
Abstract
An increase of Enterobacteriaceae isolates with reduced susceptibility to cefepime (FEP) and amoxicillin/clavulanate (AMC) has been observed in our area. The aim of this study was to characterize this antibiotic resistance phenotype and its molecular epidemiology. A total of 33 Enterobacteriaceae strains were studied. blaOXA-1 genes and their genetic environment were analyzed by polymerase chain reaction (PCR) and sequencing. Plasmids were transferred by conjugation and/or transformation and classified using PCR-based inc/rep typing and IncF subtyping. Escherichia coli isolates were typed by phylogroup, pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing. Outer membrane proteins were studied by sodium dodecylsulfate–polyacrylamide gel electrophoresis and expression of blaOXA-1 genes by reverse transcription-PCR. FEP minimum inhibitory concentration yielded values of 1–16 mg/L. Twenty-nine (87.9%) isolates produced OXA-1, of which 24 (82.7%) were located in class 1 integron, and 9 (27.3%) produced TEM-1. Among the 24 E. coli OXA-1-producers, PFGE revealed two main clusters: one belonged to C-ST88 and the other to B23-ST131. Thirteen plasmids containing blaOXA-1 were transferred, nine belonged to IncF replicon (4 F2:A1:B-, 2 F1:A1:B1, 1 F1:A2:B-, 1 F18:A2:B1, 1 F5:A-:B1) and four were nontypeable. In conclusion, reduced susceptibility to FEP was mostly due to OXA-1 beta-lactamase. In E. coli, this increase is mainly due to the dissemination of two clones, which have captured different IncF plasmids. Among non-E. coli strains, five isolates produced OXA-1 and one isolate produced only TEM-1.
Introduction
F
Materials and Methods
A total of 33 clinical Enterobacteriaceae classified as isolates intermediate/resistant (minimum inhibitory concentration [MIC] ≥ 16 [amoxicillin] – 8 [clavulanate] mg/L) to AMC and with decreased susceptibility to FEP (MIC ≥ 2 mg/L) were collected at the Virgen Macarena Hospital (Seville, Spain) between 2011 and 2012 (27 E. coli, 4 Klebsiella pneumoniae, 1 Proteus mirabilis, and 1 Salmonella spp.). Twenty-one of the isolates (63.6%) were recovered from nonhospitalized patients, and 26 (78.8%) were isolated from urine. The Health area of the Virgen Macarena Hospital serves a population of some 450,000 people and is located in the north of Seville, Spain.
Identification and in vitro susceptibility to amoxicillin (AMX), AMC, PTZ, cefazolin (KZ), cefoxitin (FOX), cefuroxime (CXM), ceftazidime (CAZ), cefotaxime (CTX), FEP, imipenem (IP), meropenem (MEM), nalidixic acid (NAL), ciprofloxacin (CIP), gentamicin (GEN), tobramycin (TOB), and amikacin (AMK) were performed using the Wider commercial system (Francisco Soria Melguizo) and Clinical and Laboratory Standards Institute (CLSI) 2014 breakpoints were used. 7 Further confirmatory testing for FEP susceptibility was determined by the broth microdilution method, according to the CLSI guidelines. 7
The double-disk synergy test was used to screen for the production of OXA-1 beta-lactamase, using FEP and AMC. 2 Genes coding for blaOXA-1 were detected by polymerase chain reaction (PCR) amplification using specific primers, followed by further sequencing. Amplification and nucleotide sequencing were likewise performed for blaTEM genes. The genetic environment of the blaOXA-1 gene was determined by PCR with primers matching class 1 integrons (5′-CS, 5′-GGCATCCAAGCAGCAAG-3′; 3′-CS, 5′-AAGCAGACTTGACCTGA-3′). In addition, blaOXA-1 gene of isolate 12–155 was cloned into pCR-Blunt II-TOPO vector in E. coli DH10B and selected with 30 mg/L kanamycin and 0.25 mg/L FEP agar plates to ensure the genetic surrounding. PCR mapping assays were performed using 5′-CS plus blaOXA-1 reverse primers and 3′-CS plus blaOXA-1 forward primers. In isolates without class 1 integrons, specific primers for insertion sequence IS26 (IS26-F, 5′-AGCGGTAAATCGTGGAGTGA-3′; IS26-R, 5′-CAAAGTTAGCGATGAGGCAG-3′) were used along with blaOXA-1 primers.
Genetic relatedness of E. coli isolates was determined by pulsed-field gel electrophoresis (PFGE) using XbaI, according to Pulsenet protocol (www.pulsenetinternational.org/protocols/pfge.asp). Dendograms were created with Fingerprinting 3.0 software (BioRad), using the Dice coefficient with a position tolerance of 1%. Additional typing procedures for E. coli included multiplex PCR for assignment of phylogenetic groups, 4 screening of the ST131 clone (PCR of rfb O25b and allele 3 of the pabB gene), 5 and multilocus sequence typing (MLST) according to the University of Warwick Medical School (United Kingdom) scheme for E. coli (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli). A collection of 34 CTX-M-15-producing ST131 E. coli isolates from a previous study 14 was used to compare with OXA-1-producing ST131 E. coli isolates.
Extraction of plasmid DNA from all isolates was performed by the Kieser method. 12 Plasmid DNA extracts were also used in transformation experiments with a Gene Pulser II electroporation system (Bio-Rad) and selected on Luria-Bertani agar plates (LBA) containing FEP (0.25 mg/L). Transferability of the blaOXA-1 gene was studied by conjugation into sodium azide-resistant E. coli J53, selecting with 100 mg/L sodium azide and 0.25 mg/L FEP LBA. Plasmids from these transformants and transconjugants were rep typed and those plasmids harboring IncF replicons were subtyped according to a replicon sequencing scheme discriminating IncF variants, 18 and then assigned using plasmid MLST database (www.pubmlst.org/plasmid/).
Ten E. coli isolates were selected by MIC and phylogroup to evaluate the expression of the blaOXA-1 gene, using reverse transcription-PCR (RT-PCR), and to study Omps with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), using 10% acrylamide/0.27% bisacrylamide/6 M urea. The LightCycler-FastStar DNA Master SYBR Green I Kit (Roche) as well as OXA-1RT-F (5′-AGCAAAGTGTGCAACGCAAA-3′) and OXA-1RT-R (5′-TGCATCCACGTCTTTGGTGT-3′) primers were used for RT-PCR. Normalized expression levels of the target gene transcripts were calculated relative to the housekeeping gene mdh using the 2−ΔΔCT method.
Results
All isolates showed a similar susceptibility phenotype: resistant to penicillins and AMC and susceptible to third-generation cephalosporins and carbapenems. FEP MICs by broth microdilution ranged between 1 and 16 mg/L (Table 1). Resistance to other antimicrobials groups was variable, with NAL (75.8%) and CIP (72.8%) showing the highest rates of resistance. Regarding aminoglycosides, the resistance rates to GEN, TOB, and AMK were 27.3%, 42.4%, and 12.1%, respectively. There was clear synergy between clavulanate and FEP for all isolates. Twenty-nine (87.9%) isolates produced OXA-1 and 9 (27.3%) produced TEM-1. Five of these nine isolates produced both OXA-1 and TEM-1 beta-lactamases and four isolates produced just TEM-1. In 24 (82.7%) isolates, a class 1 integron surrounding blaOXA-1 (with the structure, int1-blaOXA-1-aadA1-qacEΔ1-sul1) was detected. In five isolates, an IS26 structure was detected.
Antimicrobials were tested by wider system, but MIC of cefepime was confirmed by broth microdilution method.
Cefepime MICs for transformants and transconjugants were determined by broth microdilution method.
AMC, amoxicillin/clavulanate; FEP, cefepime; SFEP, cefepime; Int 1, class 1 integron; IS, internal sequence; MIC, minimum inhibitory concentration; ND, not done; NT, nontransferable by conjugation or transformation; PTZ, piperacillin/tazobactam; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; TC, plasmid obtained by conjugation; TF, plasmid transferred by transformation.
Of the 27 E. coli isolates, 24 (89%) produced OXA-1. The main phylogroup was C (67%), followed by the B23 phylogroup (25%; Table 1). By PFGE, nine different patterns were obtained from 24 OXA-1-producing E. coli (dice similarity value >80%). These patterns were grouped into two main clusters: cluster 1 (72% similarity) consisted of 16 OXA-1-producing isolates of phylogroup C corresponding to ST88 and cluster 2 (80% similarity) with five isolates of phylogroup B23 and serotype O25b corresponding to ST131 (Fig. 1). These five OXA-1-producing ST131 isolates were compared with previous CTX-M-15-producing ST131 isolates and a cluster (100% similarity) was found, which included one OXA-1 producer and two CTX-M-15 producers (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr).
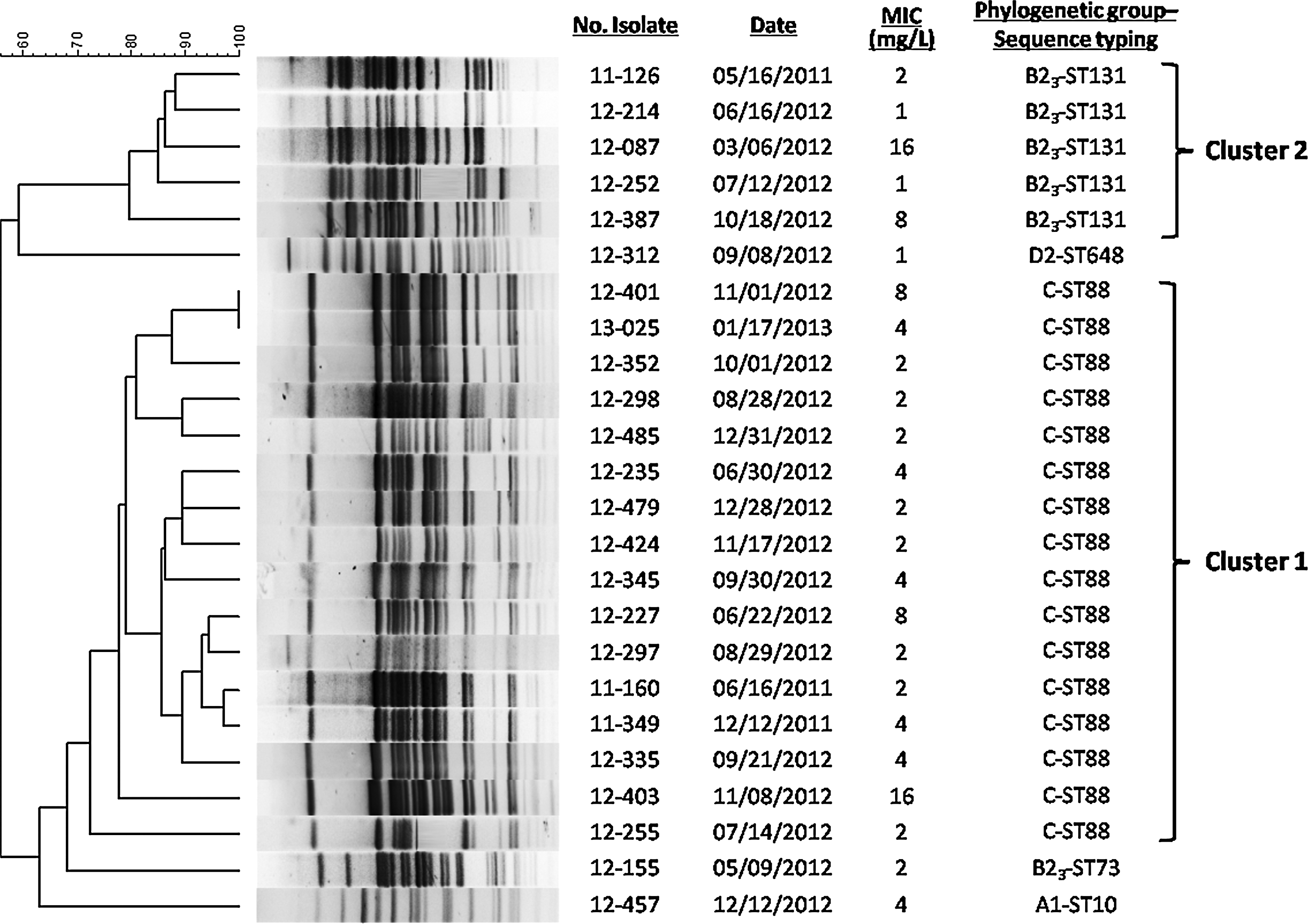
Dendogram of pulsed-field gel electrophoresis patterns of 24 OXA-1-producing Escherichia coli isolates. Minimum inhibitory concentration of cefepime was determined by broth microdilution.
Of the remaining six non-E. coli species, four isolates of K. pneumoniae and one isolate of Salmonella spp. produced OXA-1 (all of these found in the class 1 integron) and one isolate of P. mirabilis produced only TEM-1 (Table 1). These isolates were recovered from four adults and one child (<1 year old), who had suffered previous recurrent urinary tract infections (UTIs), and one Salmonella enterica from a pediatric gastroenteritis episode. All patients with UTIs had received AMC treatment in the previous year.
Plasmids between 66 and 154 kb were detected in 26 isolates of the 29 OXA-1-producing isolates (89.7%). Thirteen plasmids containing blaOXA-1 were transferred, four by conjugation, seven by transformation, and two by both mechanisms of transfer. In the transformants-transconjugants, a single plasmid of around 154 kb was found. Eight of them were included in a class 1 integron. When the MIC for a transformant or transconjugant was lower than the parent isolate (six isolates), additional mechanisms of resistance were found in the parent strain such as additional presence of blaTEM-1 (three isolates) or loss of both OmpC and OmpF porins (two isolates), and one isolate TEM-1 negative was not selected for porins study (Table 1). The cloned blaOXA-1 gene also showed 32-fold higher FEP MIC values (2 mg/L) compared to E. coli DH10B (0.06 mg/L). Nine transferable plasmids belonged to IncF replicon types (six of these isolates belonged to the B23 phylogroup, of which four vehiculized blaOXA-1 without a class 1 integron) and four were nontypeable (Table 1).
RT-PCR did not show that isolates with increased FEP MICs had higher expression of blaOXA-1 than those with lower MICs (data not shown). In 9 of the selected 10 E. coli isolates, OmpC porin was not expressed in the SDS-PAGE analysis. In three isolates, there was no expression of OmpF or OmpC (Supplementary Fig. S2).
Discussion
The results of this study show that the increased prevalence of oxyimino-cephalosporins susceptibility and decreased susceptibility to AMC and FEP among Enterobacteriaceae clinical isolates over 2 years were mainly due to the presence of blaOXA-1 genes. This reduced susceptibility to FEP was reproduced in both transferable plasmids and cloned blaOXA-1. This phenotype has been described previously for sporadic E. coli strains. 2
According to other authors, blaOXA-1 genes are part of a class 1 integron 1 and are mainly present in E. coli. In a previous national study in our country, Ortega et al. 15 detected an increase in OXA-1 production (26.1%) in clinical isolates collected in Spain between January and May 2010. In this study, isolates were selected according to AMC susceptibility (MIC ≥ 32/16, or ≤13 mm inhibition zone) and additional information of FEP susceptibility was not available. In 2010, 32 E. coli were collected for the national survey of Ortega et al. and none of them produced OXA-1 without CTX-M-15. A national study carried out between 1997 and 2008 in Spain showed an increase in the use of AMC in the community 8 and more recently, in our health area, the increasing trend remained in outpatients. 11 Further studies would be needed to associate outpatient antibiotic use and the emergence of these clones. Dissemination of OXA-1 beta-lactamase in E. coli isolates in our area is mainly due to clonal spread of two clones, the epidemic ST131 clone and the ST88 phylogroup C clone, as previously described. 15 In addition, this determinant is spreading among other Enterobacteriacea as Salmonella and Klebsiella, and was located within a class 1 integron, as previously published.3,16 To date, plasmids harboring OXA-1 lacking other beta-lactam resistance determinants have not been characterized. In our study, different IncF plasmids harboring blaOXA-1 were even detected in the same clone, although four ST131 isolates (67% of OXA-1-producing ST131 isolates) carried F2:A1:B- plasmids. In a multicenter collection of ST131 extended spectrum beta-lactamase (ESBL)-producing E. coli from the United Kingdom, Doumith et al. 9 found that 13/15 isolates harboring F2:A1:B- plasmids contained both blaCTX-M-15 and blaOXA-1.
There was a significant difference between the CLSI and European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints for susceptibility to FEP in 2013 (susceptible at MIC ≤ 8 mg/L and MIC ≤ 1 mg/L, respectively).6,10 As a result, in 2014, CLSI changed the susceptible category to MIC = 2 mg/L and added a new interpretive category, the susceptible-dose dependent (SDD; MIC range: 4–8 mg/L), in which the susceptibility of an isolate is dependent on the dosing regimen used. 7 Among the OXA-1 producers, discrepancies were found between the commercial panel and the microdilution results, particularly with respect to EUCAST breakpoint values. In our series of OXA-1 producers, 86% were intermediate and 10% were resistant with the commercial panel, while 48% were intermediate and 24% were resistant using microdilution according to EUCAST. On the other hand, 45% were SDD and none were resistant using the commercial panel, and 38% were SDD and 3% were resistant using microdilution according to CLSI.
According to our data, the combination of different resistance mechanisms, such as porin loss (mainly OmpC), expression of blaOXA-1, and/or the presence of TEM-1, could confer decreased susceptibility to FEP. This was demonstrated with transformable OXA-1 isolates; when the FEP MICs for these were lower than for the clinical parent isolates, blaTEM-1 and/or porin loss were present in the parent cell. As an example, in an isolate with a FEP MIC of 8 mg/L (2 mg/L for transferable plasmids), the presence of an OXA-1 enzyme was combined with the loss of both OmpF and OmpC porins; an isolate with a FEP MIC of 1 mg/L (0.5 mg/L for transformant) presented OXA-1 and TEM-1.
Some limitations of our study should be noted. Few plasmids could be characterized due to the fact that nontransformants/nontransconjugants were obtained. We were unable to find a direct relationship between the expression of blaOXA-1, addition of blaTEM-1 or porin loss, and FEP MIC value in the 10 selected isolates. It should be borne in mind that these differences would be associated with different copy numbers or with other additional mechanisms.
In conclusion, an increase of Enterobacteriaceae isolates with reduced susceptibility to FEP was observed in our area, mostly due to OXA-1 beta-lactamase and particularly when it was combined with other resistance mechanisms. In E. coli, this increase is mainly due to the dissemination of two clones, which have captured different IncF plasmids. In our area, these two E. coli clones along with other non-E. coli species harboring blaOXA-1 gene could have contributed to its dissemination.
Footnotes
Acknowledgments
This work was supported by Plan Nacional de I + D + i 2008–2011 and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Economía y Competitividad, and Spanish Network for Research in Infectious Diseases (REIPI RD12/0015)—cofinanced by European Regional Development Fund ERDF “A way to achieve Europe”.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.