
Abstract
We analyze the effect of amikacin, ciprofloxacin, and clarithromycin, alone and associated with N-acetylcysteine (NAC) and Tween 80, at different times and concentrations in nonpigmented rapidly growing mycobacteria (NPRGM) biofilms. For this purpose, confocal laser scanning microscopy and image analysis were used to study the development and behavior of intrinsic autofluorescence, covered area, thickness, and cell viability in NPRGM biofilms after adding antibiotics alone and associated with antibiofilm agents. In this study, ciprofloxacin is the most active antibiotic against this type of biofilm and thickness is the most affected parameter. NAC and Tween 80 combined with antibiotics exert a synergistic effect in increasing the percentage of dead bacteria and also reducing the percentage of covered surface and thickness of NPRGM biofilms. Tween 80 seems to be an antibiofilm agent more effective than NAC due to its higher reduction in the percentage of cover surface and thickness. In conclusion, the results obtained in this work show that phenotypic parameters (thickness, percentage of covered surface, autofluorescence, percentage of live/dead bacteria) are affected by combining antibiotics and antibiofilm agents, ciprofloxacin and Tween 80 being the most active agents against NPRGM biofilms.
Introduction
B
In the last decades, there has been a marked increase in the interest in nontuberculous mycobacterial infections. They are two types of infections: community-acquired infections and nosocomial infections (acquired at the hospital), including traumatic and postsurgical infections, respectively, related to several invasive procedures, such as cosmetic surgery (abdominoplasty, liposuction, breast reduction or augmentation, mammoplasty), 8 contamination of medical equipment, or poststeroid injection abscesses and bronchoscope-associated infections. Nonpigmented rapidly growing mycobacteria (NPRGM) produce a broad spectrum of human diseases (pulmonary, otitis, injection abscess, surgery-associated infections, and bacteremia). The species implicated in those are primarily Mycobacterium fortuitum, Mycobacterium chelonae, and Mycobacterium abscessus, with occasional reports of diseases caused by other species of this group. 4 These organisms are ubiquitous in nature and can be found in soil, water, and even disinfectant solutions or biocides such as the members of the M. chelonae/abscessus group. 4 They survive harsh conditions (low nutrients, low pH, and extreme temperatures) 8 and can be isolated from biofilms, dusts, and aerosols. 12 Risk factors associated with NPRGM include pre-existing pulmonary disease, 20 cystic fibrosis, 1 or immunosuppression. 6
NPRGM are resistant to a broad spectrum of antibiotics and disinfectants due to their hydrophobicity, impermeability, and slow growth.3,11 In a previous report, we have demonstrated the increasing resistance of RGM embedded in a biofilm 15 and the effect of ciprofloxacin in the ultrastructure of these biofilms. 16 If we considered that the ability of biofilm development is broadly distributed among these mycobacteria, 14 new strategies are necessary to improve the efficacy of the therapy and outcome of the patients with infections caused by these organisms. It is recommended by the Clinical and Laboratory Standards Institute (CLSI) to test nine drugs to measure the antimicrobial susceptibility of rapidly growing mycobacteria, specifically amikacin, cefoxitin, imipenem, trimethoprim-sulfamethoxazole, clarithromycin, ciprofloxacin, doxycycline, linezolid, and tobramycin. It is known also that disinfectants are effective against mycobacteria when used at the appropriate concentration. 2 It is also described in the literature that detergents such as Tween 80 (0.1–1.0%) can reduce aggregation of these organisms. 12 In this study, we analyze the effect of ciprofloxacin, amikacin, and clarithromycin and their combination with N-acetylcysteine (NAC) and Tween 80 on the growth and ultrastructure of mycobacterial biofilms during their development. The broad objective of the investigations is to identify and understand the factors involved in mycobacterial biofilm formation through employment of in vitro models.
Materials and Methods
Type strains of 4 species of RGM, M. abscessus DSM 44196, M. chelonae ATCC 19235, M. fortuitum ATCC 6841 (these three are the most common human pathogens of this group), and Mycobacterium smegmatis ATCC 607, were selected for this study.
To analyze the actual antimicrobial activity of Tween 80 (BD) and NAC (Sigma) against planktonic bacteria, we performed a standardized microdilution technique for rapidly growing mycobacteria as specified by CLSI, 21 using the agents instead of antibiotics. Minimal inhibitory concentrations (MIC) were determined using serial concentrations of NAC (0.03–4096 mg/L) and Tween 80 (0–40%) in sterile P-96 microplates (Corning). Each plate contained two wells for growth control and two wells without bacteria as a sterility control of the medium with Mueller-Hinton II (Difco). Tested mycobacteria were the same strains that were further studied in the biofilm assays. Quality controls were performed with Staphylococcus aureus ATCC 29213 and Mycobacterium peregrinum ATCC 700686, as suggested by the CLSI protocol. These studies were performed in triplicate.
To determine the minimum biofilm eradication concentration (MBEC), the biofilm was developed during a period of 96 hr in triplicate following the Calgary system with 96-well plates, MBEC™ Biofilm Inoculator (Innovotech), 5 and then tested against antibiotics, antibiofilm agents, or both combined. To evaluate the effect of the combination of antibiotics with antibiofilm agents, we exposed antibiotics at double concentrations (0.06–8192 mg/L) to NAC or Tween 80 during a period of 48 hr to know the effect in biofilms of 0.5% of Tween 80 and 512 mg/L of NAC. Data obtained from the combination of antibiotics and NAC or Tween 80 were compared with those previously published. 15 A minimum of two dilutions between the results of the antibiotic and those obtained of combinations were needed to consider it a difference attributable to the combination.
Biofilm development was analyzed at 24, 48, 72, and 96 hr using hydrophobic, uncoated, sterile slide 2- by 4-well plates (ibidy GmbH Martinsried), using a previously described protocol 17 with modifications as follows. Mycobacterial colonies were resuspended in sterile phosphate-buffered saline solution (PBS) (bioMérieux) to achieve a cell density of 1.5×108 CFU/ml. Three hundred microliters of this suspension was inoculated on each well. Inoculated slides were incubated at 37°C in a 5% CO2 atmosphere for 30 min. The suspension was then removed, and the wells were washed once with PBS. All the experiments were performed in parallel with and without antibiotic and antibiofilm agents. Three hundred microliters of Middlebrook 7H9 broth (7H9) (BD) was added at 24, 48, 72, and 96 hr after bacterial inoculation (a well without antibiotic as a control). The study with antibiotics was developed in different plates for each antibiotic. We studied the effect of the following concentrations of antibiotics at 24, 48, and 72 hr after bacterial inoculation: ciprofloxacin (CIP) (Sigma) 1 mg/L, amikacin (AN) (Sigma) 16 mg/L, and clarithromycin (CLR) (Sigma) 2 mg/L. The slides were placed on an orbital shaker (80 rpm) and incubated at 37°C in normal atmosphere for 4 days. These data were compared with those previously published with ciprofloxacin using identical methodology. 16
On the other hand, we studied antibiotics combined with antibiofilm agents at 24, 48, and 72 hr after bacterial inoculation. In all studies, the medium was replaced daily and maintained until the end of the experiment. The concentrations of the antibiotics and antibiofilm agents were CIP 1 mg/L, AN 16 mg/L, and CLR 2 mg/L, and 0.5% for Tween 80 and 512 mg/L for NAC. One well without antibacterial was used as a control (96 hr) in all the experiments. The concentrations were selected according to the described breakpoints for these organisms in the CLSI guidelines. 21 Slides were examined, and the medium, with and without antibiotic, was changed daily. The slide wells were stained using Live/Dead BacLight© stain (Invitrogen) and Nile Red© stain (Sigma-Aldrich Co.). Stains were performed according to the instructions provided by the manufacturer. At the end of the experiment (at 96-hr incubation) and after staining, autofluorescence, covered surface, live/dead bacteria, and thickness were analyzed using a Leica DM IRB confocal laser scanning microscope (Leica), and the photographs obtained were analyzed with the ImageJ software (National Institutes of Health), as being the % of dead bacteria, % of covered surface, and % of autofluorescence calculated. All the experiments were performed in triplicate for each strain, and the measures were performed on biofilms developed at the previously reported times of development.
One set of wells was used per NPRGM species to study both autofluorescence and Nile Red stain and the other was used to analyze the proportion of live and dead mycobacteria. All materials managed in the experiments emitted no autofluorescence. The covered surface was studied by taking 96 microphotographs for each stain, bacterium, and time set. Photographs were analyzed as previously described. 10 The thickness of the biofilm was measured in eight predefined points/well. Autofluorescence was analyzed as the percentage of fluorescence related to the number of bacterial cells detected with the Nile Red stain using the following formulation: (% autofluorescence of covered surface/% Nile Red covered surface) ×100.
The values of autofluorescence, percentage of covered surface, percentage of deaths, and thickness were measured and compared between the different groups regarding antibiotic and antibiofilm agent administration times (24, 48, 72 hr). Groups were compared by pairs using Mann–Whitney test with a significance level of 0.05. Separate comparisons were performed for each species. EPI INFO 3.5.1. epidemiological software (Centers for Disease Control and Prevention) was used for statistical calculations. Data obtained with ciprofloxacin were previously published for these and other species of RGM 16 and were used for comparison purposes, as well as those of susceptibility testing of antibiotics against biofilms formed by the same species. 15
Results
Sensitivity studies on planktonic bacteria and biofilms
MIC and MBEC values of antibiofilm agents (NAC and Tween 80) are listed in Table 1 and compared with those previously published with the different antibiotics. 15 Taking the CLSI reference values, in this study, we see that the MIC for CIP in the case of M. abscessus is intermediate (MIC 2 mg/L). 21 NAC and Tween 80 did not show antibacterial properties when used alone. However, NAC and Tween 80 at much lower concentrations (512 mg/L for NAC and 0.5% for Tween 80) and in combination with antibiotics showed a two-dilution reduction in the MBEC in M. fortuitum and M. smegmatis. All species showed high resistance to CLR of ≥4096 mg/L.
AN, amikacin; CIP, ciprofloxacin; CLR, clarithromycin; MIC, minimal inhibitory concentration; MBEC, minimum biofilm eradication concentration; NAC, N-acetylcysteine.
Determination of phenotypic characteristics by confocal laser scanning microscope
The use of antibiotics with antibiofilm agents (NAC and Tween 80) showed greater variations in the thickness, percentage of covered surface, and percentage of dead bacteria of RGM biofilms than the use of antibiotics alone. In that sense, the thickness and the percentage of covered surface were significantly reduced (especially with the combination of ciprofloxacin and Tween 80), and the percentage of dead bacteria was significantly increased. The percentage of autofluorescence seemed to be an independent parameter (Figs. 1–4 and Supplementary Tables S1–S4; Supplementary materials are available online at http://www.liebertpub.com/mdr).
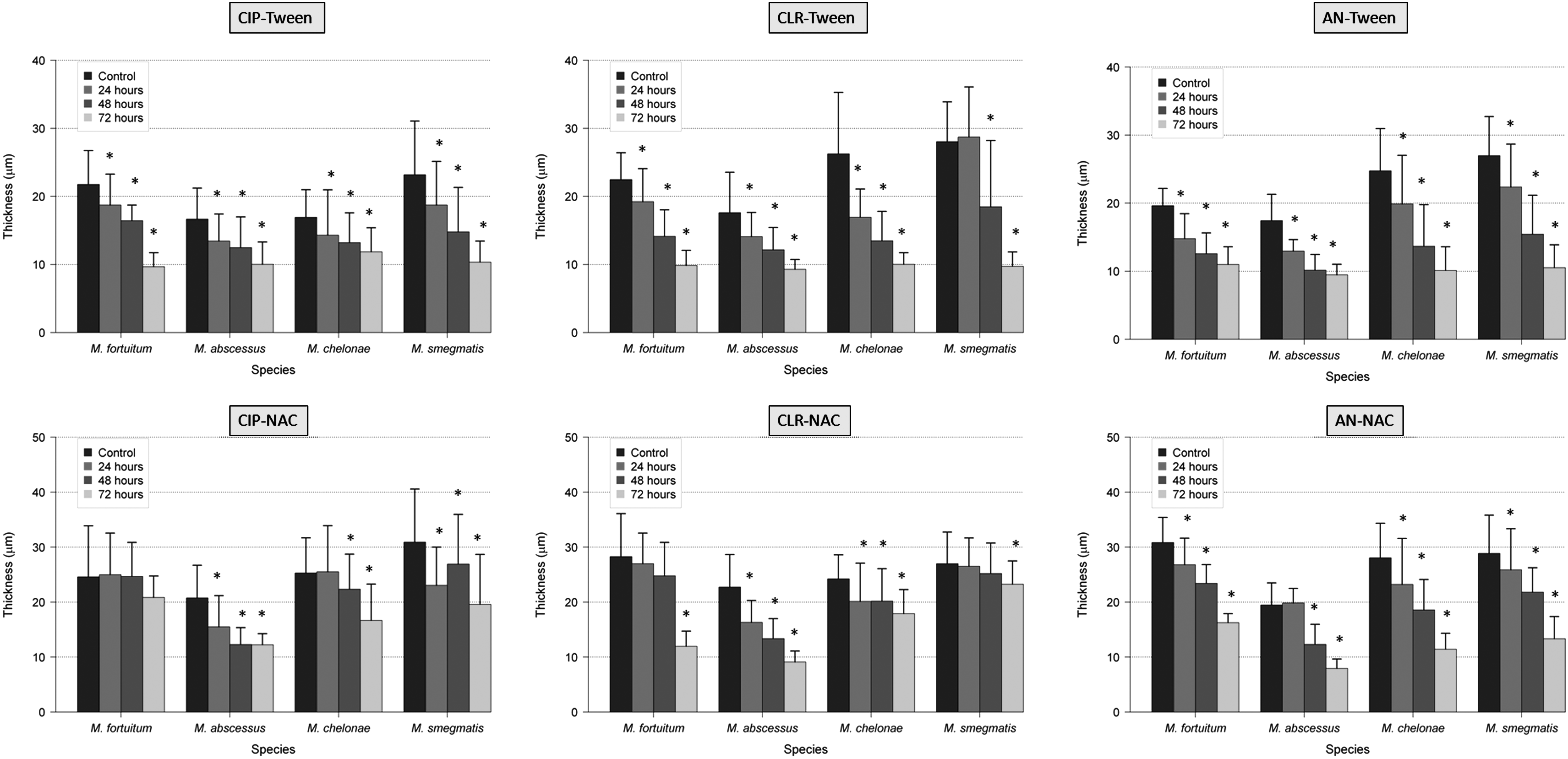
Effect of antibiotics and antibiofilm agents (Tween 80 and NAC) in the thickness of RGM biofilms (*<0.05 statistical significance; compared with control 96 hr). AN, amikacin; CIP, ciprofloxacin; CLR, clarithromycin; NAC, N-acetylcysteine; RGM, rapidly growing mycobacteria.
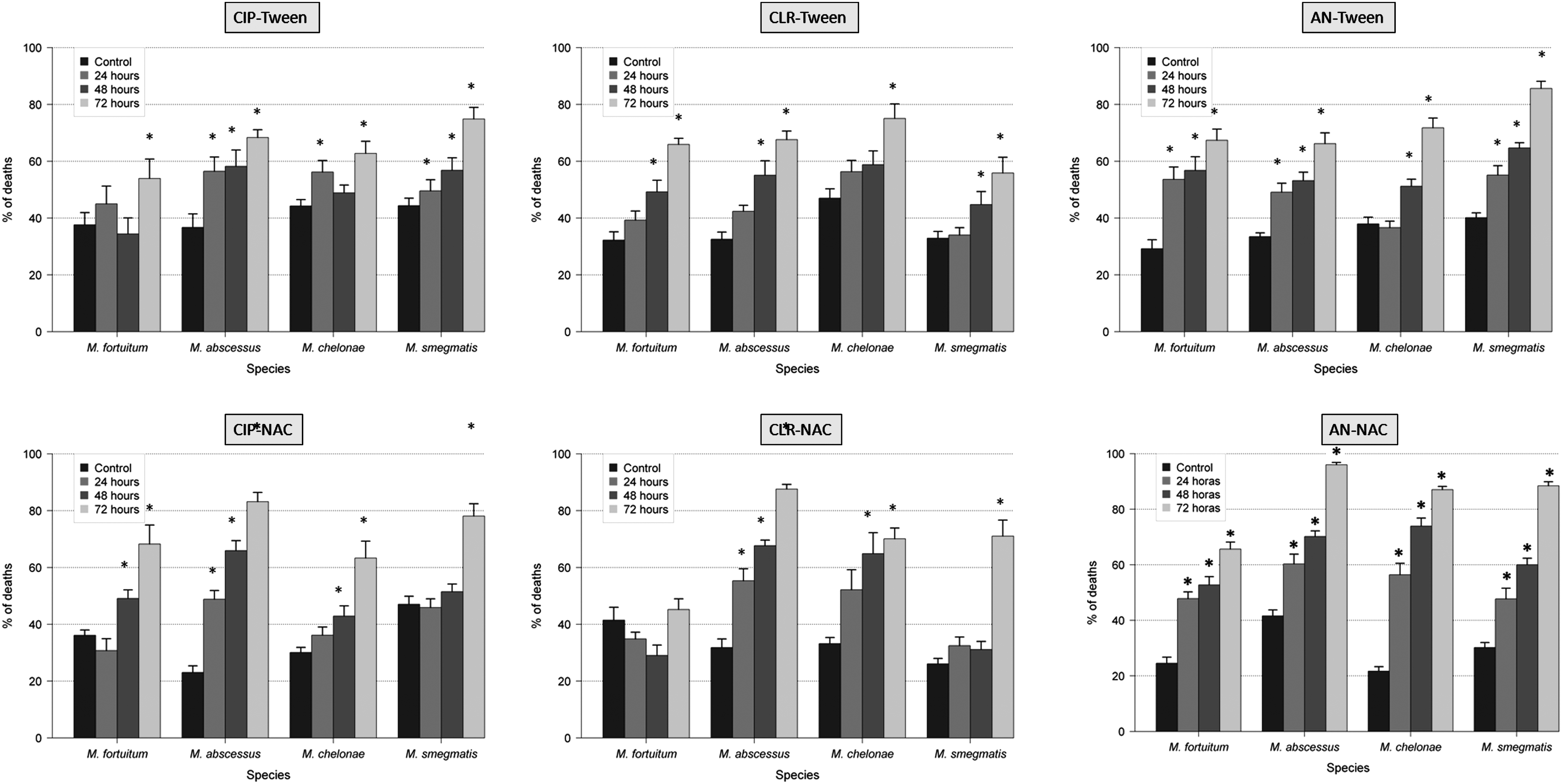
Effect of antibiotics and antibiofilm agents (Tween 80 and NAC) in the percentage of dead bacteria of RGM biofilms (*<0.05 statistical significance; compared with control 96 hr).
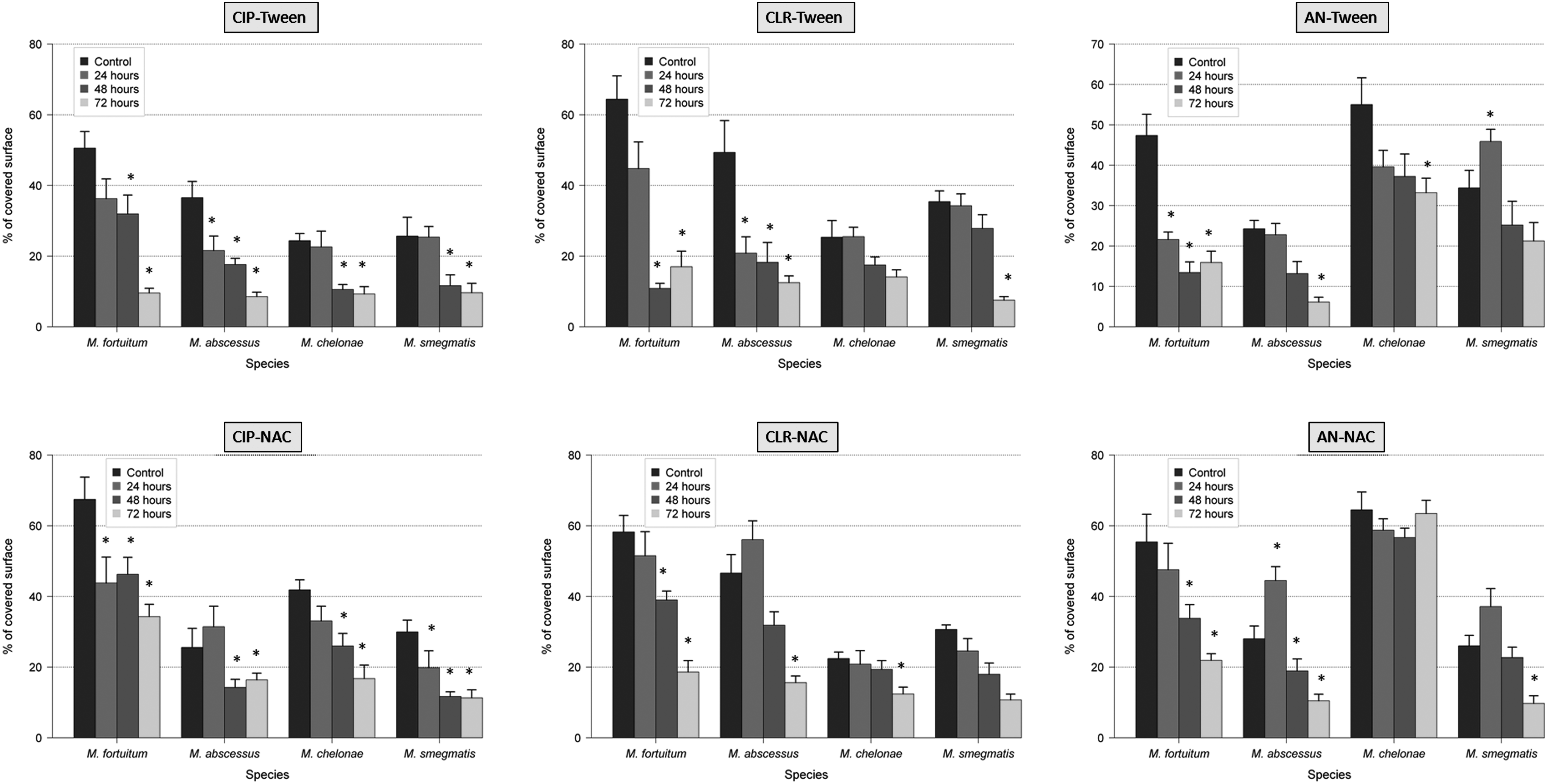
Effect of antibiotics and antibiofilm agents (Tween 80 and NAC) in the percentage of covered surface of RGM biofilms (*<0.05 statistical significance; compared with control 96 hr).
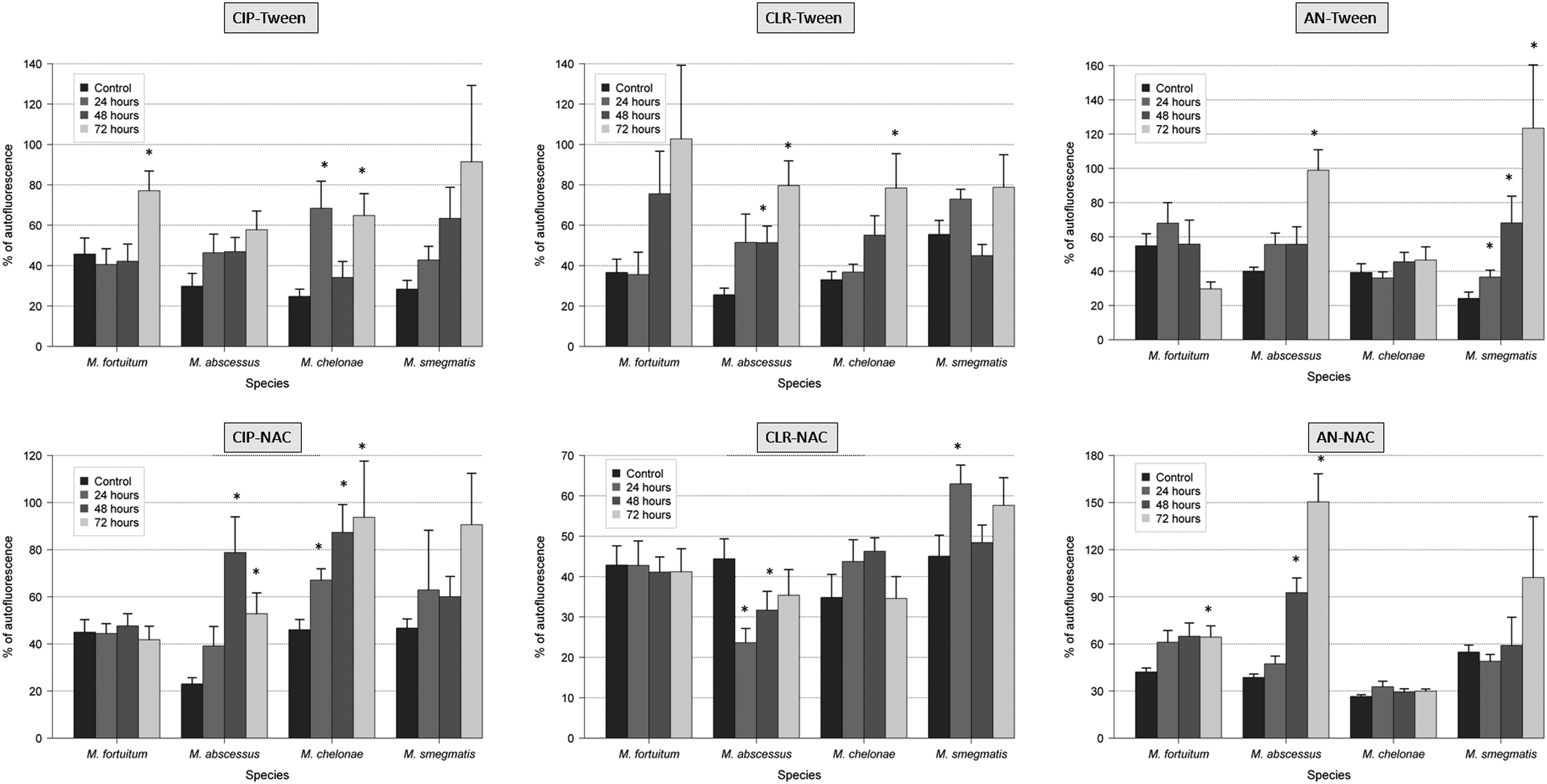
Effect of antibiotics and antibiofilm agents (Tween 80 and NAC) in the autofluorescence of RGM biofilms (*<0.05 statistical significance; compared with control 96 hr).
Discussion
Despite the absence of a standardized method to study the antimicrobial susceptibility in biofilms, the Calgary method is a technique that seems to be the most appropriate because it is highly reproducible. 5 In a previous report, the antimicrobial resistance against tested antimicrobials was higher in all the NPRGM studied when they are part of a biofilm. 15 We also described that ciprofloxacin was the most active against NPRGM biofilms (MBEC), while clarithromycin and amikacin showed practically no activity against them. 15 The effect of ciprofloxacin against these biofilms was also analyzed in detail, showing differences between studied species in different parameters of biofilm development. 16 In this study, we added to these previous reports the evaluation of supposed antibiofilm molecules (NAC and Tween 80) and the effect of the combination of these substances with previously tested antimicrobials. Moreover, the effect of these antibiotics (alone and in combination with these molecules) is currently evaluated for their effect on different parameters of biofilm structure using the methodology previously reported only for quinolones.
The spectrum of resistance of mycobacteria to antibiotics and disinfectants has been reported to be caused, at least in part, by their growth rate, hydrophobicity, and impermeability, 3 together with specific mechanisms of resistance (especially well known for macrolides and quinolones). 9 As expected, the percentage of dead bacteria after antibiotic exposure increases especially after 48 and 72 hr, except in M. fortuitum and M. chelonae with amikacin. This effect may be related to less activity of aminoglycosides in biofilms. In the case of M. fortuitum and M. chelonae after 24 hr of exposure with ciprofloxacin, 16 M. fortuitum with clarithromycin, and M. chelonae and M. fortuitum with amikacin, the average of the percentage of dead bacteria was slightly lower than the controls. Among the hypotheses is included the maturity of the biofilm when the antibiotic is added because bacteria in biofilms are metabolically inactive and more resistant to antibiotics than those metabolically active. When we compare the average results of percentage of covered surface, this parameter is especially reduced after 48 and 72 hr of antibiotic exposure. Interestingly, when controls are compared with strains treated 24 hr with antibiotic, this parameter does not vary substantially or even increases. Perhaps during the early stages of biofilm formation, adhesion phase and irreversible binding, it is more difficult to separate the cells from the surface. Regarding the percentage of autofluorescence, we can say that this is an independent parameter as it decreases or increases in some species and some antibiotics, but not following a definite tendency.
The multiple resistances to antibiotics promote the development of new therapeutic options. In this study, we analyze if there was a synergistic effect combining NAC and Tween 80 with different antimicrobials because there is no experience of these agents in biofilms formed by mycobacteria. In our experiments, NAC and Tween 80 increased antibacterial activity when they were combined with all antibiotics. We also observed a reduction in the MBEC of M. fortuitum with amikacin/NAC and in M. smegmatis with ciprofloxacin/Tween, amikacin/Tween, ciprofloxacin/NAC, and amikacin/NAC. This synergistic effect may have the potential to use them in the prophylaxis or treatment of infections associated with biofilms.
We also observed by confocal laser scanning microscope how ciprofloxacin, clarithromycin, and amikacin in combination with Tween and NAC affect the ultrastructure of biofilms in M. fortuitum, M. chelonae, M. abscessus, and M. smegmatis. In most cases, the percentage of dead bacteria is higher with a combination of antibiotics and antibiofilm agents than with only antibiotics. On the other hand, the percentage of autofluorescence does not follow a clear trend, so we consider it as an independent parameter. Note the presence of extracellular autofluorescence in M. smegmatis with combinations of amikacin/Tween and amikacin/NAC, and in M. abscessus with amikacin/NAC, and in M. fortuitum with amikacin/Tween. The actual significance of this phenomenon is currently unknown.
In all the experiments, the combinations of antibiotic with the detergent Tween 80 showed a higher reduction in the percentage of covered surface and thickness than NAC. We hypothesize that these results could be attributable to the mechanism of action of these agents and their effect on the biofilm. Tween 80 has the potential to alter the structural integrity of the membrane, lipids, and proteins, 19 while NAC acts on the polysaccharide matrix of the biofilm, breaking the disulfide bridges that link the polysaccharide fibers. 18 Due to the high lipid content of the mycobacterial cell wall and their presence in the extracellular matrix, we can understand why Tween 80 is a more active antibiofilm agent against mycobacterial biofilms than NAC.
We have clearly observed that antibiotics and antibiofilm agents affect the phenotypic features of NPRGM biofilms in a synergistic manner. This synergistic effect highlights the potential of these antibiofilm agents for use in the prophylaxis or treatment of infections associated with biofilms.
Footnotes
Acknowledgments
This study was partially communicated (Ref: PO927) at the 24th European Congress of Clinical Microbiology and Infectious Diseases (ECCMID) in Barcelona, Spain, May 11, 2014. The authors acknowledge Sylvia Powell for her help with the language of the manuscript and Mar Gonzalez Garcia-Parreño for her help with confocal microscopy studies.
This work was supported by a grant from the Spanish Ministry of Science (FUNCOAT CSD 2008-00023) and Spanish MINECO MAT2013-48224-C2-2-R.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.