
Abstract
We analyzed the replicon types, sizes, and restriction fragment length polymorphism (RFLP) typing of plasmids carrying extended-spectrum β-lactamase (ESBL) genes in Klebsiella pneumoniae isolates from Taiwan. Fifty-one Escherichia coli transconjugant strains with plasmids from ESBL-producing K. pneumoniae from the Taiwan Surveillance of Antimicrobial Resistance III Program in 2002 were included. All the 51 plasmids carried a blaCTX-M gene, the majority of which were blaCTX-M-3 (28/51 [54.9%]). Plasmids ranged in size from 126 to 241 kb by S1 nuclease digestion and subsequent pulsed-field gel electrophoresis, and the most common plasmid size (37.3%) was 161–170 kb. The most common replicon type of plasmids was incompatibility group (Inc)A/C (60.8%). The IncA/C plasmids all carried blaCTX-M (blaCTX-M-3, -14, -15), and some also carried blaSHV (blaSHV-5, -12) genes. All 51 plasmids could be typed with PstI, and 27 (52.9%) belonged to 10 clusters. Thirty-eight of the 51 plasmids were typable with BamHI, and 21 plasmids (55.3%) fell into 7 clusters. Plasmids in the same cluster belonged to the same incompatibility group, with the exception of cluster C6. In conclusion, IncA/C plasmids are the main plasmid type responsible for the dissemination of ESBL genes of K. pneumoniae from Taiwan. RFLP with PstI possessed better discriminatory power than that with BamHI and PCR-based replicon typing for ESBL-carrying plasmids in K. pneumoniae in this study. Greater than 50% of plasmids fell into clusters, and >60% of cluster-classified plasmids were present in clonally unrelated isolates, indicating that horizontal transfer of plasmids plays an important role in the spread of ESBL genes.
Introduction
K
The prevalence of ESBL-producing K. pneumoniae (ESBL-KP) increased from 8% to 18.3% between 2002 and 2009 in Taiwan 10 ; however, an analysis of the replicon types of ESBL gene-bearing plasmids in ESBL-KP collected in 2002 has not been conducted, and very few relevant studies for K. pneumoniae were reported in Taiwan.11,12 This study determined the replicon type, plasmid size, clonal relatedness, and classification of these plasmids into RFLP-based clusters to reveal the role of plasmids in the spread of ESBL-associated antibiotic resistance and the epidemiologic implications.
Materials and Methods
Bacterial strains
Clinical isolates of ESBL-KP were collected by the Taiwan Surveillance of Antimicrobial Resistance (TSAR) III Program in 2002. As previously described in the Lin study, 10 the conjugation of the ESBL genes was carried out on 66 ESBL-KP isolates by broth mating out with the rifampicin-resistant strain of Escherichia coli (JP-995) or azide-resistant strain of E. coli (J53) as recipients. Transconjugants were selected on MacConkey agar plates containing rifampicin (100 mg/L) or azide (100 mg/L), as appropriate, and cefotaxime (5 mg/L) or ceftazidime (5 mg/L). Fifty-seven (86.4%) strains were transferable, and the ESBL genes in E. coli transconjugants were characterized by Lin et al. 10 Fifty-one E. coli transconjugant strains harboring plasmids from clinical isolates of ESBL-KP were included in this study. These ESBL-KP strains were isolated from 14 hospitals located in the 4 geographic regions (north, west, south, and east) of Taiwan. To characterize the clonality of ESBL-KP strains, pulsed-field gel electrophoresis (PFGE) groups were previously determined in the Lin study. 10
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing of transconjugant strains was performed by the standard broth microdilution method according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI). 13 The following antimicrobial agents were tested: ampicillin, amikacin, ceftazidime, cefmetazole, cefazolin, ertapenem, cefepime, cefoxitin, gentamicin, levofloxacin, imipenem, ampicillin–sulbactam, trimethoprim–sulfamethoxazole, and piperacillin–tazobactam.
Plasmid replicon typing
In all 51 transconjugant strains, the plasmid incompatibility group was determined by a PBRT scheme proposed by Johnson and Nolan, 14 who modified the procedure described by Carattoli et al. 6 Three primer panels, including 18 primer pairs, were used to perform 3 multiplex PCRs. Primers used in plasmid replicon typing were described previously.6,14 Bacterial template DNA was prepared using a Geneaid DNA Isolation Kit (Geneaid, Taipei, Taiwan) according to the manufacturer's instructions. For each PCR panel, equal amounts of each primer (0.1 mM) were combined in a pooled primer tube. The PCR amplification was performed in 20 μl volumes containing 200 ng bacterial template DNA, 0.2 mM dNTP, 2 mM MgCl2, 10× PCR buffer, 1 μl primer pool, and 1 U Taq DNA polymerase (Invitrogen, Carlsbad, CA) in a GeneAmp PCR System 2720 (Applied Biosystems, Foster City, CA). The PCR conditions were as follows: 5 min at 94°C; 30 cycles of 30 sec at 94°C, 30 sec at 60°C, and 90 sec at 72°C; and a final extension of 5 min at 72°C. Amplicons were visualized after separation by electrophoresis on a 2% agarose gel.
The IncF replicon sequence typing (RST) scheme was performed according to the scheme proposed by Villa et al. 15 Alleles were assigned by submitting the amplicon sequence to the Web site www.pubmlst.org/plasmid/.
S1 nuclease digestion and subsequent PFGE to determine plasmid size
The sizes of the plasmids were determined by PFGE after S1 nuclease digestion (S1-PFGE), as previously described, 16 with some modifications. S1-digested slices were loaded onto an agarose gel, and the gel was run on a CHEF-DR III system (Bio-Rad, Hercules, CA) at 14°C, with a switch time of 7–35 sec for 20 hr at 6 V/cm on a 120° angle in 0.5× TBE buffer.
Plasmid RFLP analysis
Plasmids were extracted using a Qiagen Plasmid Midi Kit (Qiagen, Hilden, Germany) following the manufacturer's instructions. For RFLP analysis, purified plasmid DNA was digested with BamHI or PstI restriction enzyme (New England Biolabs, Ipswich, MA) and subjected to electrophoresis on a 1% agarose gel at 100 V for 3 hr. Plasmid RFLP fingerprints were analyzed using GelCompar II software 6.5 (Applied Maths, Austin, TX), and dendrograms of the patterns were constructed using the unweighted pair group method with arithmetic mean (UPGMA) based on the Dice similarity index.
Results
ESBL genes in E. coli transconjugants
The ESBL genes in E. coli transconjugants were previously characterized by Lin et al. 10 All 51 E. coli transconjugants had blaCTX-M genes, including the most prevalent (blaCTX-M-3: 28/51 [54.9%]), followed by blaCTX-M-14 (12/51 [23.5%]) and blaCTX-M-15 (11/51 [21.6%]; Table 1). Twenty-five of the 51 E. coli transconjugants carried blaSHV-12 in addition to blaCTX-M genes: 17 of them carried blaCTX-M-3, 3 carried blaCTX-M-14, and 5 carried blaCTX-M-15. Furthermore, blaSHV-5 coexisted with blaCTX-M-3 or blaCTX-M-15 in four and six transconjugants, respectively (Table 1).
ESBL genes were characterized in the Lin study. 10
Non-β-lactams are underlined. AM, ampicillin; AN, amikacin; CAZ, ceftazidime; CMZ, cefmetazole; CZ, cefazolin; FOX, cefoxitin; GM, gentamicin; IPM, imipenem; SAM, ampicillin–sulbactam; SXT, trimethoprim–sulfamethoxazole; TZP, piperacillin–tazobactam.
Plasmids with ≥80% similarity in the RFLP patterns were grouped into clusters.
PFGE groups of ESBL-KP were determined in the Lin study. 10 The locations of hospitals from which the ESBL-KP were isolated were in the northern (N), western (W), southern (S), and eastern (E) regions of Taiwan.
“—” Indicates that plasmids did not yield RFLP patterns with BamHI.
FAB formulae of plasmids by IncF RST were K2:A-:B- (transconjugants 103, 104, 114, and 199) and K9:A-:B- (transconjugant 196).
FAB formula of the IncFII plasmid was F2:A-:B- (transconjugant 136).
ESBL, extended-spectrum β-lactamase; ESBL-KP, ESBL-producing Klebsiella pneumoniae; PFGE, pulsed-field gel electrophoresis; RFLP, restriction fragment length polymorphism; RST, replicon sequence typing.
Resistance profile of E. coli transconjugants
The resistance profile of E. coli transconjugants is shown in Table 1. All strains were resistant to ampicillin. Most of the strains were resistant to cefazolin (49/51 [96.1%]), followed by gentamicin (37/51 [72.5%]) and ceftazidime (32/51 [62.7%]). One (2.0%) strain (transconjugant 123) was resistant to imipenem. Carbapenemase genes of transconjugant 123 were detected by PCR amplification as previously described. 17 The blaIMP-8 was harbored in this transconjugant strain. Among non-β-lactams, resistance to gentamicin (72.5%) was most often cotransferred with the ESBL genes, followed by trimethoprim–sulfamethoxazole (24/51 [47.1%]) and amikacin (16/51 [31.4%]).
Plasmid replicon typing
A three-panel multiplex PCR protocol was used to examine the 51 E. coli transconjugants carrying ESBL genes for the presence of 18 plasmid replicon types. RST for IncF plasmids was performed to subcategorize IncF plasmids. The most frequently detected replicon types were A/C (60.8%), followed by L/M (9.8%), FIIK (9.8%), N (3.9%), FII (2.0), and F (2.0%; Table 1). Among the five IncFIIK replicons, the FAB formulae by IncF RST were K2:A-:B- in four plasmids (transconjugants 103, 104, 114, and 199) and K9:A-:B- in one plasmid (transconjugant 196). The FAB formula of the IncFII plasmid was F2:A-:B- (transconjugant 136). Two transconjugants carried two different replicon types on a single plasmid, one with A/C and N and the remaining transconjugant with L/M and N. Additionally, the replicon type could not be determined in four transconjugants (7.8%).
Table 1 reveals the association of the predominant IncA/C replicon with blaCTX-M alone or blaCTX-M coexisting with blaSHV genes, mainly blaCTX-M-3 and blaSHV-12 (9/31 [29%]), followed by blaCTX-M-14 (8/31 [25.8%]), blaCTX-M-15 and blaSHV-5 (5/31 [16.1%]), blaCTX-M-15 and blaSHV-12 (3/31 [9.7%]), blaCTX-M-3 (2/31 [6.5%]), blaCTX-M-3 and blaSHV-5 (2/31 [6.5%]), and blaCTX-M-14 and blaSHV-12 (2/31 [6.5%]). The IncL/M and IncFIIK replicons were associated with blaCTX-M-3 or blaCTX-M-3 coexisting with blaSHV-5 or blaSHV-12 (except the plasmid with blaCTX-M-15 and blaSHV-5 in transconjugant 196).
Plasmid size determined by S1 nuclease digestion and subsequent PFGE
S1 nuclease–PFGE analysis indicated that all E. coli transconjugants contained a single plasmid (size range = 121–180 kb), with the exception that two transconjugants harbored plasmids larger than 200 kb (Table 1). The most common plasmid size (19/51 [37.3%]) was 161–170 kb. The A/C replicon, blaCTX-M, and blaSHV were distributed among various-sized plasmids.
RFLP analysis
Thirty-eight of the 51 plasmids were successfully characterized using BamHI, and 21 plasmids (21/38 [55.3%]) were classified into 7 clusters (CI–CVII), with at least 80% similarity (Fig. 1 and Table 1). The similarities in RFLP patterns of 17 unclassified plasmids were smaller than 75%. All plasmids in the same cluster belonged to the same incompatibility group, with the exception of two plasmids in cluster IV in which incompatibility groups could not be determined.
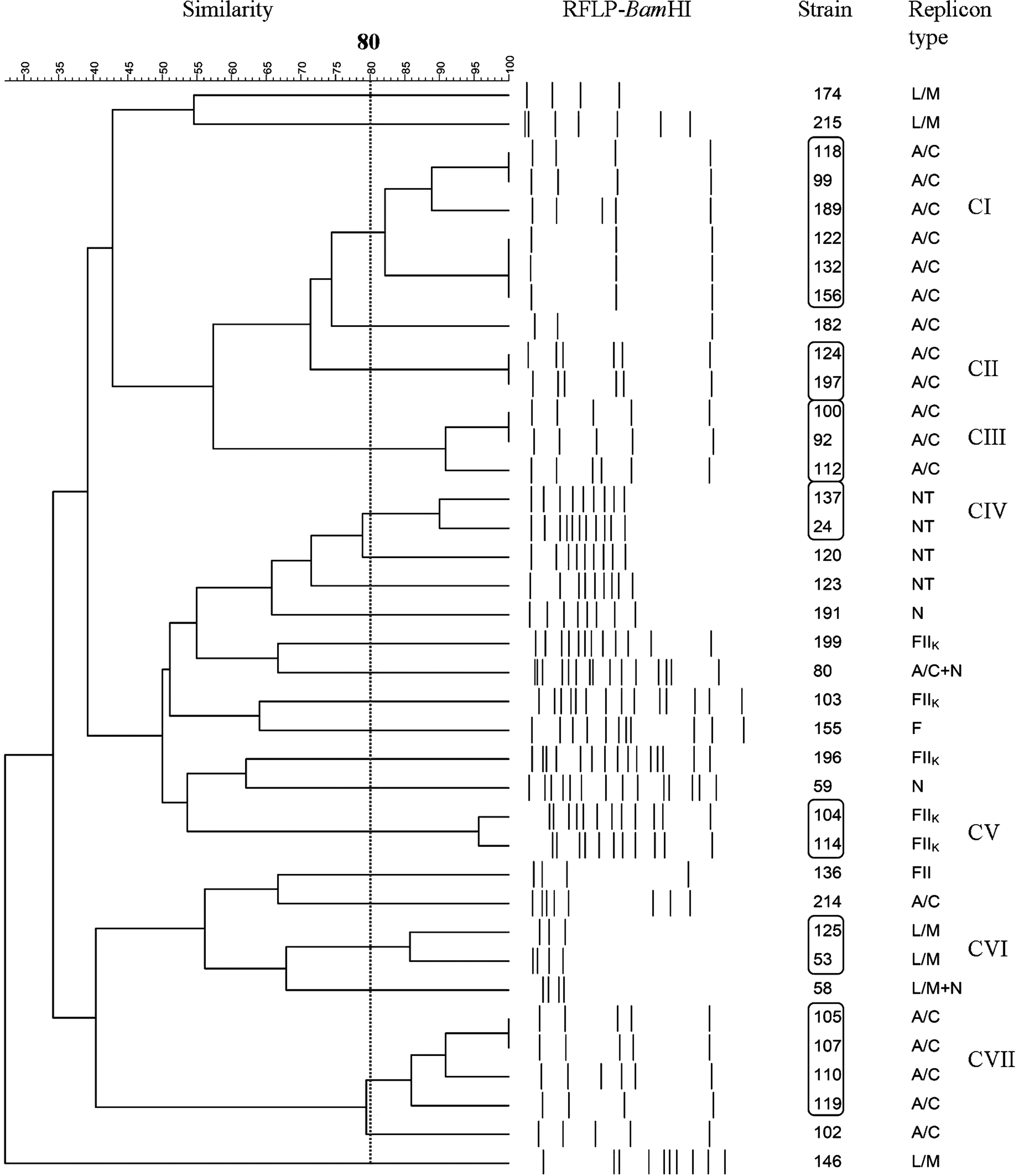
Dendrogram of RFLP patterns of plasmids digested with BamHI. Plasmids with ≥80% similarity in their RFLP patterns were grouped into clusters. Twenty-one of 38 plasmids belonged to clusters (CI–CVII). RFLP, restriction fragment length polymorphism.
Although 13 of the 51 plasmids did not yield RFLP patterns with BamHI, all plasmids were successfully typed using PstI. Twenty-seven (52.9%) of the 51 plasmids were separated into 10 clusters (C1–C10) based on at least 80% RFLP similarity (Fig. 2 and Table 1). IncA/C plasmids were the most prevalent in these 10 clusters (74.1% [20/27]). Plasmids in the same cluster belonged to the same incompatibility group, except for the plasmids in cluster C6. ESBL genes (blaCTX-M and blaSHV) were present in both cluster-classified and cluster-unclassified plasmids (Table 1).

Dendrogram of RFLP patterns of plasmids digested with PstI. Plasmids with ≥80% similarity in their RFLP patterns were grouped into clusters. Twenty-seven of 51 plasmids belonged to 10 clusters (C1–C10).
Plasmids in strains 24 and 137 with nontypable replicons could be classified into cluster CIV by BamHI restriction (Fig. 1 and Table 1). Although plasmids with nontypable replicons could not be classified into clusters by PstI, the relationship was demonstrated by RFLP by BamHI analysis. This indicated that plasmid RFLPs can be helpful in understanding plasmid relatedness when incompatibility groups are not identifiable by PCR replicon typing.
Comparing the RFLP profiles generated by BamHI to those generated by PstI, clusters CI, CIII, CV, CVI, and CVII identified by BamHI corresponded to the C9, C10, C7, C4, and C8 identified by PstI, respectively, even though the number of plasmids in the clusters was not all the same (Table 1).
PFGE analysis
PFGE groups of the ESBL-KP strains were determined in the Lin study. 10 The PFGE analysis revealed that 19 of the 51 isolates (37.3%) belonging to 5 groups (A, D, E, F, and G) indicated clonal relatedness (Table 1). The remaining 32 isolates had distinct PFGE patterns and were clonally unrelated. Groups A, D, E, F, and G comprised two, three, four, two, and eight isolates, respectively. Isolates in the same group were from the same hospital, indicating some ESBL-KP isolates were clonally spread in some hospitals. Among 27 cluster-classified plasmids by PstI restriction, 17 (63% [17/27]) plasmids were present in clonally unrelated isolates (Table 1). Moreover, only 6 clonally related isolates carried RFLP-clustered plasmids typed by BamHI and PstI (105, 107, 122, 100, 112, and 118), while the other 13 clonally related isolates carried plasmids of different clusters or cluster-unclassified plasmids (Table 1).
Discussion
In the current study, the most prevalent replicon type in ESBL-KP isolates in Taiwan was IncA/C (60.8%), which is similar to that reported in Paraguay (48% of K. pneumoniae isolates with IncA/C plasmids) 18 but different from Tunisia (93.8% of K. pneumoniae isolates with IncFIIA), 19 9 Asian countries (81.8% with IncFIIA), 20 China (66.7% with IncFII), 21 Bulgaria (58.3% with IncL/M), 22 Scotland (IncN), 23 Pakistan (IncF), 24 and the Central African Republic (IncF). 25 Thus, the prevalent plasmid replicon types are diverse in ESBL-KP worldwide.
Very few studies reported the replicon types of plasmids in clinical isolates from Taiwan, including K. pneumoniae. Lee et al. 12 reported an IncFIIA plasmid among the four self-transferable plasmids in ESBL-KP (the other three were nontypable) from a medical center in southern Taiwan between 2004 and 2005. Wang et al. 11 found that the IncF replicon type was harbored in 7 of the 16 KPC-2-positive, carbapenem-nonsusceptible K. pneumoniae isolates from four hospitals between 2010 and 2012. These results of K. pneumoniae in Taiwan are different from those of the current study.
Some studies have reported a strong link between ESBL genes and particular plasmid types in K. pneumoniae, that is, the association of blaCTX-M-15 with IncF-type plasmid,19,22,25,26 IncN, 23 or IncR, 27 the association of blaCTX-M-3 with IncL/M, 22 the association of blaCTX-M-14 with IncF, 28 and the association of blaSHV-12 with IncR or IncHI2. 27 In contrast, the current study revealed a predominant linkage of IncA/C plasmids with ESBL genes (blaCTX-M alone or blaCTX-M coresident with blaSHV genes), indicating IncA/C plasmids play an important role in the dissemination of ESBL genes in clinical strains of K. pneumoniae in Taiwan. Furthermore, resistances to non-β-lactams were cotransferred with the ESBL genes, that is, gentamicin, trimethoprim–sulfamethoxazole, or amikacin in the current study. Replicon A/C has also been associated with TEM β-lactamase genes,26,29–31 AmpC β-lactamases CMY (mainly CMY-2),29,32 and more recently associated with VIM carbapenemases. 29 The plasmids were a platform for distribution of the genes of resistance determinant enzymes.
Studies have described that the IncF plasmid family (with different subgroups) is the most prevalent in ESBL-carrying K. pneumoniae worldwide and associated with blaCTX-M genes, specifically blaCTX-M-15.19,20,25,26,29,33–35 Lee et al. 20 reported an IncFIIA-type plasmid predominantly associated with blaCTX-M-15 in nine Asian countries. Zhuo et al. 35 found an epidemic IncFII plasmid carrying blaCTX-M-15 in southern China. These results of Asian areas are different from those of the current study; however, international traffic and geographic proximity may facilitate the dissemination of plasmids and influence the trend of dominant plasmid replicon types and the linkage with ESBL genes. Moreover, clonal spread also occurred in the Asian studies, including the current study.20,35 Therefore, continuous monitoring and characterization of plasmids together with monitoring of clones are necessary.
Plasmid RFLP typing demonstrated that >50% of plasmids fell into clusters (55.3% typed by BamHI and 52.9% by PstI), and plasmids in the same cluster belonged to the same incompatibility group, with the exception of the cluster C6 typed by PstI restriction. Furthermore, 63% of the 27 cluster-classified plasmids by PstI restriction were present in clonally unrelated isolates. This highlights that the circulation of similar plasmids in K. pneumoniae strains is mediated by horizontal transfer. Various RFLP patterns were identified even in the same incompatibility group. This may be partially attributed to differences in the targets of replicon typing and RFLP analysis. The targets for PBRT are those genes associated with replication, including replication genes, ori sites, iteron sequences, and plasmid-partitioning genes6,14; however, the targets for RFLP are whole plasmid sequences recognized by restriction enzymes. RFLP is more discriminative than PBRT. RFLP with PstI possessed better discrimination power than RFLP with BamHI. Furthermore, plasmids in the same incompatibility group with distinct RFLP patterns suggest that the composition of plasmids with similar backbones is continuously evolving.
Replicon typing showed that ESBL genes (blaCTX-M and blaSHV) are carried by plasmids of different incompatibility groups other than the predominant IncA/C. Cluster analysis of plasmid RFLPs using PstI revealed that ESBL genes were distributed over cluster- and noncluster-classified plasmids. This indicated that ESBL genes can be found on similar or different plasmid platforms. Genetic elements, such as insertion sequences associated with ESBL genes, have been described. For example, studies have found that the insertion sequence, ISEcp1, is frequently located upstream of blaCTX-M-3, blaCTX-M-14, and blaCTX-M-1528,33,34,36,37; blaCTX-M-14 is flanked by ISEcp1 and IS903, 36 and blaSHV-5 and blaSHV-12 are linked to IS26.36,38 These genetic elements may be involved in the mobilization of ESBL genes and contribute to dissemination among plasmids.36,37 The genetic environment of ESBL genes within the plasmids warrants further investigation.
In summary, this study demonstrated that IncA/C that carried blaCTX-M and blaSHV genes was the predominant replicon type in clinical ESBL-KP isolates from Taiwan. Plasmid RFLP typing can help determine the relatedness of plasmids when incompatibility groups cannot be determined. PstI possessed better discriminatory power than BamHI for the plasmids included in the current study. Greater than 50% of plasmids were divided into clusters, and >60% of cluster-classified plasmids by PstI restriction were present in clonally unrelated isolates, indicating that horizontal transfer of plasmids plays an important role in the wide distribution of ESBL genes.
Footnotes
Acknowledgments
This work was supported by the Department of Health, Taiwan (DOH102-DC-1508), National Health Research Institutes, and Kaohsiung Medical University Hospital, Taiwan.
Disclosure Statement
No competing financial interests exist.