isolate using antimicrobial susceptibility profiles as a dependent variable and the presence/absence of the 11 virulence genes as an independent variable. For ciprofloxacin, significant differences were observed (p < 0.05 and an odds ratio [OR] <1) for the virulence genes racR, ciaB, and cdtC. These results indicate that susceptible strains are associated with the presence of virulence genes. Also, an association was observed between tetracycline sensitivity and the genes ciaB and cdtC, as well as between erythromycin and the genes racR, pldA, and cdtC. In contrast, the genes cdtA and dnaJ were associated with strains resistant to tetracycline or erythromycin (p < 0.05 and an OR >1) (Tables 5–7). When analyzing the association between resistance/susceptible to antimicrobials and presence/absence of virulence genes, it was observed that C. jejuni presented an association between the gene dnaJ and ciprofloxacin and tetracycline resistance. Tetracycline-susceptible C. jejuni strains were associated with the gene ciaB and erythromycin with the gene racR. Regarding C. coli, tetracycline resistance was associated with the presence of the gene dnaJ and erythromycin susceptibility with the gene racR. The presence of the gene cdtC was associated with strains that were found to be susceptible to all antimicrobials tested.
The analysis includes all isolated Campylobacter strains. Antimicrobial susceptibility is defined as an OR <1 and p ≤ 0.05, and AMR is defined as an OR >1 and p ≤ 0.05.
indicates an association between antimicrobial susceptibility and the specific virulence gene whereas
indicates an association between AMR and the specific virulence gene.
The analysis includes all isolated C. jejuni strains. Antimicrobial susceptibility is defined as an OR <1 and p ≤ 0.05, and AMR is defined as an OR >1 and p ≤ 0.05.
indicates an association between antimicrobial susceptibility and the specific virulence gene, whereas
indicates an association between AMR and the specific virulence gene.
The analysis includes all isolated C. coli strains. Antimicrobial susceptibility is defined as an OR <1 and p ≤ 0.05, and AMR is defined as an OR value >1 and p ≤ 0.05.
indicates an association between antimicrobial susceptibility and the specific virulence gene, whereas
indicates an association between AMR and the specific virulence gene.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) analysis was performed on 90 strains isolated from chicken fecal samples, of which 65 were identified as the species C. jejuni and 25 as the species C. coli. The isolates were collected from different farms, or epidemiologic units, which are all located in the same region of the country: the greater metropolitan region of Santiago. Strains were defined as having the same pulse type if they shared 100% identity. For C. jejuni strains, nine clusters with 100% similarity were found, each of which was composed of two strains. In addition, 47 pulse types were made up of only one strain.
The Simpson's diversity index for C. jejuni strains was 0.9 (Fig. 5). Of the 25 C. coli isolates, there were 25 pulse types represented by one strain. The Simpson's diversity index for C. coli strains was 0.9 (Fig. 6).
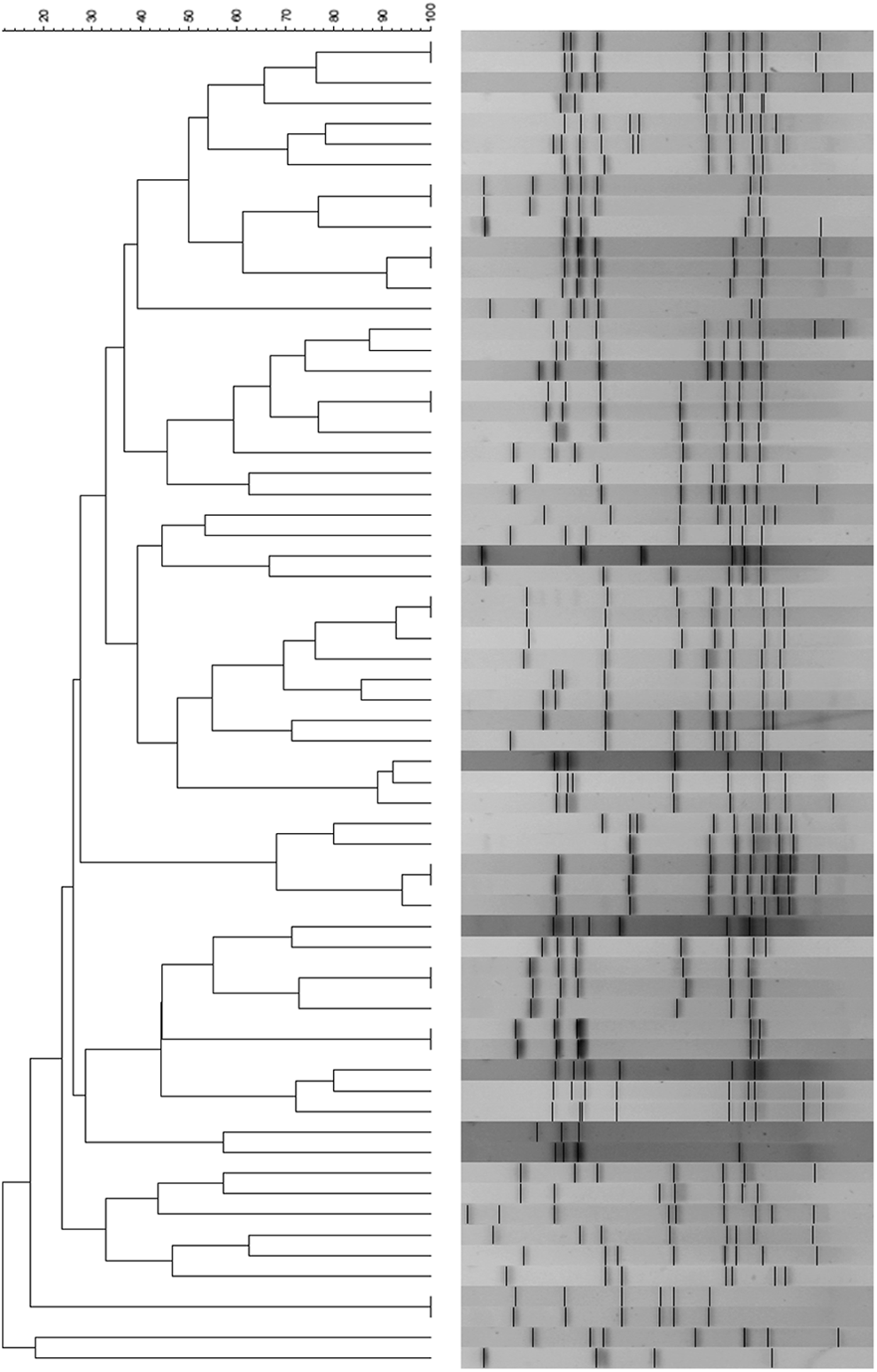
Pulsed-field gel electrophoresis (PFGE) analysis of C. jejuni isolated from chicken.
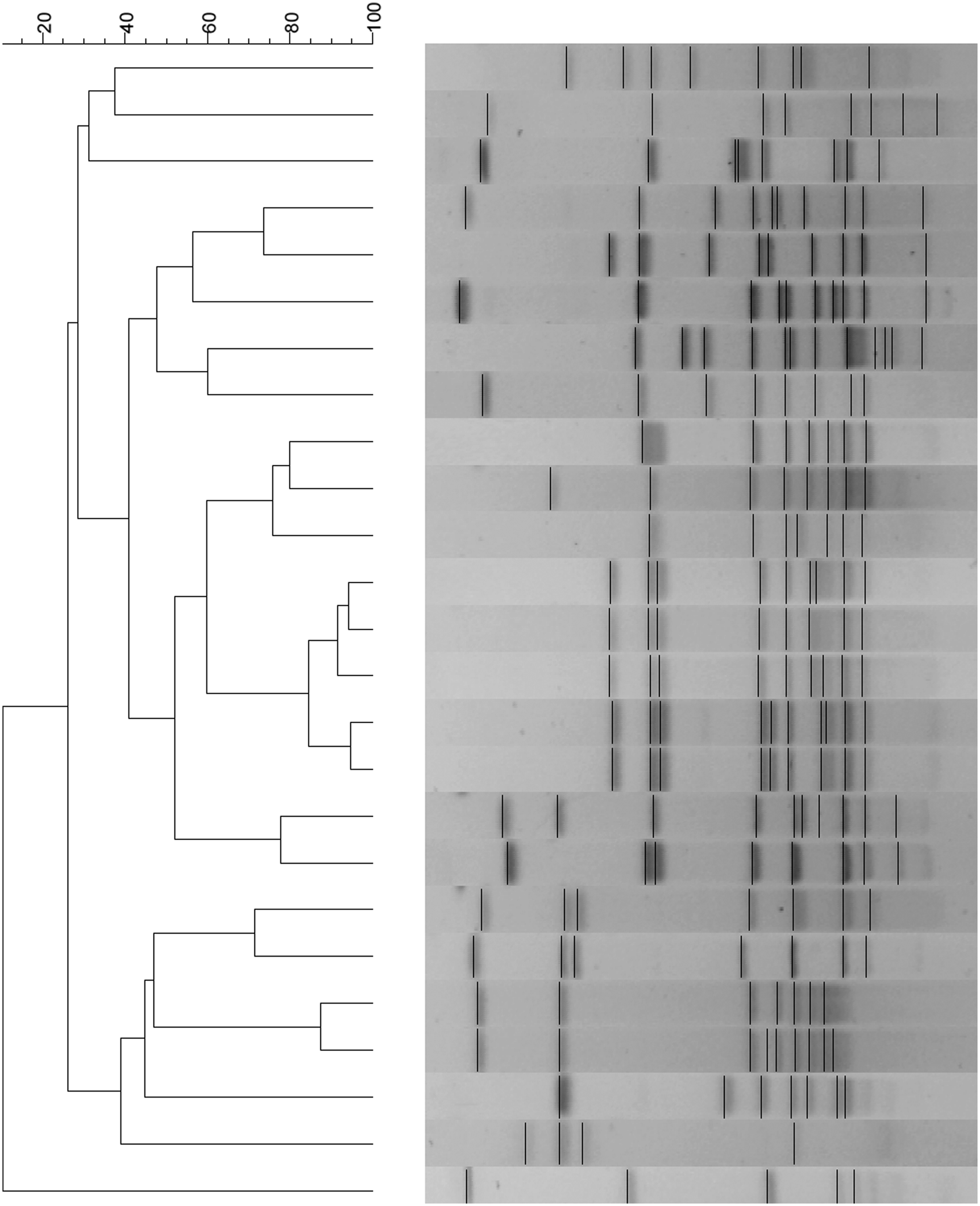
PFGE analysis of C. coli isolated from chicken.
Discussion
This study involved isolation of Campylobacter strains from different origins. A high occurrence of Campylobacter was observed in the intestines of several food-producing animals, with the exception of the bovine samples. In retail meat, most of the strains were isolated from chicken and turkey, confirming that chicken and its by-products constitute a risk of Campylobacter spp. transmission for the population. The isolates obtained from chickens were genetically diverse, with 55 clusters identified in the case of C. jejuni and 21 clusters in the case of C. coli strains (Figs. 5 and 6). The PFGE technique reached a discrimination level of 0.9 in both cases.
The results of species identification using MALDI TOF showed that 60.2% of the identified bacteria were C. jejuni and 39.8% were C. coli. However, in patients diagnosed with campylobacteriosis, only 9.6% of isolates were characterized as C. coli. In this study, there was a significant difference (p = 0.01) in the number of virulence genes present in C. coli and C. jejuni, with the latter presenting a higher number of virulence genes (Fig. 2).
In terms of the analyzed virulence genes, the most common were cadF and flaA, which have been associated with the bacterial adherence capacity to epithelial cells, which is a key mechanism in causing disease. 17 Thakur et al. 2010 also found these genes to be prevalent in Campylobacter strains. 46 Other prevalent genes included cdt, which is related to the production of cytotoxin, and cdtA, cdtB, and cdtC, which cause diarrhea by interfering with the division and differentiation of cells in the intestinal crypt. All three subunits are required for full toxin activity. 37 The least common gene was wlaN, which is involved in the biosynthesis of lipooligosaccharide that may show ganglioside-mimicking structures and thus may be related to developing Guillain–Barré syndrome after C. jejuni infection. 46
The number of virulence genes present in pathogenic strains does allow us to predict whether disease will develop in humans. It is also important to emphasize that the exact pathogenesis of Campylobacter infection in humans has not yet been fully elucidated. 7 Due to differences in the number and combination of virulence genes in different Campylobacter populations by origin (human, animal, and meat), our results indicate that the degree of virulence between these sources differs. Other authors have also identified such variability in the virulence genes found in C. jejuni strains. 8
Another factor that must be considered for pathogenic strains that cause food-borne diseases is the fact that they may be resistant to antimicrobial treatments used in human health. The occurrence of bacterial AMR is a major issue in public health, as many infections are becoming increasingly difficult to treat due to this issue. AMR is particularly acute in the case of food-borne pathogens, due to the fact that these pathogens often cause outbreaks that affect many people in disparate geographical areas, which may result in the transmission of resistance determinants to a considerable portion of the population.
In this article, we analyzed antimicrobial sensitivity in strains from Campylobacter isolates. Our results show that resistance varies from moderate to high, and that C. coli strains in general show a higher percentage of AMR than C. jejuni, a finding that has been described previously by other authors.11,13,25,41,42
Erythromycin resistance levels were relatively low; however, a greater number of macrolide-resistant strains were observed in C. coli fecal isolates from pigs. Environmental contamination with erythromycin-resistant C. coli originating from the intestines of pigs, which could in turn contaminate water, soil, and wild animals carrying resistance genes, represents a hazard for ecosystem conservation, human and animal health, and even the economy of some countries. Ciprofloxacin and tetracycline resistance was high in both species of Campylobacter, regardless of the origin. This is an important issue with regard to the use of quinolones, which has previously been described by other authors.8,9,12,43,46,50 The cause of such high resistance to quinolones could be related to the broad use of enrofloxacin in veterinary medicine.16,20 The WHO 48 has indicated that this antimicrobial class should be classified as a critical group that must be considered when developing resistance monitoring programs and guidelines for appropriate antimicrobial administration.
Our data showed differences in resistance levels and in the distribution of MICs between C. jejuni and C. coli. Similarly, other authors have described that C. coli isolates from animals are more prone than C. jejuni isolates to harbor resistance to antimicrobials, including macrolides and fluoroquinolones. 11 D'lima et al. 200711 found higher levels of multiresistance in C. coli isolated from turkeys than from other animals, such as chicken, pigs, and cattle. In this study, strains isolated from turkeys showed high levels of resistance to tetracyclin, ciprofloxacin, and erythromycin. Thus, it is possible that animals involved in food-borne diseases, especially turkeys and pigs, are subjected to inadequate doses of antimicrobials, or that these doses are being administered for a long period of time. Thus, the inadequate use of antimicrobials in food-producing animals, especially in the case of C. coli strains, could lead to a resistant phenotype with more ease than C. jejuni strains. One possible explanation for such a difference is that C. coli strains are more likely to acquire horizontal resistance determinants and/or that target genes mutate faster in the presence of inadequate antimicrobial doses.
On looking at the relationship between virulence gene prevalence, antimicrobial susceptibility, and strain origin, our results show that there is a greater number of virulence genes as well as an increased diversity of virulence genes in antimicrobial-susceptible strains than in resistant strains (Figs. 3 and 4). We also observed that C. jejuni strains are less resistant to antimicrobials than C. coli strains. This difference is due to the fact that C. jejuni strains had lower MIC values than antimicrobials studied. In addition, these strains had a higher number of resistant genes than C. coli species. The reason for such a difference between these two Campylobacter species, especially regarding a higher AMR in C. coli strains, has not yet been clarified.
It is important to highlight that although there are fewer cases of gastroenteritis in humans caused by C. coli, this species is capable of producing the disease. Thus, cases of C. coli gastroenteritis may pose additional risk due to the fact that this species shows high MIC against antimicrobials typically used for treatment. Some studies have shown an association between AMR, especially to quinolones and erythromycin, and the virulence of Campylobacter strains in humans, 26 whereas other reports do not support or confirm this relationship. 15 There are studies that indicate decreased virulence in erythromycin-resistant strains, 30 whereas others show an increase in in vitro invasion of resistant strains as compared to susceptible strains. 53 The bacterial infection process requires complete adhesion, invasion, and subsequent cytotoxin production. This complexity could explain the differences that have been reported.
Based on these data, one aim of this research was to demonstrate the existence of an association between antimicrobial susceptibility/resistance and virulence genes and, thereby, an association between the presence of each virulence genes and susceptibility and/or resistance to antimicrobials found in isolates of different origins. Statistical analysis shows a clear correlation between the presence of virulence genes and antimicrobial susceptibility, which could indicate that for antimicrobial-resistant C. jejuni and C. coli, there is an associated cost to virulence. More studies are needed to determine whether this correlation is related to the presence of a specific resistance determinant that could alter the presence of one or more virulence genes in Campylobacter strains, and whether this decrease in the presence of virulence genes translates into a decrease in adhesion or invasion of cells in vitro.
Analyzing our results for particular genes showed that there is an association between susceptibility to all antimicrobials studied here and the cdtC gene. In addition, ciprofloxacin-susceptible strains were associated with the presence of racR and ciaB, tetracycline-susceptible strains with the presence of ciaB, and erythromycin-susceptible strains with the presence of racR and pldA. Antimicrobial-resistant strains were associated with the presence of the cdtA and dnaJ genes. In general, a higher presence of virulence genes was associated with susceptible strains in C. jejuni isolates as well as in C. coli strains.
Regarding the function of virulence genes, the gene racR is a component of the racR and RACS (reduced ability to colonize) regulation system and plays a role in the ability of these bacteria to colonize the intestinal tracts of chicken. 4 The ciaB gene is important for the invasion of epithelial cells and the colonization of the intestines. The pldA gene is related to cell invasion and encodes a protein involved in the synthesis of an outer membrane phospholipase. 54 Virulence genes associated with AMR included cdtC, a cytotoxin unit, and dnaJ, which is involved in adherence to epithelial cells. As indicated previously, the presence of the three cdt genes (cluster) is required for the cytotoxin to be functional, which is why the presence of a single cdt gene would not have any effect on the strains' virulence. In contrast, the dnaJ gene could be very important in Campylobacter pathogenicity.
Some of the virulence genes associated with antimicrobial-susceptible strains in C. jejuni and C. coli are involved in bacterial invasion capability. It could be deduced then that susceptible strains have a higher invasion potential than resistant ones, which contrasts with previous descriptions by Zeitouni et al. 2013. 53 Nevertheless, this author showed this association using mutant strains invading Caco-2 cells in vivo, which does not allow for a comparison of results. The link between resistance and virulence relies on several factors, such as bacterial population, bacterial diversity, strain, and origin, among other factors, all of which should be considered to validate the observations.
In summary, our work shows high levels of AMR to ciprofloxacin and tetracycline in C. jejuni and C. coli, which could indicate a low clinical usefulness. In addition, we observed a disperse distribution of the 11 virulence genes in the analyzed strains. Strikingly, C. coli showed higher levels of AMR and a lower number of virulence genes than C. jejuni. This could be related to the fact that C. jejuni causes the majority of campylobacteriosis cases in humans in both developing and developed countries. Finally, we showed a statistically significant association between antimicrobial susceptibility and the presence of virulence genes, specifically those genes related to invasion capacity. Further research is needed on this subject. Likewise, further investigation should be carried out on this important emergent pathogen, aiming to establish better control and defense measures, to decrease the risk of contamination with this pathogen and to mitigate the emergence and transmission to the population and environment of resistant or multiresistant strains.
Footnotes
Acknowledgment
The authors acknowledge the funding from FONDECYT Project number 11110200.
Dislosure Statement
No competing financial interests exist.