
Abstract
The aim of this study was to evaluate the antimicrobial susceptibility profile of 85 Staphylococcus epidermidis and 84 Staphylococcus haemolyticus strains isolated from blood cultures to oxacillin, vancomycin, tigecycline, linezolid, daptomycin, and quinupristin/dalfopristin over a period of 12 years. S. epidermidis and S. haemolyticus isolated from blood cultures of inpatients, attended at a teaching hospital, were analyzed for the presence of the mecA gene and by SCCmec typing. The minimum inhibitory concentration (MIC) values of tigecycline, linezolid, daptomycin, quinupristin/dalfopristin, and vancomycin were determined. Isolates exhibiting vancomycin MICs of ≥2 μg/ml were typed by pulsed-field gel electrophoresis (PFGE). The rate of mecA positivity was 92.9% and 100% in S. epidermidis and S. haemolyticus, respectively. The most frequent SCCmec types were type III (53.2%) in S. epidermidis and type I (32.1%) in S. haemolyticus. All isolates were susceptible to linezolid and daptomycin, but 7.1% of S. haemolyticus and 2.3% of S. epidermidis isolates were resistant to tigecycline, and 1.2% each of S. haemolyticus and S. epidermidis were resistant and intermediately resistant to quinupristin/dalfopristin, respectively. S. epidermidis exhibited higher vancomycin MICs (40% with MIC of ≥2 μg/ml). Clonal typing of strains with vancomycin MIC of ≥2 μg/ml revealed the presence of different PFGE types of S. epidermidis and S. haemolyticus over a period of up to 4 years (2002–2004, 2005–2008, 2006–2009, 2010–2011). Despite the observation of a high prevalence of mecA, the clinical strains were fully susceptible to vancomycin and to the new drugs linezolid, daptomycin, tigecycline, and quinupristin/dalfopristin. The PFGE types with vancomycin MIC of ≥2 μg/ml exhibited a great diversity of SCCmec cassettes, demonstrating that S. epidermidis and S. haemolyticus may easily acquire these resistance-conferring genetic elements.
Introduction
S
Methicillin-resistant staphylococci are a public health concern. In the 1990s, the global prevalence of methicillin resistance among CoNS ranged from 70% to 80%, 11 whereas surveillance studies have shown an oxacillin resistance rate of 75–85% among CoNS in recent years.14,27,51 The empirical use of the glycopeptide vancomycin for the treatment of sepsis and other infections caused by CoNS is supported by the high frequency of strains carrying the mecA gene. This gene encodes an additional penicillin-binding protein (PBP2a), which confers resistance to beta-lactam antibiotics, including methicillin/oxacillin. The gene is carried by a mobile genetic element, SCCmec (staphylococcal cassette chromosome mec), which serves as a vehicle for the exchange of antimicrobial resistance genes among staphylococci. The indiscriminate use of vancomycin is a matter of concern and CoNS were the first organisms in which glycopeptide resistance was recognized (vancomycin minimum inhibitory concentration [MIC] of ≥8 μg/ml). 5 The vanA and vanB genes, which encode resistance to vancomycin and were first described in enterococci 2 and subsequently identified in Staphylococcus aureus (www.cdc.gov/HAI/settings/lab/vrsa_lab_search_containment.html), have recently been reported in CoNS, such as S. epidermidis and Staphylococcus saprophyticus.18,35
MICs of vancomycin have increased progressively in recent years, with levels higher than 2 μg/ml being observed in a large number of S. epidermidis and S. haemolyticus isolates. 34 Although they are within the range of susceptibility, these MICs may indicate a decrease in the in vivo efficacy of vancomycin. 37
Considering the steadily increasing rates of resistance to oxacillin and the reduction in susceptibility to vancomycin, new drugs are needed for the treatment of staphylococcal infections. Potential alternatives include linezolid, tigecycline, daptomycin, and quinupristin/dalfopristin. Although not approved for the treatment of bloodstream-associated infections due to high mortality rates in clinical trials, linezolid and tigecycline show high efficacy against CoNS. Most CoNS strains isolated from bacteremias are highly susceptible to daptomycin, and quinupristin/dalfopristin exhibits an excellent bactericidal activity and a prolonged postantibiotic effect against gram-positive cocci. 25
In view of the emergence of antimicrobial resistance in CoNS and the limitation of treatment options, the objective of the present study was to evaluate oxacillin resistance based on mecA positivity and MICs of vancomycin, linezolid, tigecycline, daptomycin, and quinupristin/dalfopristin in S. epidermidis and S. haemolyticus isolated from blood cultures over a period of 12 years.
Materials and Methods
Isolates
The strains were isolated between 2000 and 2011 from blood cultures of inpatients admitted to the University Hospital of the Botucatu Medical School (Hospital das Clínicas, Faculdade de Medicina de Botucatu—HC-FMB), UNESP (Universidade Estadual Paulista), Botucatu, São Paulo, Brazil. The University Hospital of FMB is a 418-bed tertiary care hospital and referral center that offers high-complexity services and receives patients from other secondary services in the region. Only one isolate per patient was included in the study. The strains were isolated on blood agar. Bacterial colonies were stained by the Gram method and submitted to catalase and coagulase tests, as described by Koneman et al. 22
Species identification
Glucose oxidation and fermentation tests were performed as described by Koneman et al. 22 for differentiation from Micrococcus. S. epidermidis and S. haemolyticus were identified by the simplified method proposed by Cunha et al. 10
DNA extraction
The Illustra kit (GE Healthcare, Little Chalfont, United Kingdom) was used for DNA extraction after the digestion of staphylococcal cells with lysozyme (10 mg/ml) and proteinase K (20 mg/ml).
Genotypic identification of S. epidermidis and S. haemolyticus
Species identification was genetically confirmed by PCR amplification of the 16S–23S internal transcribed spacer region, as described by Couto et al. 9 The following reference strains were used as controls: S. epidermidis ATCC® 12228™, S. epidermidis ATCC® 35983™, and S. haemolyticus ATCC® 29970™.
Detection of the mecA gene
Real-time PCR for detection of the mecA gene was performed using the StepOnePlus® System (Life Technologies, Carlsbad, CA) according to the parameters of Vandecasteele et al. 46 Reference strains were included in all reactions as positive (S. aureus ATCC 33591) and negative (S. aureus ATCC 25923) controls.
SCCmec typing
SCCmec typing was performed by multiplex PCR according to the protocol of Machado et al. 24 The following international reference strains were used as controls of SCCmec types: S. aureus COL (SCCmec type I), S. aureus N315 (type II), S. aureus 85/2082 (type III), and S. aureus JCSC3624 (type IV). For the nontypable isolates, multiplex PCRs 1 and 2 were used according to Kondo et al. 21 The protocol of Machado et al. 24 consists of a single multiplex reaction that permits the identification of types I–IV, while the two multiplex reactions of the protocol of Kondo et al. 21 identify SCCmec types I–VI. The method of Kondo et al. 21 with some frequency yields inconclusive results for SCCmec of CoNS due to the absence or duplicity of one of the ccr/mec complexes, but can detect cassette types other than types I–IV. These problems in SCCmec typing are not encountered with the technique of Machado et al. 24 ; however, this method only detects types I–IV.
Determination of MICs of the antimicrobials by the E-test
The MICs for vancomycin, linezolid, tigecycline, daptomycin, and quinupristin/dalfopristin were determined by the E-test® (BioMérieux, Marcy l'Étoile, France) using the Mueller–Hinton agar medium, an inoculum with a final concentration adjusted to a 0.5 McFarland turbidity standard, and incubation at 35°C for 24 hr according to the manufacturer's instructions. According to the guidelines of the CLSI and of the European Committee on Antimicrobial Susceptibility Testing (EUCAST), testing for daptomycin susceptibility in staphylococci should be performed by broth dilution in cation-adjusted Mueller–Hinton broth (CAMHB, supplemented with 50 mg/L calcium). In addition, CLSI guidelines 8 clearly indicate that susceptibility testing by agar dilution has not been validated for daptomycin. In the present study, susceptibility to daptomycin was determined by the E-test according to the manufacturer's recommendations, which do not define the addition of calcium. CLSI guidelines 8 were used to classify the susceptibility of the isolates to all drugs, except for tigecycline. Susceptibility to tigecycline was evaluated according to the breakpoints defined by EUCAST 12 for staphylococci. The E-test was repeated for strains that were nonsusceptible to the drugs tested.
Determination of vancomycin MICs by the broth microdilution method
For broth microdilution, log2 serial dilutions of vancomycin were prepared in CAMHB (Oxoid, Basingstoke, United Kingdom) in sterile microtiter plates according to CLSI recommendations. 8 The plates were incubated in an oven at 35°C, and MICs were determined after 24 hr (CLSI 8 ) and 48 hr of incubation for comparison. Enterococcus faecalis ATCC 29212, S. aureus ATCC 29213, and E. faecalis ATCC 51299 were used as controls. The cutoff values proposed by the CLSI 8 were used for classification of the isolates (susceptibility: ≤4 μg/ml; intermediate resistance: 8–16 μg/ml; resistance: ≥32 μg/ml).
Comparison of the broth microdilution method and the E-test
Agreement between 24-h broth microdilution and E-test was defined as a variation of up to ±1 log2 dilution (±1 two-fold dilution) between results. Differences of ≥2 log2 dilutions were defined as discordant results.
Detection of the vanA and vanB genes
All isolates with vancomycin MICs of ≥2 μg/ml were analyzed regarding the presence of the vanA and vanB genes. The parameters described by Clark et al. 7 were used for PCR detection of the vanA and vanB genes.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) typing was performed according to the modified protocol of McDougal et al. 29 A higher volume of the inoculum was used (250 μl), and 2 μl lysozyme (10 mg/ml) was added for complete lysis. A dendrogram was generated with the UPGMA method using the Dice coefficient calculated using the Bionumerics 7.1 software, with 1.0% optimization and 1.25% band position tolerance. A similarity index of 80% was used to define PFGE types.
Statistical analysis
For evaluation of the association of drug MICs with oxacillin resistance (mecA) and SCCmec, the MICs determined by the E-test were divided into the following intervals (mg/L): vancomycin: 0.125–0.38, 0.5–1.0, and 1.5–3.0; linezolid: 0.047–0.19, 0.25–0.75, and 1.0–1.5; quinupristin/dalfopristin: 0.025–0.094, 0.125–0.75, and 0.75–4.0; tigecycline: 0.016–0.094, 0.0125–0.5, and 0.75–2.0; and daptomycin: 0.016–0.032, 0.047–0.094, and 0.125–0.25. The χ 2 test was used to evaluate associations between variables, adopting a level of significance of 0.05.
Results
A total of 169 isolates, corresponding to 85 S. epidermidis and 84 S. haemolyticus strains, were analyzed. Table 1 shows the characteristics of the patients, such as age and gender, and the year of isolation of the strains. The mecA gene was detected in 84 (100%) of the S. haemolyticus isolates and in 79 (92.9%) of the S. epidermidis isolates. These strains were submitted to SCCmec typing. Table 2 shows the distribution of SCCmec types among the S. epidermidis and S. haemolyticus isolates. The isolates that were nontypable by the protocol of Machado et al. 24 were submitted to multiplex PCRs 1 and 2, as described by Kondo et al. 21 : one S. epidermidis could be classified as type V. All other strains carried nontypable SCCmec with distinct patterns, including the lack of a mec and/or ccr amplicon in 20 (12.2%) strains and double ccr or mec allotypes in 2 (1.2%) strains.
Surgical centers: Cardiac, Gastric, and Thoracic Surgery and Neurosurgery. Internal medicine: Clinical Medicine, Cardiology, Clinical Gastroenterology, Hematology, and Nephrology.
Central ICU includes Coronary ICU.
F, female; ICU, intensive care unit; M, male; NI, no information.
The MICs of linezolid, tigecycline, daptomycin, and quinupristin/dalfopristin are shown in Table 3. All isolates were classified as susceptible to linezolid and daptomycin. Six (7.1%) S. haemolyticus and two (2.3%) S. epidermidis isolates were classified as resistant to tigecycline. All these isolates were positive for the mecA gene; four S. haemolyticus isolates carried SCCmec type II, one S. epidermidis and one S. haemolyticus isolate carried type III, one S. epidermidis isolate carried type I, and one S. haemolyticus isolate could not be typed. One (1.2%) of the S. haemolyticus isolates was classified as resistant to quinupristin/dalfopristin, and one (1.2%) S. epidermidis isolate exhibited intermediate resistance. Both isolates carried the mecA gene and SCCmec type IV.
Breakpoints according to CLSI: linezolid (S ≤ 4; R ≥ 8 μg/ml); quinupristin/dalfopristin (S ≤ 1; I = 2; R ≥ 4 μg/ml); daptomycin (S ≤ 1 μg/ml).
Tigecycline breakpoints according to EUCAST (S ≤ 0.5 μg/ml). 39
Intermediate resistance.
DAP, daptomycin; CLSI, Clinical & Laboratory Standards Institute; EUCAST, European Committee on Antimicrobial Susceptibility Testing; LIN, linezolid; MIC50, minimum inhibitory concentration required to inhibit the growth of 50% of organisms; MIC90, minimum inhibitory concentration required to inhibit the growth of 90% of organisms; QN/DF, quinupristin/dalfopristin; TIG, tigecycline.
The E-test showed vancomycin MICs for CoNS ranging from 0.125 to 3 μg/ml. In contrast, the broth microdilution method provided MICs of 0.125–2 μg/ml after 24 hr of incubation and of 0.125–4 μg/ml after 48 hr. Table 4 and Fig. 1 show the MICs of vancomycin obtained by the broth microdilution method and E-test after 24 hr of incubation, as well as the comparison of the results obtained by the two methods for the two species. According to the E-test and broth microdilution, 25 (14.8%) and 57 (33.7%) isolates exhibited MICs of ≥2 μg/ml, respectively. Eleven (13%) and 34 (40%) S. epidermidis isolates exhibited MICs of ≥2 μg/ml according to the E-test and broth microdilution, respectively. Fourteen (16.7%) and 23 (27.4%) of the S. haemolyticus isolates exhibited MICs of ≥2 μg/ml according to the E-test and broth microdilution, respectively. Considering a variation of up to ±1 log2 concentration, agreement between the broth microdilution method and the E-test was observed among 76 (89.4%) S. epidermidis isolates, among 73 (86.9%) S. haemolyticus isolates, and among 149 (88.2%) of all isolates. None of the isolates with a vancomycin MIC of ≥2 μg/ml was positive for the vanA and vanB genes.
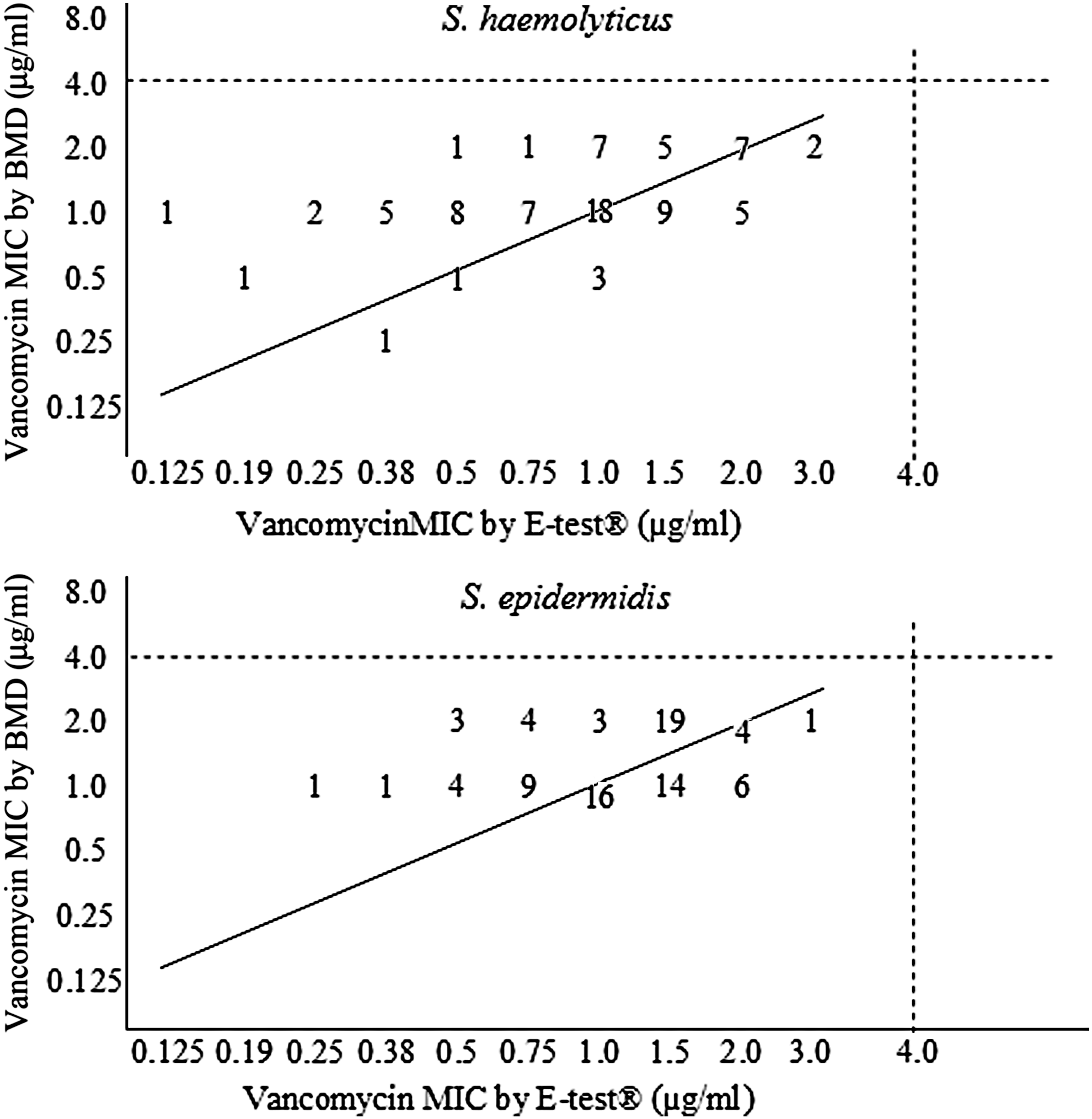
Dispersion graph showing the correlation between vancomycin minimum inhibitory concentrations (MICs) obtained by the broth microdilution method (BMD) and E-test® for Staphylococcus haemolyticus and Staphylococcus epidermidis (upper and lower panels, respectively). Dotted lines indicate the susceptibility cutoff values of coagulase-negative staphylococci (CLSI 35 ). The solid line indicates total agreement between the results of the two methods.
BMD, broth microdilution.
The percentage of isolates with vancomycin MICs of 1–1.5 μg/ml by the E-test declined over the period of 12 years (from 73.9% in 2000–2002 to 42.4% in 2009–2011), while the percentage of isolates with MIC of ≥2 μg/ml increased over the same periods from 13% to 23%. Figure 2 illustrates the separate analysis for the two species.

Evolution of vancomycin MICs (μg/ml) determined by the E-test from 2000 to 2011. The results are reported as number of isolates. SE, S. epidermidis; SH, S. haemolyticus.
Statistical analysis of the MIC intervals, presence of the mecA gene, and SCCmec type revealed an association between higher quinupristin/dalfopristin MICs and the presence of the mecA gene (P = 0.01), and between lower tigecycline MICs and SCCmec types I and IV and higher MICs and types II and III (P = 0.01). Moreover, an association was observed between lower tigecycline MICs and absence of the mecA gene. All mecA-negative isolates exhibited tigecycline MICs in the lowest range (P = 0.008).
The clonal profile of strains with vancomycin MICs of ≥2 μg/ml was analyzed by PFGE. Several PFGE types that included more than one isolate were found, eight S. epidermidis PFGE types (Fig. 3) and six S. haemolyticus PFGE types (Fig. 4). The year of isolation is shown in Figs. 3 and 4. Major S. epidermidis PFGE types were present from 2001 to 2004 and from 2008 to 2009, and major S. haemolyticus PFGE types comprised strains isolated from 2006 to 2007 and from 2010 to 2011 (Figs. 3 and 4). Most of the S. epidermidis isolates with vancomycin MICs of ≥2 μg/ml, which formed clusters, also carried SCCmec types III and I, and the most prevalent PFGE types carried the same types. A greater diversity of SCCmec types was observed among S. haemolyticus PFGE types.
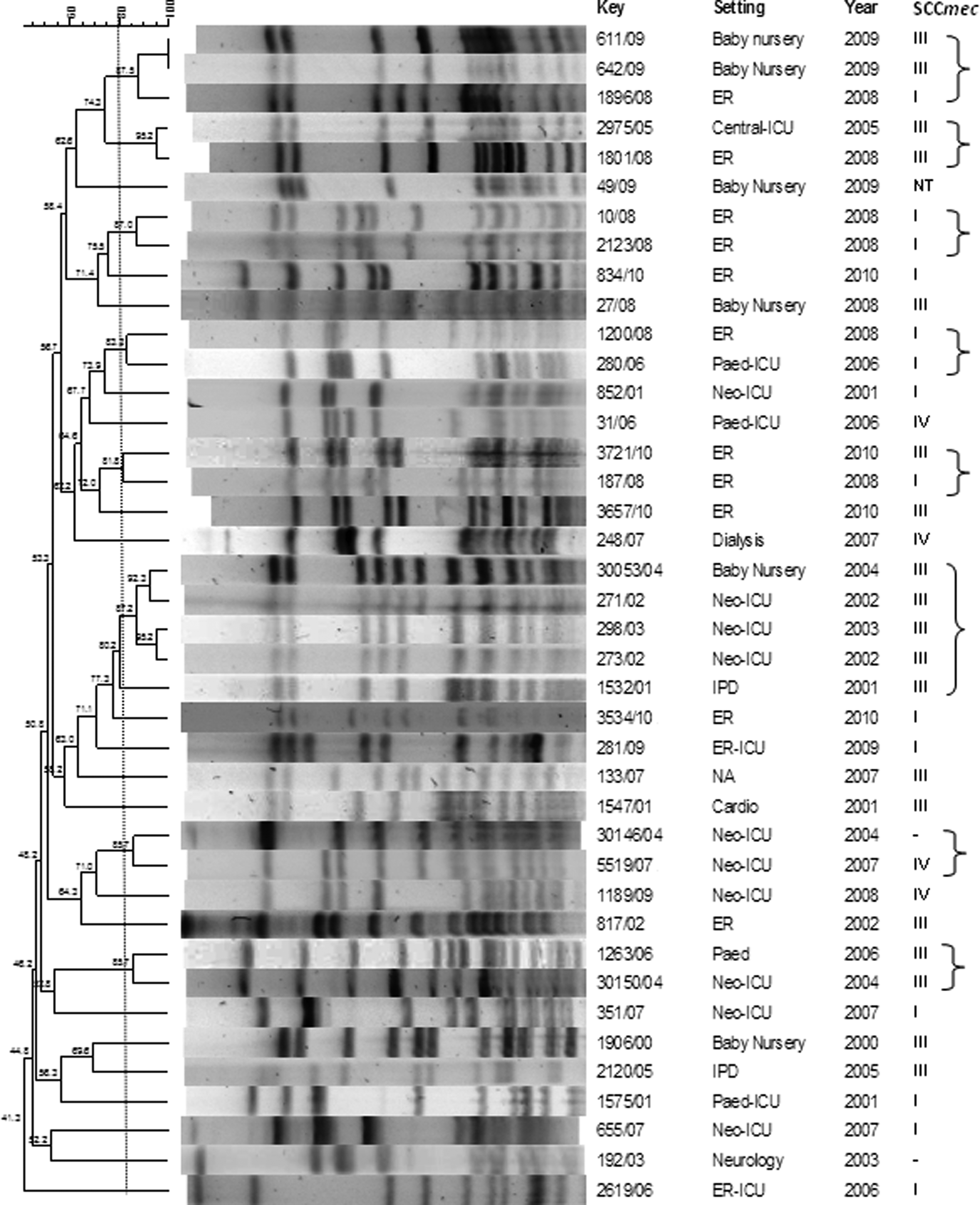
Pulsed-field gel electrophoresis (PFGE) of S. epidermidis isolates with vancomycin MICs of ≥ 2 μg/ml and their SCCmec type. The lines and brackets separate strains of the same PFGE type (similarity coefficient ≥ 80%).
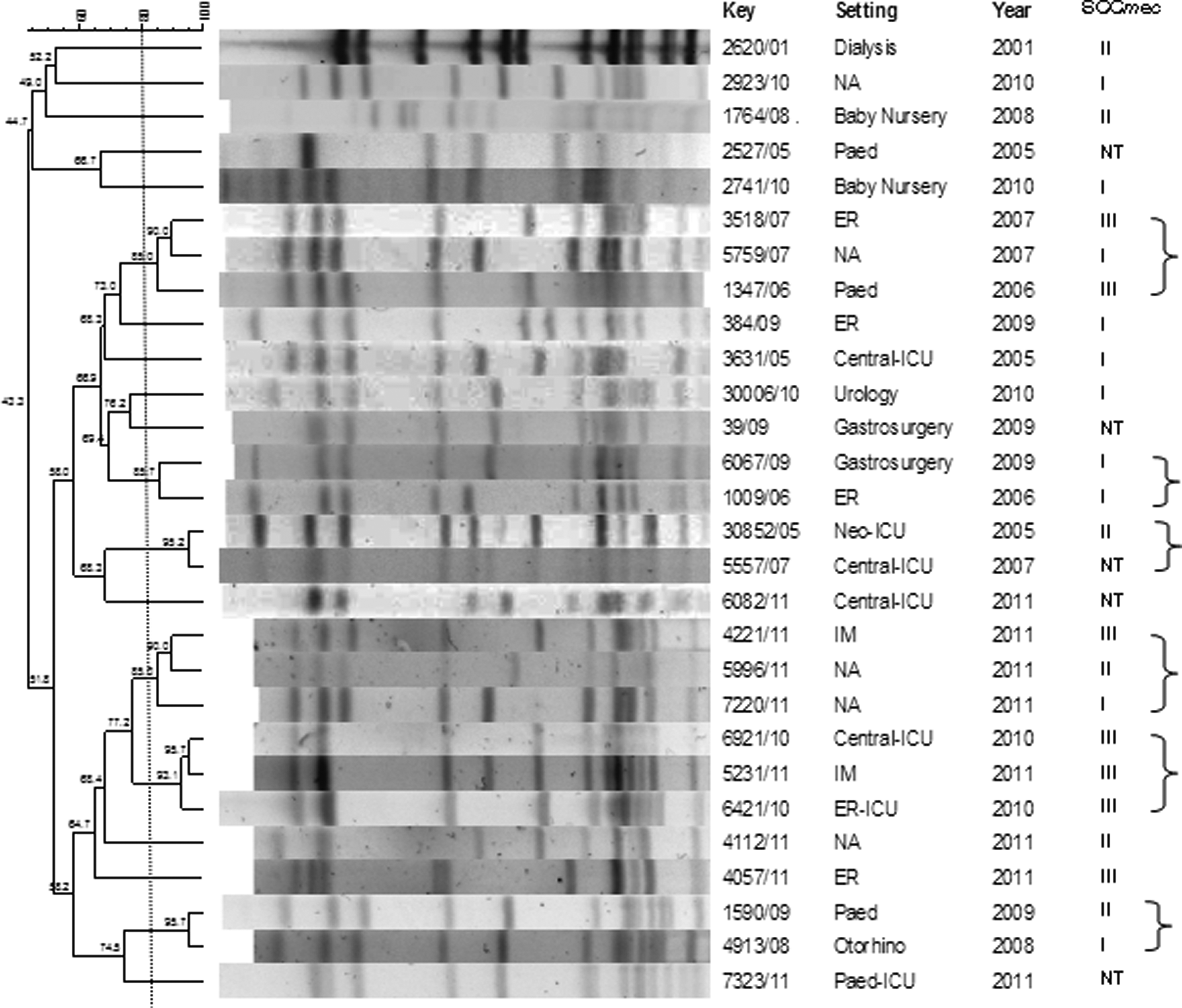
PFGE of S. haemolyticus isolates with vancomycin MICs of ≥ 2 μg/ml and their SCCmec type. The lines and brackets separate strains of the same PFGE type (similarity coefficient ≥ 80%).
Discussion
Resistance of CoNS associated with human infections to antimicrobial drugs is becoming increasingly common, especially resistance to beta-lactam antibiotics represented by oxacillin/methicillin. In recent decades, a worldwide increase in the number of CoNS with reduced susceptibility to vancomycin has also been reported, 17 and novel antimicrobial drugs have been developed in an attempt to solve the problem of resistance in CoNS.
The high rates of mecA positivity among CoNS are a matter of concern since transfer of the element that contains this resistance gene, SCCmec, occurs across staphylococcal species. The present results revealed that 100% of S. haemolyticus hospital isolates and 92.9% of S. epidermidis isolates were resistant to oxacillin. A high rate of oxacillin resistance has also been reported in previous studies, with 80% of resistant hospital isolates in Brazil.38,45 The differences between the present results and those reported in previous studies may be explained by the higher sensitivity of real-time PCR for detecting the mecA gene compared to conventional PCR, and by the fact that this study included only S. epidermidis and S. haemolyticus isolates and no other CoNS species that commonly exhibit much lower oxacillin resistance rates. 1 The mecA positivity rate of 100% observed for S. haemolyticus agrees with the description of a multidrug resistance phenotype in this species.
SCCmec typing using two different protocols permitted the identification of types I, III, IV, and V in S. epidermidis, and of types I, II, III, and IV in S. haemolyticus. SCCmec type II was detected only in S. haemolyticus and was not found in any of the S. epidermidis isolates. The frequency of SCCmec type III, the most complex element that is associated with high levels of oxacillin or cefoxitin resistance and resistance to nonbeta-lactam antibiotics, was expected in the two species since the strains studied were hospital isolates.
The S. epidermidis isolates showed a relatively low frequency of SCCmec type IV compared to previous studies. 30 Although it is small and has a lower metabolic cost for transfer, SCCmec type IV is generally associated with lower rates of antimicrobial resistance and does not carry any other resistance genes. Furthermore, the isolates with this cassette exhibit lower oxacillin MICs than hospital types I, II, and III.4,19,36
The percentage of strains with untypable SCCmec was higher for S. haemolyticus (22.6%), a finding that might be explained by the presence of novel classes of SCCmec not identified by the protocols used in this study. The genomic plasticity suggested for CoNS species, such as S. epidermidis and S. haemolyticus, 43 may facilitate the insertion of different genetic elements in orfX and thus contribute to the frequent generation of new SCC elements. S. haemolyticus is considered a reservoir of SCCmec type V, 40 but the protocol by Kondo et al. 21 could not identify this type in that species, suggesting the presence of novel classes or mutations in S. haemolyticus SCCmec. Although S. epidermidis has been considered a reservoir of SCCmec type IV, 40 our results show a great diversity of SCCmec in this species, relevant findings for the epidemiology of MRSE (methicillin-resistant S. epidermidis).
PFGE combined with SCCmec typing showed a great diversity among PFGE types with reduced susceptibility to vancomycin, particularly among S. haemolyticus isolates. These findings suggest that these microorganisms easily lose and acquire SCCmec elements because of a high rate of recombination events and genetic acquisition. 30 According to Miragaia et al., 30 strains of the same PFGE type or subtype, clonal complex, or sequence type (multilocus sequence typing) can carry different SCCmec types due to the frequent acquisition of SCCmec and the high rate of insertion/excision of SCCmec in the chromosome of CoNS. 31
Recent surveillance studies have reported resistance of CoNS to linezolid, with rates ranging from 1% to 2%. 13 Some studies have shown that the frequent use of linezolid is associated with resistance, 39 mainly involving cases of clonal dissemination. In contrast, the isolates studied here were fully susceptible to linezolid, with MICs ranging from 0.047 to 1.5 μg/ml. Linezolid susceptibility of 100% in CoNS has also been reported by other authors, 17 with maximum MICs of 4 μg/ml. The MIC50 and MIC90 values of linezolid were slightly higher for S. haemolyticus. In contrast, Stuart et al. 42 observed the lowest values for S. haemolyticus (1 μg/ml) when compared to other CoNS (2 μg/ml). Nevertheless, linezolid continues to be one of the most effective antibiotics against CoNS and the low rate of resistance to this drug is a good determinant for its use against multidrug-resistant gram-positive bacteria.
Tigecycline is considered effective in the treatment of infections caused by gram-positive and gram-negative bacteria. In the present study, tigecycline MICs ranged from <0.016 to 2.0 μg/ml in S. epidermidis and S. haemolyticus. Other studies reported slightly lower values of 0.125–0.5 μg/ml. 42 While the CLSI has not yet determined tigecycline breakpoints for staphylococci, EUCAST 12 (for staphylococci) and the FDA (only for S. aureus; Tygacil package insert [June 2005]; Wyeth Pharmaceuticals, Inc., Philadelphia, PA) recommend a susceptibility breakpoint of ≤0.5 μg/ml. The FDA also states that, although MICs of 0.5 μg/ml or less are found in 90% of isolates, the clinical significance and efficacy of this antimicrobial agent are unknown. 44
The tetracycline resistance genes (tet) that mediate the production of efflux pump proteins and ribosomal protection may lead to an increase in MIC values but do not confer resistance to tigecycline because these efflux pumps are not able to export glycylcyclines from the cell. 16 Some studies have traced the tigecycline resistance of S. aureus to increased expression of the mepRAB gene cluster (multidrug export protein), particularly the mepA efflux pump. 28 Since tigecycline is not the drug of choice for bloodstream infections caused by S. epidermidis and S. haemolyticus, the resistance of the eight isolates of this study may have been mediated by an antimicrobial therapy-independent mechanism. Studies are needed to identify the mechanisms of tigecycline resistance in CoNS.
In the present study, the greatest variations in tigecycline MICs were also observed for S. haemolyticus (<0.016–2.0 μg/ml). One (1.2%) S. haemolyticus isolate exhibited an MIC of 2 μg/ml. This isolate also carried the mecA gene and had a vancomycin MIC of 2 μg/ml. Several studies have reported S. haemolyticus strains with MICs higher than 1 μg/ml and suggested a possible correlation between high tigecycline MICs and the presence of the mecA gene,42,17 although previous studies failed to establish such correlation. 15
S. haemolyticus has been described to have a great potential of developing multidrug resistance. We detected one (1.2%) isolate that was resistant to quinupristin/dalfopristin. Ribosomal target modification (erm genes), export by efflux pumps (vgaA and vgaB genes), and inactivation by enzymes (vatA, vatB, and vatC genes) have already been associated with resistance to quinupristin/dalfopristin. 33 The resistant strain also carried the mecA gene and SCCmec type IV, features that may permit its dissemination in the hospital environment. These data highlight the importance of S. haemolyticus as a multidrug-resistant pathogen that is difficult to treat with conventional antibiotics.
One S. epidermidis isolate was not susceptible to quinupristin/dalfopristin (MIC = 1.5 μg/ml). A previous study showed that mechanisms of resistance to both quinupristin and dalfopristin are essential for the development of resistance in staphylococci. 48 The loss of resistance to dalfopristin or quinupristin or the acquisition of a single mechanism only leads to intermediate susceptibility, 48 which may be the case for the nonsusceptible S. epidermidis isolate found in this study. Nevertheless, quinupristin/dalfopristin appears to have good efficacy, and the potential of developing resistance to the drug is low as demonstrated by its prolonged postantibiotic effect. 47
All S. epidermidis and S. haemolyticus clinical strains proved to be susceptible to daptomycin, which was the drug showing the lowest MICs that were below the susceptibility breakpoints defined by the CLSI. 8 Since the drug has been approved only recently by the FDA and is still used rarely in Brazil, daptomycin resistance is very uncommon among CoNS isolates. Therefore, despite occasional cases of resistant CoNS, daptomycin shows high in vitro efficacy against these microorganisms. 6
The present results confirm the efficacy of vancomycin against CoNS, with MICs below the susceptibility breakpoint (≤4 μg/ml). However, treatment with vancomycin might be less effective in infections caused by isolates with MICs higher than 2 μg/ml. 32 Since vancomycin MICs of 2 μg/ml were observed in 33.7% of the isolates by the gold standard method, caution is necessary when indicating this antimicrobial drug for the treatment of these microorganisms. Higher vancomycin MICs were observed in S. epidermidis (40% of the isolates with MICs of 2 μg/ml), in agreement with some studies.38,23
The difference in the percentage of isolates with vancomycin MICs of ≥2 μg/ml between the E-test and broth microdilution method (14.8% vs. 33.7%) might be due to the different scales used, since fewer dilutions were used in the latter method. In this respect, the MIC50 and MIC90 values of vancomycin confirm that the E-test subtly overestimated MICs in relation to the gold standard, that is, broth microdilution. In the present study, the agreement between the two methods was 88.2%, slightly higher for S. epidermidis than S. haemolyticus.
The higher vancomycin MIC determined by the broth microdilution method after 48 hr might be related to the presence of subpopulations with increased vancomycin MICs, which are only detectable after a longer incubation period, since the MICs for the controls S. aureus ATCC 29213 and E. faecalis ATCC 29212 and ATCC 51299 continued to be constant during the same period. The longer incubation period (48 hr) may also permit the expression of vancomycin-heteroresistant subpopulations. 38
Clonal analysis of isolates with vancomycin MICs of ≥2 μg/ml revealed the presence of some S. epidermidis PFGE types present from 2001 to 2004 and from 2008 to 2009, and of S. haemolyticus PFGE types present from 2006 to 2007 and from 2010 to 2011. These data suggest the dissemination of some PFGE types in hospital settings that, although susceptible to vancomycin, have higher vancomycin MICs.
Furthermore, a subtle increase in vancomycin MICs of the S. epidermidis and S. haemolyticus isolates was observed over the period of 12 years. These observations characterize an increase in vancomycin MICs in the hospital environment and the dissemination of less susceptible PFGE types.
Footnotes
Acknowledgments
We thank Prof. Dr. Antonio Carlos Pignatari, Laboratório Especial de Microbiologia Clínica (UNIFESP), for providing the Enterococcus faecalis ATCC 51299 and E. faecalis ATCC 29212 strains. This work was supported by the state funding agency Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; Grant 2011/15396-1).
Disclosure Statement
No competing financial interests exist.