
Abstract
Emergence and spread of antibiotic-resistant Acinetobacter baumannii have become a major public health concern. This study was designed to investigate the efficacy of Holarrhena antidysenterica extract and its major steroidal alkaloid conessine as resistance-modifying agents (RMAs) on the susceptibility of A. baumannii to novobiocin and rifampicin. A significant synergistic activity of both the extract and conessine in combination with either novobiocin or rifampicin with fractional inhibitory concentration index ≤0.5 was demonstrated. Fluorescent dyes and different efflux pump inhibitors were used to further investigate the synergism. Increase in the uptake of 1-N-phenylnaphthylamine in the bacterial cells treated with the extract and conessine was not observed indicating that both substances did not act as permeabilizers. With regard to efflux pump inhibition, no accumulation in ethidium bromide (EtBr) was noticed suggesting that the AdeABC pump was not involved. In contrast, accumulation in Pyronin Y was significantly increased (p < 0.05) demonstrating that the synergism was due to interference with the AdeIJK pump. Study on frequencies of the spontaneous mutational resistance to the extract in combination with antibiotics demonstrated attenuation in drug-resistant organisms. Thus, H. antidysenterica extract and conessine as RMAs may offer a combinatory therapy to restore antibiotic susceptibility in the extensively drug-resistant A. baumannii.
Introduction
A
Carbapenems remain the antibiotics of choice for the treatment of A. baumannii infections. However, carbapenem-resistant A. baumannii is now increasingly reported worldwide. 19 Colistin, an old antibiotic from the polymyxin group, has become the last resort for treatment of MDR A. baumannii. In addition, colistin demonstrates adverse effects regarding nephrotoxicity and neurotoxicity. Recently, colistin-resistant A. baumannii has already been reported. Therefore, combination therapy of colistin with other antibiotics might be an applicable alternative. Many in vitro, in vivo, and clinical studies have shown synergistic effects of colistin in combination with rifampicin for MDR or extensively drug-resistant (XDR) A. baumannii. 4 In addition, a recent study confirmed the synergistic colistin/rifampicin combination against heteroresistant isolates and prevented the development of colistin-resistant mutants. 32 However, moderate hepatic cytolysis has been noticed in clinical study. 25
The search for new compounds with biological properties is viewed as an important path toward fighting and reducing antibiotic resistance. 1 Holarrhena antidysenterica (L.) Wall., belonging to the Apocynaceae family, is an important medicinal plant used in Ayurvedic, Unani, and Thai traditional medicines for the treatment of amoebic dysentery and diarrhea. H. antidysenterica barks are composed of alkaloids, particularly steroidal alkaloid conessine that is the principle compound used as a therapeutic drug.3,21 Interestingly, the extract demonstrated a resistance-modifying ability to enhance the novobiocin activity by an increase in the 1-N-phenylnaphthylamine (NPN) uptake on outer membrane permeability of A. baumannii isolates that acted as a permeabilizer.6,30 In addition, our previous study showed that H. antidysenterica extract displayed synergistic effect with rifampicin against clinical A. baumannii isolates. 7
To search for plant natural products which can restore extensive drug resistance in A. baumannii, this study was designed to investigate the efficacy of H. antidysenterica extract and its steroidal alkaloid, conessine as resistance-modifying agents (RMAs) in combination with rifampicin and novobiocin, current recommended antibiotics for the treatment of A. baumannii infections.
Materials and Methods
Preparation of plant extract and conessine, antibiotics, and reagents
H. antidysenterica barks (4 kg) were collected from the Queen Sirikit Botanic Garden, Chiang Mai, Thailand and kindly authenticated by a taxonomist, Dr. Katesarin Maneenoon. The fresh barks were washed with distilled water and dried at 60°C overnight. Finely powdered plant material was macerated with 95% ethanol (1: 2 w/v) for 7 days at room temperature. After filtration through Whatman No. 1 paper, the extract was evaporated using a rotatory evaporator, and kept at 55°C until it was completely dried. The extract (92 g) was stored at −20°C until used. Conessine was obtained from Santa Cruz Biotechnology. Antibiotics, including novobiocin and rifampicin, were from Sigma-Aldrich. Stock solutions of H. antidysenterica extract, conessine, and antibiotics were prepared in dimethyl sulfoxide (DMSO).
Carbonyl cyanide m-chlorophenyl hydrazone (CCCP), EtBr, NPN, and Pyronin Y were purchased from Sigma-Aldrich. Ethylenediaminetetraacetic acid (EDTA) was purchased from Fisher Scientific. Mueller Hinton Broth (MHB), Tryptic Soy Broth, and agar were obtained from Becton Dickinson Microbiology Systems.
Bacterial strains
Twenty-seven clinical A. baumannii Natural Product Research Center (NPRC) collection from pus (n = 1), tissue (n = 1), blood (n = 3), sputum (n = 9), body fluid (n = 5), urine (n = 7), and cerebrospinal fluid (n = 1) were originally obtained from infected patients at the Songklanagarind Hospital. The isolates were originally classified as non-MDR (n = 8), MDR (n = 4), and XDR (n = 15) according to the antimicrobial susceptibility profile. A. baumannii ATCC 19606 was used as a quality control.
Determination of minimum inhibitory concentration
Minimum inhibitory concentration (MIC) was determined by broth microdilution assay in accordance with the Clinical and Laboratory Standards Institute (CLSI) recommendation. Serial dilutions of H. antidysenterica extract, conessine, novobiocin, and rifampicin were prepared in MHB. To investigate the effect of each agent, 100 μL culture (1 × 106 cfu/ml) was mixed with 100 μL H. antidysenterica extract, conessine, or antibiotics. Synergistic effects of the extract or conessine and antibiotics were assessed by adding 100 μL bacterial culture into a well containing 50 μL H. antidysenterica extract or conessine and 50 μL novobiocin or rifampicin. The test was then read after 18 hr of incubation at 37°C. Each value was mean of triplicates from three independent experiments. MIC is defined as the lowest concentration that inhibited bacterial growth. Synergistic effect was classified by fractional inhibitory concentration index (FICI) parameter according to the following formula
13
:
The results were interpreted as follows: FICI ≤0.5 synergistic, 0.5< FICI <1 partially synergistic, FICI = 1 additive, 1< FICI ≤4 indifferent, and FICI >4 antagonistic.
The resistance-modifying ability of H. antidysenterica extract and conessine
The resistance-modifying ability of H. antidysenterica extract and conessine was determined by growth inhibition (GI) assay as previously described
6
and further confirmed by the time–kill assay. Briefly, 100 μl culture (1 × 106 cfu/ml) was inoculated into a 96-well sterile microtiter plate containing each of 50 μl of the extract (125 mg/L), conessine (2.5 mg/L), novobiocin, rifampicin (1/2 to 1/16MIC), and MHB. Combinations of 50 μl H. antidysenterica extract or conessine and 50 μl novobiocin or rifampicin were added with 100 μl bacterial culture. The antibacterial activity was calculated as percentage of GI after incubation at 37°C for 18 hr according to the following equation:
where ODcontrol is optical density (OD620nm) of the bacterial culture in MHB supplemented with 1% DMSO as positive control and ODtest is OD620nm of the bacterial culture in MHB supplemented with the tested agent.
Outer membrane permeabilization
The efficacy of H. antidysenterica extract or conessine to permeabilize bacterial membrane was determined using the NPN uptake assay. NPN is a membrane potential-sensitive fluorescent probe that shows increased fluorescence upon exposure to a hydrophobic environment. 16 In this study, A. baumannii (1 × 106 cfu/ml) was incubated with agitation at 37°C for 4–5 hr. The cells were harvested by centrifugation at 3,000 g for 15 min at room temperature, washed twice, and resuspended to an OD620 nm of 1.0 in 5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) buffer (pH 7.2). An aliquot of 100 μl of the bacterial suspension was added into a well in black microtiter plate containing each of 50 μl H. antidysenterica extract (7.81, 15.63, 31.25, 62.5, 125 mg/L), conessine (2.5, 5 mg/L), EDTA (50 μM) as a permeabilizer, followed by adding 50 μl of NPN (40 μM). NPN fluorescence intensity (excitation 322 nm, emission 424 nm) was monitored at 37°C after 10 min for 3 hr using a Varioskan Flash spectral scanning multimode reader (Thermo Fisher Scientific, Finland). Each value was mean of triplicates from three independent experiments. The comparisons of the concentrations accumulated in different experiments were analyzed by a two-tailed Student's t-test. A p < 0.05 was considered significant.
Inhibition of efflux pumps
Measurement of fluorescent dye accumulation, which is an efflux pump substrate, was performed to assess the role of efflux pumps in A. baumannii. Inhibition of A. baumannii efflux pumps was determined by measuring accumulation of EtBr, an indicator of the AdeABC efflux pump inhibition and Pyronin Y, an indicator of the AdeIJK efflux pump inhibition, 9 using fluorescent dye whole cell accumulation assay as previously described. 8 EtBr binds to double-stranded DNA resulting in a substantial increase in fluorescence, whereas Pyronin Y binds to RNA resulting in fluorescence quenching. 24 Briefly, A. baumannii (1 × 106 cfu/ml) was incubated with agitation at 37°C for 4–5 hr. The bacterial cells were harvested by centrifugation at 3,000 g for 15 min at room temperature, washed twice, and resuspended to an OD620nm of 1.0 in 5 mM HEPES buffer (pH 7.2). An aliquot of 100 μl of the cell suspension was added into a well in black microtiter plate containing each of 50 μl H. antidysenterica extract (7.81, 15.63, 31.25, 62.5, 125 mg/L), conessine (2.5, 5 mg/L), CCCP (50 μM) as a positive control, followed by adding 50 μl of EtBr or Pyronin Y (4 mg/L). Relative fluorescence intensity was recorded at 37°C after 10 min for 3 hr using a Varioskan Flash spectral scanning multimode reader. Excitation and emission wavelengths of EtBr and Pyronin Y were set at 518 and 605 nm and 545 and 570 nm, respectively. Each value was mean of triplicates from three independent experiments. The mean and standard deviations were determined by a two-tailed Student's t-test. A p < 0.05 was considered significant.
Determination of spontaneous mutation frequencies for resistance to novobiocin or rifampicin combined with H. antidysenterica extract
Reduction in spontaneous mutation of antibiotics resistance by H. antidysenterica extract in A. baumannii strains were estimated using mutation frequency as previously described.28,31 In this study, the bacterial strains were cultivated on TSA plates at 37°C overnight, transferred into MHB, and incubated without agitation at 37°C for 18 hr. The suspensions were adjusted to a 0.5 McFarland standard (1.5 × 108 CFU/ml) and diluted to achieve a final inoculum of approximately 1.5 × 104, 1.5 × 106, and 1.5 × 108 CFU/ml, respectively. An aliquot of 100 μl was inoculated on agar plates supplemented with H. antidysenterica extract (2MIC), novobiocin (5MIC and 10MIC), rifampicin (5MIC and 10MIC), H. antidysenterica extract in combination with either novobiocin or rifampicin. After 48 hr incubation at 37°C, colonies were enumerated. Mutation frequencies were expressed as the number of colonies/number of CFU inoculated. Bacterial isolates with spontaneous mutation rates over 10−7 were considered as mutator. The reduction rate of spontaneous mutation frequencies was defined as a 2-fold decrease in colony count after 48 hr with the combination compared to that with the most active single agent alone. Each value was mean of triplicates from three independent experiments.
Results
Determination of MIC
MICs of H. antidysenterica extract, conessine, novobiocin, and rifampicin against twenty-seven A. baumannii isolates were determined following CLSI recommendation. MICrange values of the extract, novobiocin, and rifampicin were 625 to >1,000, 1.56–25.0, and 0.31–5.0 mg/L, respectively. MIC90 values of novobiocin and rifampicin were 12.5 and 2.5 mg/L, respectively. Three representative A. baumannii isolates, including A. baumannii ATCC 19606, non-MDR A. baumannii NPRC AB029, and XDR A. baumannii NPRC AB010, were selected based on the antimicrobial susceptibility profile for further studies. An MIC value of conessine against three representative A. baumannii isolates was 10 mg/L.
Synergistic effects of H. antidysenterica extract or conessine in combination with novobiocin or rifampicin against A. baumannii were classified using the FICI parameter. The extract (Fig. 1A and B) and conessine (Fig. 1C and D) markedly lowered MICs of both antibiotics against all of the tested A. baumannii isolates (Table 1). The combined activity of interaction between H. antidysenterica extract and antibiotics against A. baumannii resulted in FICIs ranging from 0.13 to 0.16 in NPRC AB029 and 0.33 to 0.45 in NPRC AB010. The synergistic effect of interaction between conessine and antibiotics against A. baumannii resulted in FICIs ranging from 0.28 to 0.31 and 0.38 to 0.5 in the non-MDR and XDR isolates, respectively. All of the tested A. baumannii isolates showed 5- to 10-fold reduction in MIC values with the extract, 4-fold reduction in MIC values with conessine, and 4-fold reduction in MIC values with both antibiotics. The results revealed that H. antidysenterica extract and conessine in combination with antibiotics could effectively inhibit A. baumannii growth.

Growth inhibition of Acinetobacter baumannii isolates by Holarrhena antidysenterica extract (HA; 125 mg/L)
The resistance-modifying ability of H. antidysenterica extract and conessine was assayed using broth microdilution method.
Fractional inhibitory concentration index (FICI) as a measure of efficacy of combinatory therapy according to the following formula: FICI = (MIC of extract in combination/MIC of extract alone) + (MIC of antibiotics in combination/MIC of antibiotics alone).
FICI was interpreted as follows: FICI ≤ 0.5 synergistic, 0.5 < FICI < 1 partially synergistic, FICI = 1 additive, 1< FICI ≤ 4 indifferent, and FICI > 4 antagonistic.
With HA 125 mg/L, CON 2.5 mg/L.
CON, conessine; HA, H. antidysenterica; MDR, multidrug-resistant; NOV, novobiocin; XDR, extensively drug-resistant.
The resistance-modifying ability of H. antidysenterica extract and conessine on A. baumannii
The resistance-modifying ability was performed by the same principle as the antimicrobial activity assay. Figure 1 presented the resistance-modifying ability of H. antidysenterica extract (125 mg/L) or conessine (2.50 mg/L) combined with both novobiocin and rifampicin (1/4MIC) against A. baumannii isolates. The extract effectively displayed greater synergistic antibiotic activities with both novobiocin and rifampicin than conessine in all tested isolates.
Outer membrane permeabilization
Accumulation of NPN in A. baumannii isolates after treated with H. antidysenterica extract (125, 62.5, 31.25, 15.63, 7.81 mg/L), conessine (5, 2.5 mg/L), and a permeabilizer, EDTA (50 μM) was shown in Fig. 2. EDTA weakened the outer membrane of both A. baumannii isolates and a reference strain as clearly indicated by an increase in NPN accumulation whereas the extract and conessine produced no effect on the level of NPN accumulation in all A. baumannii isolates. The results implied that H. antidysenterica extract and conessine did not act as permeabilizers in A. baumannii isolates.
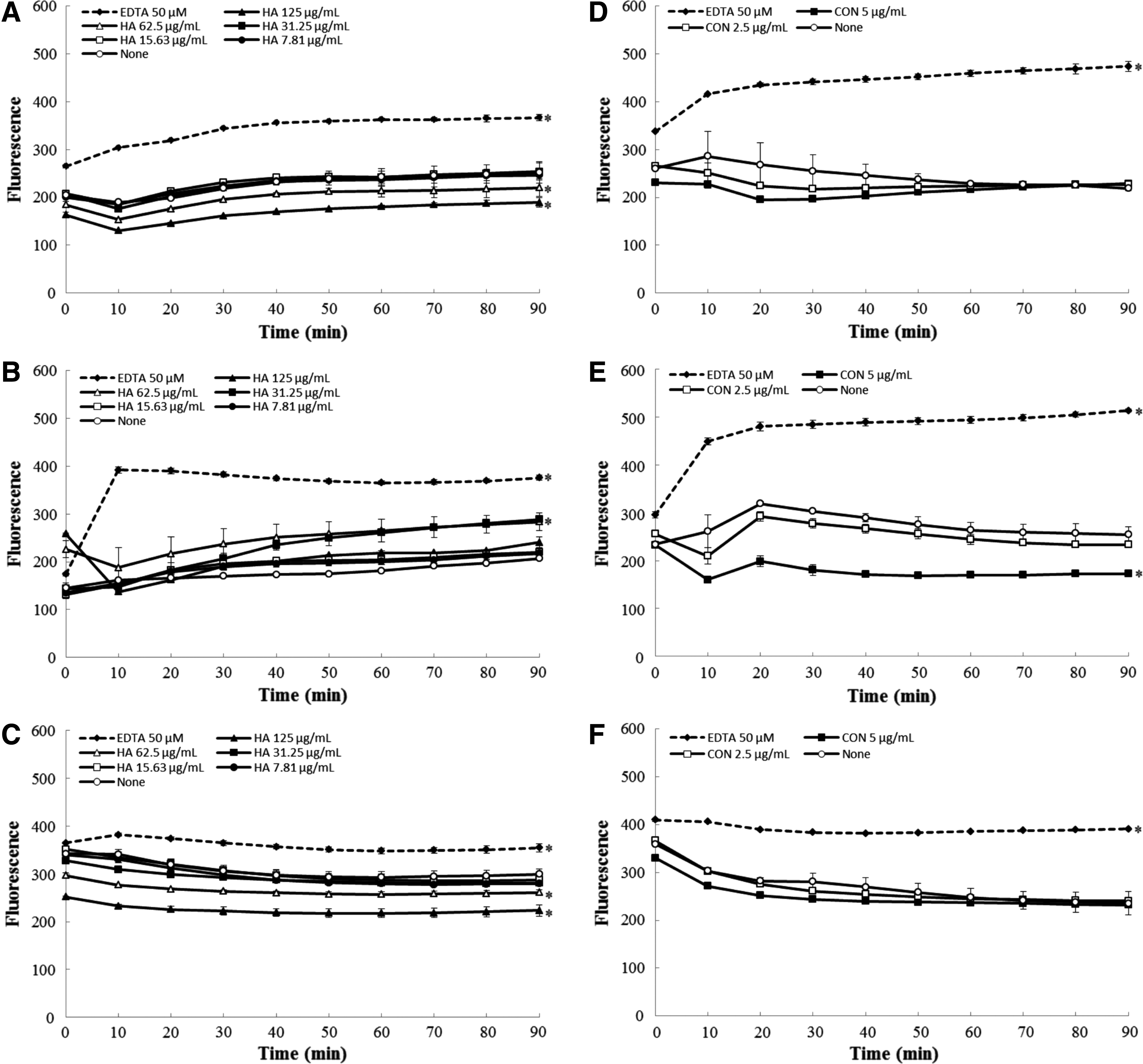
Effects of H. antidysenterica extract (HA) and conessine (CON) on 1-N-phenylnaphthylamine accumulation in A. baumannii ATCC 19606
Inhibition of efflux pumps
To elucidate a mechanism of the resistance-modifying ability of H. antidysenterica extract and conessine in A. baumannii isolates, we further evaluated their potential to increase the accumulation of efflux pump substrate EtBr, an indicator of AdeABC efflux inhibition, and Pyronin Y, an indicator of AdeIJK efflux inhibition. Accumulation of EtBr or Pyronin Y in A. baumannii isolates after treating with the extract (125, 62.5, 31.25, 15.63, 7.81 mg/L), conessine (5, 2.5 mg/L), and an efflux pump inhibitor (EPI), CCCP (50 μM) was demonstrated in Figs. 3 and 4, respectively. In the presence of H. antidysenterica extract and conessine at various concentrations, Pyronin Y accumulation was significantly increased (p < 0.05), whereas no increase in EtBr accumulation was noticed. The study indicated that H. antidysenterica extract and conessine interfere with AdeIJK pump in A. baumannii isolates.
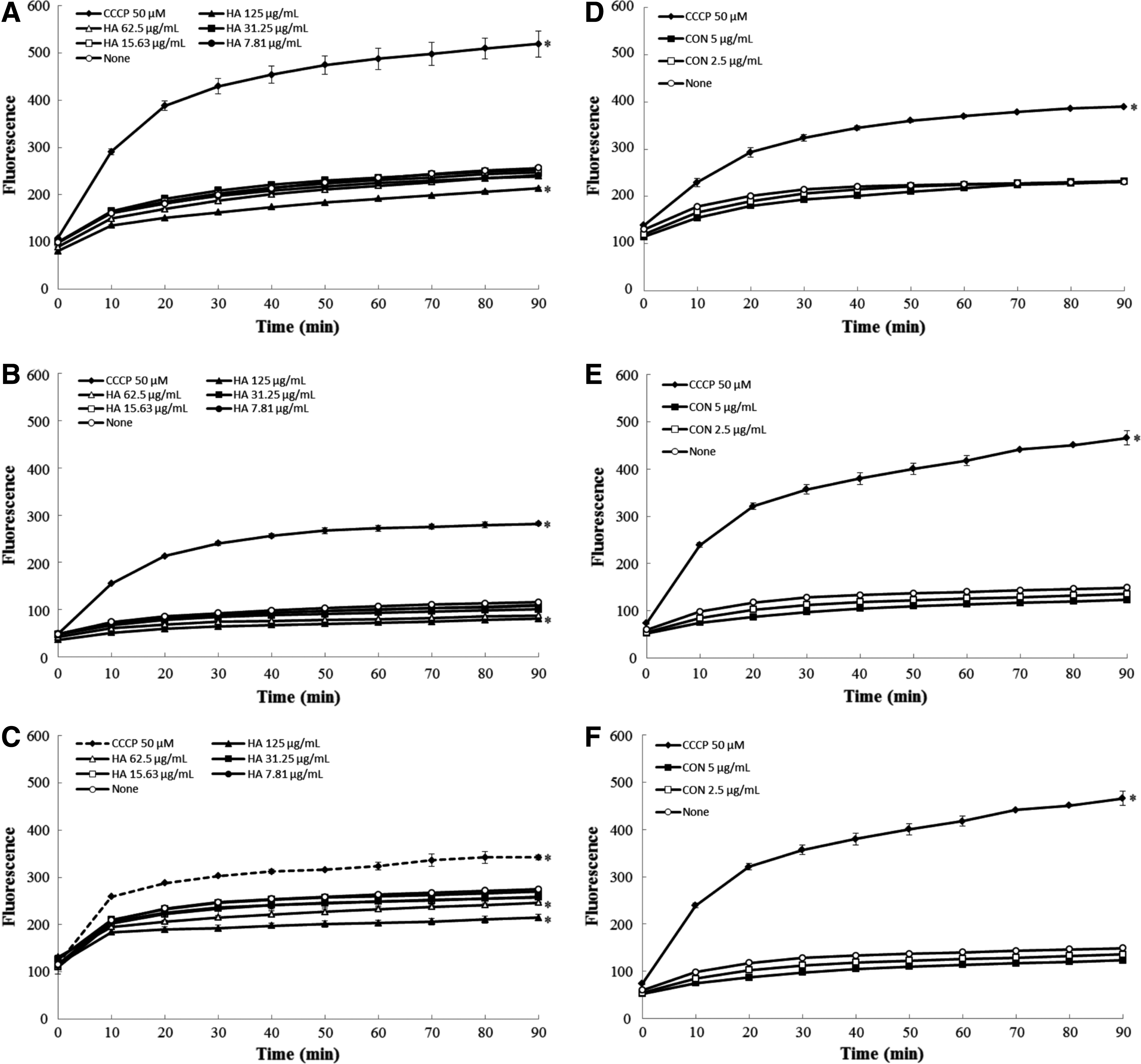
Effects of H. antidysenterica extract (HA) and conessine (CON) on ethidium bromide accumulation in A. baumannii ATCC 19606

Effects of H. antidysenterica extract (HA) and conessine (CON) on Pyronin Y accumulation in A. baumannii ATCC 19606
Frequencies of spontaneous mutational resistance to novobiocin or rifampicin combined with H. antidysenterica extract
Spontaneous mutation frequencies of H. antidysenterica extract (2MIC), novobiocin (5MIC and 10MIC), rifampicin (5MICand 10MIC), and the combinations of H. antidysenterica extract and novobiocin or rifampicin were determined in representative A. baumannii isolates (A. baumannii ATCC 19606, non-MDR A. baumannii NPRC AB029, and XDR A. baumannii NPRC AB010) (Table 2). Drug-resistant mutants were attenuated (mutation rate <10−8) after treatment in non-MDR A. baumannii NPRC AB029 and XDR A. baumannii NPRC AB010 with the combinations of the extract and novobiocin at 5MIC and 10MIC, respectively. Interestingly, no mutants were recovered in A. baumannii ATCC 19606 upon selection with the combination of H. antidysenterica extract and rifampicin (5MIC). However, such combination could not attenuate the mutation in non-MDR and XDR A. baumannii.
Reduction in spontaneous mutation frequencies of antibiotic resistance by H. antidysenterica extract in A. baumannii strains were estimated using mutation frequency.
Mutation frequencies were expressed as the number of resistant colonies/number of CFU inoculated. Mutation rates over 10−7 were considered as mutator. The reduction rate of spontaneous mutation frequencies was defined as showing a 2-fold decrease in colony count after 48 hr with the combination, compared with the most active single agent alone.
MIC, minimum inhibitory concentration.
Discussion
The spread of antibiotic-resistant bacteria has become a major public health concern. A. baumannii is one of the most life-threatening Gram-negative pathogens; MDR clones of A. baumannii are spreading worldwide, causing outbreaks in hospitals.10,17,29 Hence, there is an urgent need for discovery and development to provide novel antibacterial agents to confer antimicrobial susceptibility of the organisms. Plant natural products as RMAs are considered as one of the important strategies to restore the efficacy of antibiotics. 1 Some experimental evidences indicated that H. antidysenterica extract has a resistance-modifying ability against A. baumannii.6,7,30 However, the potential roles of the extract and its steroidal alkaloid, conessine as RMAs, in conferring modulation against XDR A. baumannii have not been well documented.
The expression of both intrinsic and acquired multidrug resistance in A. baumannii has been related to the synergistic interaction between low outer membrane permeability and constitutive expression of efflux pumps. 39 Very few compounds have been reported to affect membrane permeability against A. baumannii. Plant-derived compounds, 6-dehydrogingerdione, 10-gingerol, 6-gingerol, and 6-shogaol, isolated from Zingiber officinale, were displayed to modulate tetracycline resistance. 40 Geraniol derived from Helichrysum italicum increased the efficacy of β-lactams, quinolones, and chloramphenicol. 22 Coriander oil isolated from Coriandrum sativum improved the antibiotic effectiveness of ciprofloxacin, gentamicin, tetracycline, and chloramphenicol. 12
Efflux systems, which extrude antibiotics from the bacterial cell, have been related to play an important role in clinically relevant antibiotic resistance in MDR A. baumannii. To date, three different RND pumps, namely AdeABC, AdeIJK, and AdeFGH have been described in A. baumannii. The AdeABC system was demonstrated to efflux EtBr, whereas the AdeIJK pump effluxes novobiocin, rifampicin, and pyronin. 9 Inhibition of efflux systems is a strategy for potentiating antibiotic efficacy. To determine EPIs in antibiotic-resistant A. baumannii, measurement of the both EtBr and Pyronin Y accumulation was provided to assess the role of efflux pumps. EPIs have been shown to modulate antibiotic activity against A. baumannii such as CCCP, 1-(1-naphthylmethyl)-piperazine (NMP), and phenyl-arginine-β-naphthylamide (PAβN).1,9 We demonstrated that the extract and its steroidal alkaloid, conessine interfered the efflux system, in particular, the AdeIJK pump as evidenced by the increment of Pyronin Y accumulation. Moreover, interference of this pump by H. antidysenterica extract and conessine facilitated the higher intracellular accumulation of novobiocin and rifampicin, which are pump-specific substrates, resulting in the higher antibiotic susceptibility in combination therapy (Table 1). Conversely, previous study by Chusri et al., 6 suggested that H. antidysenterica extract acted as an outer membrane permeabilizer to enhance novobiocin activity after being pretreated for 18 hr. It is speculated that the membrane-permeabilizing activity of the extract was not an immediate effect as the NPN uptake was not seen during the first 3 hr of treatment.
A number of plant alkaloids have been documented to potentiate antibiotic activity against antibiotic-resistant strains. 35 Main mechanisms of action have been claimed to inhibit efflux pump systems in bacterial cells. Several plant alkaloids, such as 5′-methoxyhydnocarpin 36 from Berberis fremontii, piperine 20 from Piper nigrum and Piper longum, capsaicin 18 from genus Capsicum, and reserpine 33 from Rauwolfia vomitoria, were shown as EPIs to inhibit NorA efflux pump in Staphylococcus aureus. In other context, piperine remarkably increased the mupirocin activity and attenuated mutation frequency by acting as an inhibitor of MdeA efflux pump in the organisms, 23 whereas reserpine could restore the tetracycline activity by interfering with Tet (K) efflux pump in S. aureus 15 and Bmr efflux pump in Bacillus subtilis. 27 Curcumin derived from Curcuma longa could act as an EPI to enhance gentamicin and ciprofloxacin activity against Pseudomonas aeruginosa. 26 In this study, H. antidysenterica extract effectively showed a synergistic inhibitory activity with antibiotics in all tested A. baumannii isolates. Moreover, its synergistic effect was greater than the purified compound, conessine. Siddiquia et al. 34 reported that the % yield of conessine from H. antidysenterica bark extract was approximately 0.09%. In this study, H. antidysenterica extract at 125 mg/L contains approximately 4.5 mg/L conessine content, which was higher than the purified conessine used (2.5 mg/L). Previous studies documented that conessine was the major active compound in H. antidysenterica extract.5,11,37 Therefore, the synergistic inhibitory effect was thought to be contributed by conessine content rather than other components in the extract.
Combinations of RMAs and antibiotics are commonly reported for the treatment of MDR A. baumannii and hindering the development of resistance. This study clearly demonstrated that the combinations of H. antidysenterica extract and novobiocin could delay the development of antibiotic resistance. However, rifampicin resistance was detected in non-MDR and XDR A. baumannii. A previous report by Giannouli et al. 14 showed that both EPIs PAβN and NMP produced no effect on rifampicin susceptibility of A. baumannii isolates. Specific rifampicin resistance mechanism of A. baumannii isolate was reported in chromosomal mutations in the RNA polymerase β-subunit rpoB target gene. 38 However, novobiocin resistance mechanism was not previously reported in A. baumannii. A study in Escherichia coli suggested an involvement of β-subunit of the DNA gyrase mukB target gene. 2
The strategy of resistance-modifying activity implies that H. antidysenterica extract and conessine could be coadministered with novobiocin or rifampicin to disrupt drug resistance mechanism in A. baumannii, thus increase the efficacy of both novobiocin and rifampicin. Hence, H. antidysenterica extract and conessine as RMAs might be of important benefit for the treatment of A. baumannii infections.
Conclusions
Nosocomial outbreak of infection with antibiotic-resistant A. baumannii has become a major public health problem. In this study, H. antidysenterica extract and its steroidal alkaloid conessine, as RMAs, demonstrated a markedly synergistic activity in combination with novobiocin and rifampicin against XDR A. baumannii. Interestingly, H. antidysenterica extract and conessine could restore the antibiotic activity by interfering with AdeIJK pump and impeded development of antibiotic resistance. Therefore, H. antidysenterica extract and conessine as RMAs may offer the combinatory therapy to restore antibiotic susceptibility in XDR organisms.
Footnotes
Acknowledgments
This work was supported by the Thailand Research Fund through the Royal Golden Jubilee PhD Program (Grant No. PHD/0041/2556) and TRF Senior Research Scholar (Grant No. RTA5880005), the Thailand Research Fund.
Disclosure Statement
No competing financial interests exist.