
Abstract
Klebsiella variicola, a bacterium closely genetically related to Klebsiella pneumoniae, is commonly misidentified as K. pneumoniae by biochemical tests. To distinguish between the two bacteria, phylogenetic analysis of the rpoB gene and the identification of unique genes in both bacterial species by multiplex-polymerase chain reaction (PCR) provide the means to reliably identify and genotype K. variicola. In recent years, K. variicola has been described both as the cause of an intrahospital outbreak in a pediatric hospital, which resulted in sepsis in inpatients, and as a frequent cause of bloodstream infections. In the present study, K. pneumoniae and K. variicola were isolated from a unique patient displaying different antimicrobial susceptibility phenotypes and different genotypes of virulence determinants. Eight clinical isolates were obtained at different time intervals; all during a 5-month period. The isolates were identified as K. pneumoniae by an automated identification system. The clinical (biochemical test) and molecular (multiplex-PCR and rpoB gene) characterization identified imipenem resistance in the first six K. pneumoniae ST258 isolates, which encode the SHV-12 cephalosporinase and KPC-3 carbapenemase genes. The two last remaining isolates corresponded to susceptible K. variicola. The bacterial species showed a specific profile of virulence-associated determinants, specifically the fimA, fimH, and ecpRAB fimbrial-encoding genes identified only in K. pneumoniae isolates. However, the entb (enterobactin), mrkD (fimbrial adhesin), uge (epimerase), ureA (urease), and wabG (transferase) genes were shared between both bacterial species. Recent studies attribute a higher mortality rate to K. variicola than to K. pneumonia. This work highlights the identification of K. pneumoniae and the closely related K. variicola isolated from the same patient. The value of distinguishing between these two bacterial species is in their clinical significance, their different phenotypes and genotypes, and the fact that they can be isolated from the same patient.
Introduction
K
Likewise, K. variicola has been associated with clinical bovine mastitis infections. 21 A general misidentification results from the use of currently available biochemical techniques that are not able to identify K. variicola among K. pneumoniae isolates. 15 Reports concerning K. variicola causing infections in humans have been increasing in recent years. These two species can be differentiated only by either rpoB gene analysis or a recently described multiplex-polymerase chain reaction (PCR) method. The latter is based on the amplification of unique genes identified by genomic comparison of K. variicola and K. pneumoniae.15,23
The lack of a biochemical test and the fact that Matrix-Assisted Laser Desorption Ionization Time of Flight (MALDI-TOF) technology was discarded, due to an incorrect identification of species among K. variicola and K. pneumoniae. 29 This limits the options for distinguishing K. variicola from K. pneumoniae in the same patient and as a result has not been described. The current study describes the identification of K. pneumoniae and K. variicola in the same patient at different time intervals and further clinically and molecularly characterizes the mechanisms of antimicrobial resistance and virulence factors of both bacterial species.
Materials and Methods
Clinical isolates
A total of eight clinical isolates were obtained from a nosocomial infection in a 74-year-old female patient admitted at the “Dr. Manuel Gea Gonzalez” General Hospital in Mexico City. The patient was hospitalized from January to May of 2010. During this period, six clinical cultures were obtained from wound purulent secretions and two from blood.
Biochemical and molecular identification of clinical isolates
The clinical isolates were identified using the automated MicroScan Walkaway system (Dade Behring, West Sacramento, CA). The isolates were sent to the National Institute of Public Health (www.insp.mx) for molecular characterization. The molecular test, multiplex-PCR using the M-PCR-1 amplification condition, was used to distinguish K. variicola from K. pneumoniae isolates. 15 In addition, phylogenetic analysis of the rpoB gene (using 501 bases) was carried out using the primers described for K. pneumoniae rpoB-multilocus sequence typing (MLST). 8
Extended-spectrum β-lactamase production, carbapenemase activity, and susceptibly testing
Extended-spectrum β-lactamase (ESBL) production was assayed by double-disc synergy according to Clinical Laboratory Standard Institute (CLSI). 7 The carbapenemase activity was determined by the Carba NP-test. 9 The minimal inhibitory concentrations were performed by broth microdilution for ceftazidime (CAZ), cefotaxime (CTX), piperacillin (PIP), ciprofloxacin (CIP), gentamicin (GEN), imipenem (IMP), meropenem (MER), tigecycline (TGC), and colistin (COL), following CLSI recommendations. 7
Pulsed-field gel electrophoresis and MLST analyses
The genetic relatedness of all isolates was examined by pulsed-field gel electrophoresis (PFGE), 28 and the results were analyzed using GelCompar II software (Applied Maths, Kortrijk, Belgium). K. pneumoniae MLST was conducted for the K. pneumoniae isolates according to Diancourt et al. 8
Identification of ESBL and carbapenemase genes by PCR
The ESBL and carbapenemase genes were screened by PCR for the SHV- and CTX-M- 27 or KPC- 12 and NDM-type 3 genes, respectively. All PCR products were sequenced using the chain termination method with a Big-Dye Terminator kit (Applied Biosystems, Foster City, CA) and ABI PRISM 3100 (Applied Biosystems). The nucleotide sequences were analyzed by BLASTx (www.ncbi.nlm.nih.gov).
Analysis by PCR of virulence determinants and hypermucoviscous phenotype
The following virulence-determinant genes were screened by PCR (under standard conditions) in all K. pneumoniae and K. variicola isolates using specific primers (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr): entB (enterobactin), iroB (salmochelin), irp2 (yersiniabactin), iucA (aerobactin), fimA (fimbrial), fimH (fimbrial), ecpA (pilin subunit), ecpRAB (fimbrial), mrkD (fimbrial), mrkA (fimbrial), uge (uridine diphosphate galacturonate-4 epimerase), ureA (urease), allS (allantoinase), kfuBC (siderophore), cf29a (adhesin), and wabG (glucuronic acid transferase).1,5,24,31 The hypermucoviscous phenotype of K. pneumoniae and K. variicola isolates was analyzed by semi-quantitative string tests. 11
Plasmid analyses, mating experiments, and southern hybridizations
Plasmid DNA profiles were obtained from all isolates according to the method described by Kieser. 16 Escherichia coli NCTC 50192 plasmids 154-, 66-, 48-, and 7-kb were used as molecular size markers. 19 Carbapenem resistance transfer was performed by mating according to Miller 17 using the E. coli J53 strain. Southern hybridization experiments were conducted using a nonradioactive probe (ECL direct nucleic acid labeling and detection system; GE Healthcare, Piscataway, NJ) obtained by PCR amplification from the entire SHV-12 and KPC-3 genes.
Results and Discussion
Clinical patient history and clinical isolates
A 74-year-old female patient was admitted to the emergency room with the following symptoms: constipation for the previous 4-days, nausea, vomiting, and abdominal pain without fever. She was diagnosed with an intestinal obstruction and chronic constipation. A derivative gastrostomy was performed through laparotomy and peritoneal adhesions were resolved. The patient had a satisfactory evolution and was discharged from the hospital after 5 days. However, 4 days later, she returned to the emergency room in septic shock due to a bowel perforation. An intestinal resection (∼60 cm in length) was performed, followed by placement of a jejunostomy feeding tube. The patient, in respiratory failure and without fever, was transferred to the intensive care unit. Treatment with metronidazole 500 mg/day, metamizol 1 g/day for 10 days, and ceftazidime 1 g/day for 2 days was initiated. The third day was substituted by cefuroxime 759 mg/day for the next 7 days. The patient remained in respiratory failure and developed fever. Severe continuous secretion from the surgical feeding tube site was present and empirical treatment with meropenem 1 g/8 h was initiated (Fig. 1).
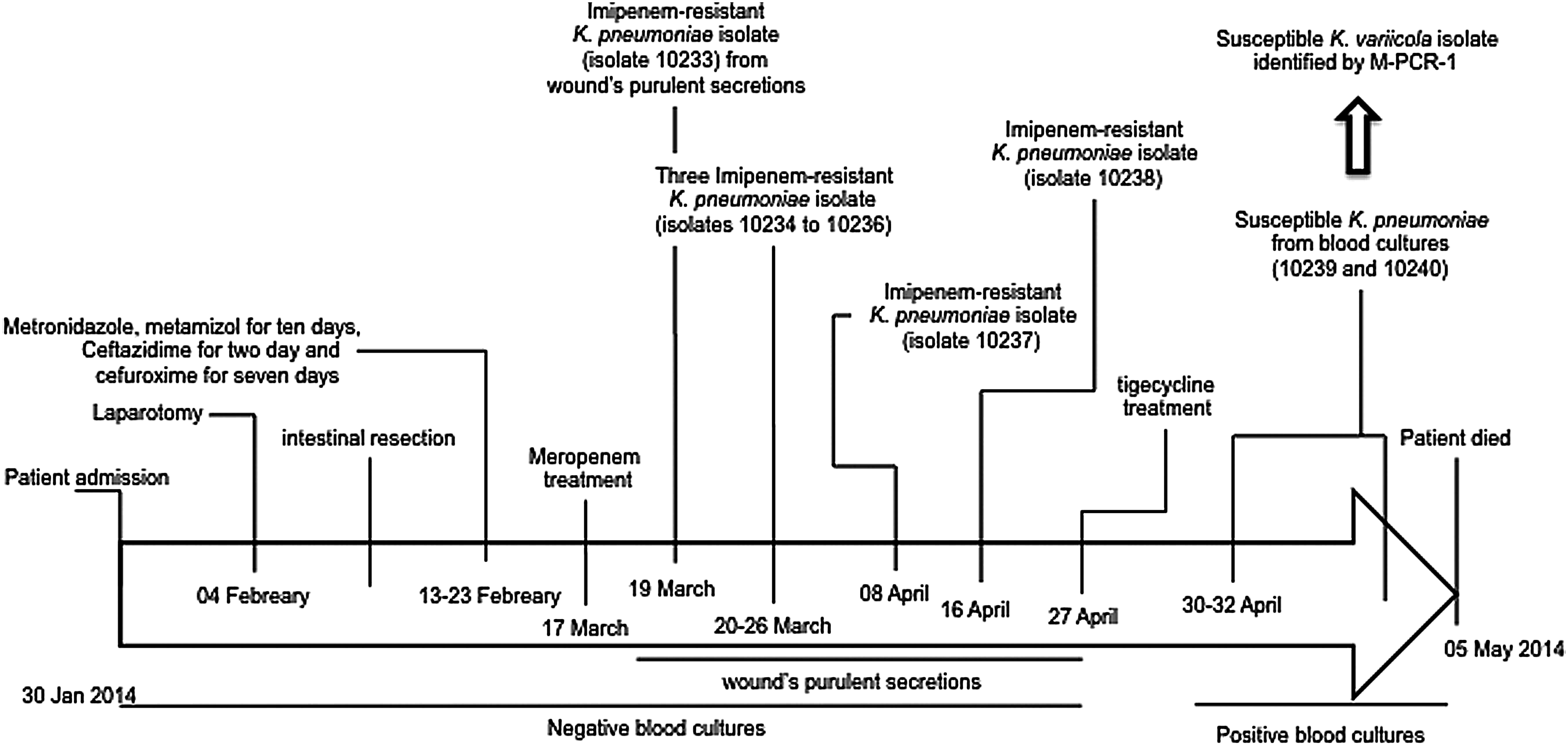
Clinical history, antimicrobial treatment, and isolates obtained during the hospitalization period.
Two days later, the first imipenem-resistant K. pneumoniae isolate (10233) was retrieved from the wound's purulent secretions (Fig. 1). After 6 days, three positive imipenem-resistant K. pneumoniae cultures were obtained from the surgical wound (10234, 10235, and 10236 isolates, each one obtained from the other at 2-day intervals). After 62 days of hospitalization, the patient's condition had slightly improved, and she was afebrile; however, purulent discharges continued from the wound. Antimicrobial treatment was suspended at that time. Three days later, an imipenem-resistant K. pneumoniae isolate (10237) was obtained from the wound's purulent secretions, and the patient slowly developed afebrile respiratory distress. Later, the purulent secretion increased without fever. She was placed in fasting, and leakage from the jejunostomy catheter was observed. A sixth imipenem-resistant K. pneumoniae (isolate 10238) was isolated from the purulent secretion, and treatment with tigecycline 50 mg/12 h was initiated. During this entire time, negative blood cultures were observed (Fig. 1).
Seventy-three days following admission into intensive care, fever, malaise and severe respiratory distress continued with additional decreased renal function. Five days later, she was prostrated with severe respiratory distress, lung edema, and abundant purulent exudate from the surgical site. Treatment with tigecycline was continued and positive susceptible K. pneumoniae blood cultures (isolate 10239) were obtained (Fig. 1). Two days later, on the 80th day of hospitalization, the patient was in a critical condition, with severe respiratory distress, fever, and severe renal failure, and purulent exudate continued to drain from the surgical wound. Another positive susceptible K. pneumoniae blood culture (isolate 10240) was obtained (Fig. 1). At this point, the tigecycline treatment was completed due to identification of imipenem susceptibility of the last two isolates (10239 and 10240). Unfortunately, the patient died 2 days later due to respiratory and renal complications.
Characterization of K. pneumoniae and K. variicola isolates
The eight isolates obtained from the same patient were identified as K. pneumoniae using a MicroScan Walkaway system. When the isolates were assayed to differentiate K. variicola from K. pneumoniae isolates using the multiplex-PCR, the results indicated that the first six imipenem-resistant isolates corresponded to K. pneumoniae (10233–10238), whereas the last two susceptible isolates corresponded to K. variicola (10239 and 10240) (Fig. 1 and Table 1). This result was confirmed by rpoB gene analysis (data not shown). All K. pneumoniae and K. variicola isolates were susceptible to tigecycline and colistin. The six imipenem-resistant K. pneumoniae isolates were resistant to ceftazidime, cefotaxime, piperacillin, ciprofloxacin, gentamicin, imipenem, and meropenem. Susceptibility (except to ampicillin) was confirmed for the last two K. variicola isolates (10239 and 10240) (Table 1).
The bacterial species identified by MicroScan Walkaway system.
The bacterial identification by M-PCR-1 and confirmed by rpoB analysis. 15
Identification of ESBL and carbapenemase activity identified by double-disc synergy 7 and biochemical test, 9 respectively.
The underline and boldface plasmids contain the KPC-3 and SHV-12 genes, respectively, identified by Southern hybridization.
MIC to tigecycline was determined without tween 80 according to CLSI. 7 The E. coli ATCC 25922 was used as control strain.
CAZ, ceftazidime; CIP, ciprofloxacin; CTX, cefotaxime; COL, colistin; ESBL, extended-spectrum β-lactamase; GEN, gentamicin; IMP, imipenem; MER, meropenem; MLST, multilocus sequence typing; MIC, minimal inhibitory concentrations; NA, not applied; PCR, polymerase chain reaction; PIP, piperacillin; PFGE, pulsed-field gel electrophoresis; SW, surgical wound; TGC, tigecycline; ND, not determined; +, positive; −, negative.
The PFGE analysis identified two clusters, A and B, where A corresponded to the six imipenem-resistant K. pneumoniae isolates and clone B included the two susceptible K. variicola isolates (Table 1). The nucleotide analyses of the ESBL and carbapenemase genes identified the ESBL SHV-12 and carbapenemase KPC-3 genes in the six imipenem-resistant K. pneumoniae isolates (clone A). K. pneumoniae MLST analysis identified the sequence type 258 (ST258) for the SHV-12-KPC-3-producing K. pneumoniae isolates (10233 and 10235) (Table 1). This ST258 was recently described in a high-mortality KPC-3 K. pneumoniae outbreak at the same hospital in Mexico City. 22 The mortality attributed to KPC-producing K. pneumoniae cultures isolated from respiratory tract infections and mechanical ventilation is a well-recognized risk factor. 10
Recently, several K. variicola isolates obtained from nosocomial infections in Mexico were identified. The isolates showed two antimicrobial susceptibility phenotypes: susceptibility to most of antimicrobials (except amoxicillin) and multiresistance (positive to ESBL production). 15 In addition, this susceptibility phenotype has also been identified in K. variicola BZ19 and 8917 isolates that were obtained from the feces of a hospitalized patient in Rio de Janeiro, Brazil, in 2005 and from a patient's sputum in Zapata Morelos, Mexico, in 2011, respectively.2,14 K. variicola is a recently described species that is closely related to K. pneumoniae, and its misclassification among K. pneumoniae isolates has been described; K. pneumoniae 342 and K. pneumoniae KP5-1 both correspond to K. variicola environmental isolates.15,23 The biochemical identification of K. pneumoniae by commercial kits does not include K. variicola, and molecular identification of K. variicola by phylogenetic analysis of the rpoB gene and/or multiplex-PCR is necessary. 15 Using the first option, K. variicola has been identified with variable prevalence (4.4–24.4%) in clinical, veterinarian, and ambient settings.2,4,18,21,23,26,30 Using the second method, it has been possible to identify K. variicola in several Mexican hospitals with susceptible, multiresistant, and hypermucoviscous phenotypes.14,15 However, virulence-determinant genes have been analyzed in few isolates of K. variicola. The genomes of the K. variicola BZ19 and 8917 clinical isolates were sequenced, and in silico analysis showed that they shared some virulence-associated determinants with K. pneumoniae. In the K. variicola BZ19, ureD, mrkD, uge, ureA, and wabG genes were identified and of these, the last four genes in addition to entB, iroN, iutA, kfuABC, and mceG genes were identified in the K. variicola 8917 genome. However, both genomes were negative for the allS gene.2,14 In the present study, 16 virulence-associated determinants were screened by PCR in both K. pneumoniae and K. variicola isolates. In K. pneumoniae, the isolates were positive for entB (enterobactin), fimA, fimH, ecpRAB, mrkD, uge, ureA, and wabG genes and three isolates were positive for iroB gene (Table 1). In K. variicola, the two isolates were positive for entB, mrkD, uge, ureA, and wabG genes (Table 1). K. pneumoniae isolates shared 5 of 16 virulence-associated genes with K. variicola. The main difference between this bacterial species was observed in fimA, fimH, and ecpRAB fimbrial-encoding genes identified only in K. pneumoniae isolates (Table 1). In the case of the entB gene, present in both K. pneumoniae and K. variicola, it encodes the enterobactin synthase component B. A siderophore that has been described as a growth and/or survival enhancer in K. pneumoniae, this gene has been specifically described in hypervirulent K. pneumoniae. 25 Nevertheless, the K. pneumoniae and K. variicola isolates in this study were negative for hypermucoviscous phenotype using the string test.
K. pneumoniae and K. variicola showed different plasmid profiles (Table 1) and the mating experiment assayed on K. pneumoniae isolates was unsuccessful. This situation eliminated the possibility of horizontal transfer among the K. pneumoniae and K. variicola isolates analyzed in this study. Nevertheless, these results do not remove the possibility that K. variicola may acquire plasmids from other bacterial species similar to K. pneumoniae, as was described in the K. variicola BZ19 isolate that contains the pBz19 plasmid. 2 Southern hybridization experiments showed on imipenem-resistance K. pneumoniae isolates, the presence of KPC-3 and SHV-12 genes on either the 100- or 70-kb and the 40-kb plasmids, respectively (Table 1). These results highlight the possibility of horizontal transfer of plasmids that encode cephalosporinase and carbapenemase genes in independent events in K. pneumoniae. Likewise, this could be occurring in K. variicola isolates, which have a similar plasmid pattern, but in this case occurs without plasmid-borne β-lactamase genes (Table 1). Furthermore, we know that the amoxicillin resistance is due to chromosomal-LEN β-lactamase described in K. variicola isolates. 15
In clinical practice, K. variicola isolates are initially identified as K. pneumoniae. However, this study for the first time accurately distinguished between K. pneumoniae and its bacterial sister K. variicola in the same patient. Likewise, the molecular characterization of K. pneumoniae isolates identified for the first time the presence of SHV-12-KPC-3-producing K. pneumoniae isolates in Mexico. This bacterium would have contributed to a prolonged hospitalization time and resulted in a change to the selected treatment (meropenem and tigecycline) (Fig. 1). A subsequent nosocomial infection with susceptible K. variicola was identified in the same patient and was obtained as a bloodstream infection when the patient was in a critical condition (severe respiratory distress, fever, and severe renal failure, in addition to continued drainage of a purulent exudate from the surgical wound). We considered that due to the critical condition of the patient when the susceptible K. variicola sample was acquired, it was not involved in the respiratory and renal complications. However, the recent description of a higher mortality rate for patients infected with K. variicola in comparison with K. pneumoniae could be considered, 18 highlighting the importance of differentiating the two bacterial species with very similar biochemical profiles. Consequently, the implementation of multiplex-PCR may be a useful tool in clinical laboratories.
In conclusion, the identification of K. variicola by biochemical techniques alone is not possible and may be enhanced by phylogenetic analysis of rpoB and/or multiplex-PCR. K. variicola could be considered an emerging bacterial species, with an increasing prevalence and mortality rate in comparison with other Klebsiella spp., such as Klebsiella quasipneumoniae. 6 The present report urges clinicians to identify K. variicola among K. pneumoniae infections and be aware of the potential of K. variicola involvement as a nosocomial infection, which may have been previously underestimated due to its susceptible phenotype.
Footnotes
Acknowledgment
We thank T. Rojas-Moreno for her excellent technical assistance.
Funding
This work was supported by grant CB-2009-01-130224 from CONACyT (Mexican Council for Science and Technology).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.