
Abstract
Introduction
H
Critical amino acid substitutions in PBP3 of beta-lactamase-negative ampicillin-resistant (BLNAR) isolates of H. influenzae are usually categorized according to Ubukata et al., where the N526K substitution defines group II, and isolates encoding the additional substitutions S385T and L389F are categorized as group III. 25 Other PBP3 substitutions also contribute to resistance, as has been unambiguously shown by site-directed mutagenesis. 9 Thus, clinical isolates of the same PBP3 group can exhibit different levels of resistance, although isolates of PBP3 group II are typically resistant to AMP, while isolates of PBP3 group III are typically also resistant to third-generation cephalosporins, according to EUCAST interpretative criteria. 3
Antimicrobial resistance has been less studied in the related species, Haemophilus parainfluenzae. In this species, four different TEM beta-lactamases have been documented, TEM-1, TEM-15, TEM-34, and TEM-182, of which TEM-15 and TEM-34 are encoded on small plasmids.4,24 TEM-15 has been expressed in laboratory strains of H. influenzae and H. parainfluenzae and was shown to hydrolyze third-generation cephalosporins. 24 Various PBP3 substitutions have been described in resistant strains of H. parainfluenzae, including N526K, N526H, N526S, and S385T.4,21,24 However, many of these strains also carry other resistance determinants, and no studies have hitherto documented the significance of PBP3 alterations in H. parainfluenzae by horizontal transfer of the mutated PBP3 gene to a susceptible host.
In this study, we characterize the relative contribution of PBP3 alterations and TEM-1, TEM-15, or ROB-1 beta-lactamases on cefotaxime (CTX) resistance using an isogenic environment of H. influenzae and H. parainfluenzae. Clinical isolates of the two species served as donors for mutated PBP3 genes and/or small plasmids bearing beta-lactamases. The specific enzymatic activity (SEA) of the beta-lactamases were assessed by hydrolysis of nitrocefin.
Materials and Methods
Bacterial strains and culture conditions
The wild-type strains H. influenzae strain Rd (ATCC 51907) and H. parainfluenzae strain HK 23 (CCUG 49489) were used as recipients in transformation experiments. H. parainfluenzae strain HK 23 is a naturally competent clinical isolate characterized elsewhere.5,6 A small plasmid encoding TEM-1 (pA1606) was isolated from a strain of H. influenzae, and a small plasmid encoding TEM-15 (pSF3) was isolated from a strain of H. parainfluenzae.18,24 Plasmid pB1000 encoding ROB-1, which has been isolated from several Pasteurellaceae species including H. influenzae,12–14,23 was kindly donated by Professor Bruno Gonzales-Zorn (Universidad Complutense de Madrid, Spain). The H. influenzae ftsI donor strain ATCC 49247 is a BLNAR H. influenzae quality control strain used for susceptibility testing. 1 Based on PBP3 amino acid substitutions, strain ATCC 49247 is categorized as a group II (encoding N526K) BLNAR strain according to Ubukata et al. 25 The H. influenzae ftsI donor strain UTAS252 encodes S385T and L389F in addition to N526K, and is categorized as a group III BLNAR strain. 19 No classification of PBP3 substitutions in H. parainfluenzae has been defined, therefore the suffix “-like” is used throughout with reference to the positions, but not necessarily the substitutions, in H. influenzae. The H. parainfluenzae ftsI donor strain ATCC 29242 is a group II-like strain (encoding N526K). The H. parainfluenzae ftsI donor strain SF3 is a group III-like strain (encoding N526H and S385T). 24 All PBP3 substitutions in the transpeptidase region of ftsI of H. influenzae donor strains relative to H. influenzae strain Rd, and in H. parainfluenzae donor strains relative to H. parainfluenzae strain HK 23 are given in Tables 1 and 2.
ftsI mutation classification according to Ubukata et al. 25
Amino acid positions of substitutions in the transpeptidase region (aa 265–580 of PBP3) relative to H. influenzae strain Rd. The critical mutations for group classification are in bold.
CTX, cefotaxime; MIC, minimum inhibitory concentration; PBP3, penicillin-binding protein 3.
ftsI mutation classification according to Ubukata et al. 25 for H. influenzae.
Amino acid positions of substitutions in the transpeptidase region (aa 265–580 of PBP3) relative to H. parainfluenzae strain HK 23. The critical mutations for group classification are in bold.
SF3 harbors a plasmid-encoded TEM-15, a chromosomally encoded TEM-1, and group III-like PBP3 substitutions.
H. influenzae and H. parainfluenzae were routinely cultured in Brain Heart infusion broth (Oxoid), supplemented with XV supplement (HTM supplement; Oxoid) (sBHI), and on chocolate agar plates (Statens Serum Institut, Denmark). Selective chocolate agar used for isolation of transformants harboring ftsI mutations or plasmids were produced from BHI agar (Oxoid) supplemented with 5% lysed defibrinated horse blood and 1 mg/L (ftsI) or 4 mg/L (plasmids) AMP. Inoculated chocolate agar plates were incubated at 37°C in an atmosphere containing 5% CO2. Broth cultures were incubated at 37°C with continuous rotation (130 rpm).
CTX susceptibility testing was performed using Etests® (Biomérieux) on Mueller-Hinton agar supplemented with 5% defibrinated horse blood and XV-supplements (MH-F; Oxoid) according to EUCAST recommendations. H. influenzae antimicrobial resistance control strain ATCC 49247 was routinely included to assess the accuracy of the Etest results.
Transformation
Electro-competent cells were prepared from H. influenzae strain Rd and H. parainfluenzae strain HK 23 as previously described. 25 A 2718 nt PCR product consisting of the entire ftsI open reading frame (ORF) (1833 nt) and flanking regions (316 nt upstream and 569 nt downstream) was amplified from the H. influenzae ftsI donor strains ATCC 49247 and UTAS252, as previously described. 20 A 2255 nt PCR product consisting of the entire ftsI ORF (1821 nt) and flanking regions (226 nt upstream and 207 nt downstream) was amplified from the H. parainfluenzae ftsI donor strains ATCC 29247 and SF3, as previously described. 24 Plasmid DNA was extracted using a QIAprep Spin Miniprep Kit (Qiagen) according to the manufacturer's instructions. Introduction of DNA into recipient strains H. influenzae Rd and H. parainfluenzae HK 23 was performed by electroporation, as previously described. 25 The transformation mixture was plated on selective agar plates and subcultured twice before assessment of successful transformation by the nitrocefin assay (plasmids) or by sequencing (ftsI transformants). Primers used for ftsI amplification and sequencing are listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr).
Nitrocefin hydrolysis assay
The SEA of plasmid-harboring transformants was determined by nitrocefin hydrolysis measurements. Strains were grown in 30 ml sBHI to an OD600nm of 0.2–0.3 at 37°C with continuous rotation (130 rpm). The cells were pelleted by centrifugation at 4,200 g for 15 min at 4°C. The pellet was resuspended in cold phosphate buffered saline (PBS) buffer, washed, and resuspended in cold PBS to obtain a ×10 concentration. The suspension was ultrasonicated on ice using a Branson Digital Sonifier in a setting consisting of eight bursts of 30 sec at 16% of full amplitude. The cellular debris was pelleted by centrifugation for 15 min at 12,000 g and 4°C, and the protein concentration of the supernatant (crude extract) was measured using the Quick Start Bradford Protein Assay Kit (BioRad) and bovine serum albumin as standard. The supernatant was stored at −20°C until use. Beta-lactamase activity was measured in triplicates from two separate preparations. Nine hundred microliters of 10 μM nitrocefin (Oxoid) and 100 μl crude extract was mixed, and the OD482nm was measured every 60 sec for 10 min at ambient temperature using a NanoPhotometer (IMPLEN). The sample was diluted to give a constant nitrocefin hydrolysis for at least 4 min. The SEA was calculated as the amount (μmol) of nitrocefin hydrolysed/min/mg of protein using the average change in absorbance at 482 nm/min during the linear phase of the reaction and the molar extinction coefficient of nitrocefin (14,000 M−1 cm−1).
Results
An isogenic environment was used to study the relative contribution of PBP3 substitutions and TEM-1, TEM-15, or ROB-1 beta-lactamases to CTX resistance in H. influenzae and H. parainfluenzae. We used PCR-amplified ftsI genes from two H. influenzae ftsI donors to transform the susceptible H. influenzae strain Rd, and PCR-amplified ftsI genes from two H. parainfluenzae ftsI donors to transform the susceptible H. parainfluenzae strain HK 23. The donors encoded substitutions near one (group II or group II-like) or two (group III or group III-like) of the highly conserved amino acid motifs surrounding the active site in PBP3. PCR-amplified ftsI genes were able to transform recipient strains of the same species, while the small plasmids isolated from H. influenzae (encoding TEM-1 or ROB-1) and H. parainfluenzae (encoding TEM-15) were able to transform recipient strains of both species.
CTX minimum inhibitory concentrations (MICs) of recipients and transformants are shown in Table 3. For H. influenzae strain Rd, transformation with ftsI from a PBP3 group II or group III donor increased the CTX MIC 6 or 60-fold, respectively, as previously shown.9,25 Presence of a TEM-1 or ROB-1 beta-lactamase did not significantly affect the susceptibility to CTX, while expression of TEM-15 increased the CTX MIC 19-fold. Thus, both expression of a group III PBP3 or expression of a TEM-15 beta-lactamase conferred resistance on H. influenzae strain Rd according to EUCAST interpretative criteria (MIC >0.125 mg/L) (Table 3).
Categorization of PBP3 substitutions are according to Ubukata et al. 25 MIC values in bold are categorized as resistant according to EUCAST interpretative criteria for H. influenzae.
ftsI from H. influenzae strain ATCC 49247.
ftsI from H. influenzae strain UTAS252.
ftsI from H. parainfluenzae strain ATCC 29242.
ftsI from H. parainfluenzae strain SF3.
GM, geometric mean of three independent measurements.
For H. parainfluenzae strain HK 23, transformation with ftsI from a PBP3 group II-like or group III-like donor increased the MIC six or ninefold, respectively, but did not confer resistance according to EUCAST interpretative criteria for H. influenzae. Presence of a TEM-1 or ROB-1 beta-lactamase did not significantly affect the susceptibility to CTX, while expression of TEM-15 increased the CTX MIC 29-fold. Presence of TEM-15 on a background of native PBP3 conferred resistance on H. parainfluenzae strain HK 23 according to EUCAST interpretative criteria for H. influenzae. The CTX MIC was further increased by the presence of PBP3 substitutions (Table 3).
We assessed the enzymatic activity of TEM-1, TEM-15, and ROB-1 beta-lactamases in H. influenzae and H. parainfluenzae by quantitating the hydrolysis of nitrocefin. The SEA of TEM-1 was three to fivefold higher than that of TEM-15 (Fig. 1).
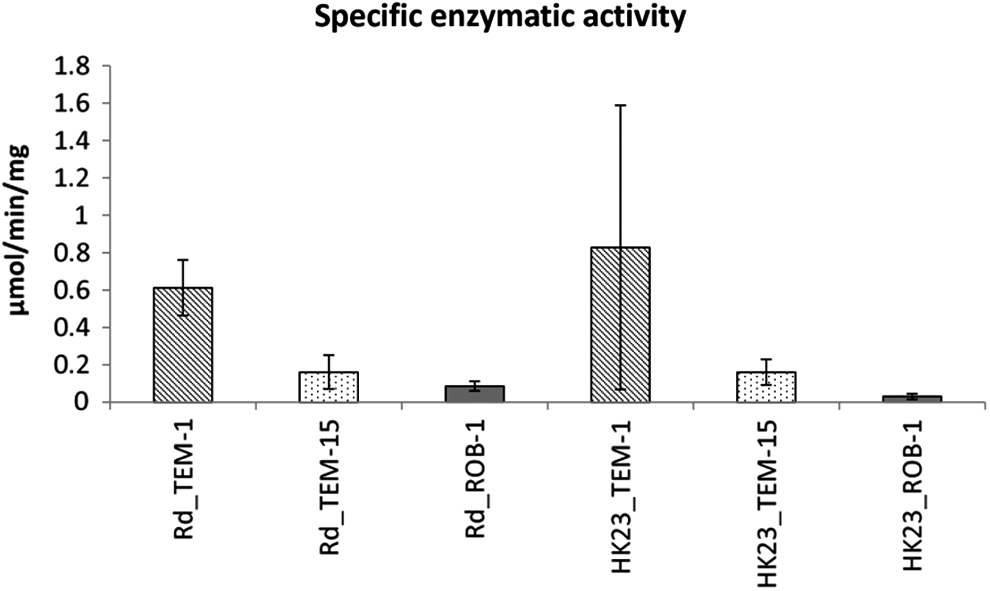
SEA of TEM-1, TEM-15, and ROB-1 in isogenic transformants of Haemophilus influenzae strain Rd and Haemophilus parainfluenzae strain HK 23 measured by nitrocefin hydrolysis per mg membrane protein. SEA of TEM-1-expressing transformants are shown in cross line bars; SEA of TEM-15-expressing transformants are shown in dotted bars; and SEA of ROB-1-expressing transformants are shown in solid bars. Each bar represents the average of six individual transformants. The high standard deviation of HK 23_TEM-1 is caused by one sample with very high SEA. SEA, specific enzymatic activity.
Discussion
Transformation of susceptible strains with PCR-amplified ftsI genes have been crucial for identification and assessment of significance of PBP3 substitutions in H. influenzae.9,19,25 Here, we report the first successful transformation of susceptible strains of H. parainfluenzae with PCR-amplified ftsI genes from resistant isolates. The mechanism responsible for the BLNAR phenotype of H. parainfluenzae can thus be addressed experimentally. We were unable to perform inter-species transformation with ftsI, probably due to sequence divergence or other species barriers. Thus, we were limited to intra-species transfer of the ftsI gene, and only two reference strains of H. parainfluenzae with reduced susceptibility to beta-lactams were available for this study.1,24 In H. influenzae, the predominant substitution in PBP3 at position 526 is replacement of asparagine with lysine (N526K), with only a single report of N526H. 16 For H. parainfluenzae, three substitutions at this position have been reported (N526K, N526H, and N526S) among the relatively few BLNAR strains that have been characterized.4,21,24 In addition, substitutions at two positions near the highly conserved amino acid motif S379-S-N, namely S385T and L389F, have been observed for H. influenzae, whereas only the former substitution have so far been reported for H. parainfluenzae. While the group II substitutions of PBP3 conferred similar increases in CTX MIC in both species (Table 3), the group III-like substitutions only marginally increased the CTX MIC over the group II-like substitutions in H. parainfluenzae. Whether N526H confer less resistance than N526K, or whether S385T is only of marginal importance in H. parainfluenzae, are not clear at present. However, the significance of various substitutions in PBP3 in H. parainfluenzae can now be experimentally addressed.
The TEM-1 and ROB-1 beta-lactamases are common in clinical strains of H. influenzae. These enzymes do not significantly affect the susceptibility to CTX in this species or in H. parainfluenzae. ROB-1 is capable of hydrolyzing the first generation cephalosporin cefachlor at an appreciable rate, although significant increases in cefachlor MICs appear related to inoculum effects of the microbroth method or simultaneous presence of PBP3 substitutions. 23 Extended-spectrum beta-lactamases conferring CTX resistance have never been documented in naturally occurring strains of H. influenzae, although TEM-3 and TEM-4 have been expressed in strain Rd in the laboratory and shown to confer CTX resistance. 2 Plasmid pSF3 bearing TEM-15 originated from a strain of H. parainfluenzae. 24 The TEM-15 beta-lactamase conferred CTX resistance on both species using EUCAST interpretative criteria for H. influenzae, as previously shown. 24 We assessed the SEA of TEM-1, TEM-15, and ROB-1 in both H. influenzae and in H. parainfluenzae. The TEM-1 nitrocefin hydrolyzing rate surpassed that of TEM-15, suggesting that the increase of hydrolytic spectrum of the latter enzyme comes at the cost of efficiency. An increased transcription of beta-lactamase, which has recently been shown for plasmid pSF3, 17 may compensate for the decrease of efficiency. The competitive disadvantage of pSF3 on H. parainfluenzae observed in that study may partly explain why TEM-15 has only sporadically been reported in this species; however, it is a matter of concern that the fitness cost of this replicon is lower in the more pathogenic H. influenzae than in its theoretical native host. 17 In addition, further studies are needed to clarify strain-to-strain variability of the capacity to retain beta-lactamase-bearing plasmids.
In conclusion, TEM-15 but not TEM-1 or ROB-1 beta-lactamases confer resistance to CTX on both H. influenzae and H. parainfluenzae. Similar to H. influenzae, PBP3 substitutions affect the susceptibility to third generation cephalosporins in H. parainfluenzae, although the N526H in combination with S385T are insufficient to transgress the EUCAST interpretative criteria of CTX resistance in this species. Site-directed mutagenesis of ftsI and gene recombinants 9 are needed to elucidate the BLNAR phenotype of H. parainfluenzae.
Footnotes
Acknowledgments
Tanja Sørensen is acknowledged for technical assistance with the nitrocefin assay and susceptibility testing. We are grateful to Dr. Stephen Tristram (University of Tasmania, Australia) and to Professor Bruno Gonzales-Zorn (Universidad Complutense de Madrid, Madrid, Spain) for the generous gift of H. parainfluenzae strain SF3 and plasmid pB1000, respectively.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.