
Abstract
Sequence type (ST) 239 with SCCmec type III methicillin-resistant Staphylococcus aureus (ST239-MRSA-III) is the most predominant multidrug-resistant clone in China. The subclone ST239-MRSA-III-t037 has been gradually replaced with ST239-MRSA-III-t030 since 2000. Subclones are characterized by drug resistance profiles. However, the mechanisms of the clonal dynamics and determinants of distinct drug resistance remain poorly understood. In the present study, 12 ST239-MRSA-III-t030 and 12 ST239-MRSA-III-t037 strains were collected from Chongqing, Guangzhou, and Shanghai; these strains were selected and investigated in terms of t030/t037 strain pairs. Independent growth curve assay revealed that the ST239-MRSA-III-t030 strains grew more rapidly, with significantly shorter doubling times, than the ST239-MRSA-III-t037 strains (p < 0.001). The ST239-MRSA-III-t037 strains exhibited slightly to moderately higher (3–13%) fitness cost than the ST239-MRSA-III-t030 strains in a competition assay in vitro. The ST239-MRSA-III-t037 strains yielded lower bacterial loads in the kidneys of the infected mice than the ST239-MRSA-III-t030 rivals in a coinfection assay (p < 0.05). The ST239-MRSA-III-t030 strains were resistant to rifampicin but susceptible to trimethoprim/sulfamethoxazole (SXT). In contrast, the ST239-MRSA-III-t037 strains were susceptible to rifampicin but resistant to SXT. The genetic determinants of the resistance to rifampicin and SXT in the MRSA strains were determined. Our results suggest that the relatively low fitness cost and characteristic drug resistance phenotype can help explain the current predominance of these ST239-MRSA-III-t030 strains in Chinese hospitals.
Introduction
M
In China, ST239-MRSA with the staphylococcal cassette chromosome mec type III (ST239-MRSA-III) is the most prevalent clone that accounts for >50% of MRSA infections.6,13 The ST239-MRSA-III strains are mainly associated with the two spa types: t030 and t037.20,26 A comparison of genome phylogeny by spa typing has suggested that t037 represents an ancestral ST239 spa type. 12 The ST239-MRSA-III-t037 was the most dominant subclone in Beijing before 2000, but this strain has been gradually replaced with ST239-MRSA-III-t030. 4 A survey was conducted in 2014 using isolates collected from seven tertiary care hospitals in distinct geographical locations; the survey has revealed that ST239-MRSA-III-t030 accounts for 73.5% of the tested MRSA isolates; in contrast, ST239-MRSA-III-t037 accounts for only 4% of these isolates. 5 This result suggests that t030 has successfully replaced t037 as the most frequent MRSA subclone in China. However, the reasons for the clonal dynamics remain poorly understood. A comparative microarray study has shown that three gene clusters, namely, vSa4, phage φSa1, and φSa3, are unique to MRSA-t030 and may contribute to the replacement of MRSA-t037. 19 Whole-genome sequencing with representative t037 strains from Hong Kong and t030 strains from Beijing has further revealed that t037 belongs to the traditional Asian clade; in contrast, t030 belongs to the newly emerged Turkish clade. 29 Nevertheless, the mechanisms underlying the MRSA clonal dynamics may be complex. The distinct clonal adaptive capability, environmental antibiotic pressure, and active social activity are the proposed reasons for the dissemination of a specific clone. 18
The MRSA strains may suffer severe fitness cost upon developing resistance to certain antibiotics, such as resistance to gentamicin or ciprofloxacin, and this fitness cost could be a disadvantage for clonal spreading in a given region.14,18 Epidemiological studies have revealed that most ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains from Chinese hospitals are resistant to gentamicin and ciprofloxacin; however, most ST239-MRSA-III-t030 strains are resistant to rifampicin but susceptible to trimethoprim/sulfamethoxazole (SXT); in contrast, most ST239-MRSA-III-t037 strains are susceptible to rifampicin but resistant to SXT.6,20 The fitness cost and determinants associated with the characteristic drug resistance phenotypes of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 subclonal strains remain unclear. In this study, 12 ST239-MRSA-III-t030 and 12 ST239-MRSA-III-t037 strains isolated from Chongqing, Guangzhou, and Shanghai were selected and investigated in t030/t037 strain pairs. Our study aimed to determine (1) the independent growth curve and doubling time (DT) of the selected ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains, (2) the relative fitness of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains through competitive culture and coinfection assay, and (3) the genetic determinants of the rifampicin and SXT resistance of MRSA strains.
Materials and Methods
Bacterial strains and growth conditions
In our previous study on 309 MRSA isolates recovered from nine hospitals in six cities from 2009 to 2012, 87 ST239-MRSA-III-t030 strains and 46 ST239-MRSA-III-t037 strains were characterized. 6 In the present study, 24 characterized MRSA isolates, including 12 ST239-MRSA-III-t030 strains and 12 ST239-MRSA-III-t037 strains collected from southwest (Chongqing), southeast (Guangzhou), and middle east (Shanghai) regions in China, were selected. All selected strains were sensitive to linezolid, teicoplanin, and vancomycin but resistant to penicillin, oxacillin, levofloxacin, erythromycin, gentamicin, and ciprofloxacin. The ST239-MRSA-III-t030 strains were resistant to rifampicin (12/12, 100%) but susceptible to SXT (12/12, 100%). In contrast, the ST239-MRSA-III-t037 strains were all susceptible to rifampicin (12/12, 100%) but resistant to SXT (12/12, 100%) (Table 1). The strains were paired based on the distinct geographical locations where the strains were recovered and the sources from which the strains were isolated. The strains were grown in a brain–heart infusion broth (BHIB; Oxoid) or on BHI agar (BHIA, 1.5%) plates supplemented with rifampicin or SXT when needed.
Determination of MIC for each strain was repeated at least thrice to confirm reproducibility. The independent growth curve for each strain was conducted at least thrice, and its DT is calculated as described in the Materials and Methods section and expressed in minutes ± standard deviation. The level of significance for strain DT in each pair was determined by two-tailed Student's t-test, and p-value is indicated.
DT, doubling time; MIC, minimum inhibitory concentration; RIF, rifampicin; SXT, trimethoprim/sulfamethoxazole, the data indicated as ratios of trimethoprim/sulfamethoxazole.
Growth curve and DT determination
The independent growth curves of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains were determined as previously described. 31 Briefly, the tested strains were cultured in BHIB at 37°C with shaking overnight, and 0.2 ml of each culture was added to 20 ml of fresh BHIB in a sterile 50-ml flask. The optical density readings at 600 nm (OD600) were obtained every 15 min for 24 hr after inoculation. DT was calculated by the slope of linear line obtained from the semilogarithmic graph of the growth curve and with the following formula: DT = [(t2 − t1) × log2]/(log OD600 at t2 − log OD600 at t1), where t2 and t1 are the times at the end and start of the logarithmic growth phase, respectively. 18 Other growth media (Trypticase soy broth [TSB] and Luria-Bertani broth [LBB]) were tested and shown to give similar results. All isolates were tested through at least three independent experiments, and the standard deviation of the mean was reported.
Competitive culture in vitro and relative fitness calculation
Two strains in each strain pair were compared for competitive growth in a mixed culture as previously described. 18 Briefly, bacteria were cultured on BHIA at 37°C for 24 hr and suspended in a T2 buffer [70 mM NaCl, 30 mM K2SO4, 10 mM KH2PO4, 20 mM Na2HPO4·2H2O, 1 mM MgSO4·7H2O, 1 mM CaCl2·2H2O, and 0.001% gelatine (w/v)] to obtain a No. 0.5 McFarland standard density. Approximately, 10 μl of each suspension in a strain pair was mixed (totally 20 μl) and inoculated into 5 ml of BHIB. Afterward, the tubes were incubated at 37°C with gentle agitation. The number of viable bacteria was determined at the start (t = 0 hr) and at the end of the culture (t = 24 hr) by dilution plating in triplicate onto BHIA with or without rifampicin (4 mg/L) or SXT (4 mg/L trimethoprim and 76 mg/L sulfamethoxazole) and cultured at 37°C for 18 hr. The colony counts on the rifampicin-selective plate showed the bacterial levels of ST239-MRSA-III-t030, whereas those on the SXT-selective plate indicated the population density of ST239-MRSA-III-t037. 17 The colony numbers from the selective plates (rifampicin- and SXT-selective plates) and nonselective plates of each strain pair differed by <5%. The initial ratio [Nt037(0)/Nt030(0)] and final ratio [Nt037(24)/Nt030(24)] were obtained, and the relative fitness (F) was calculated with the following formula: F = ln[Nt037(24)/Nt037(0)]/ln[Nt030(24)/Nt030(0)], where Nt037(t) and Nt030(t) represent the bacterial levels of the ST239-MRSA-III-t037 and ST239-MRSA-III-t030 strains, respectively, at 0 or 24 hr. 24 The fitness cost was then calculated with the following formula: (1 − F) × 100%. Three independent experiments were performed in each pair of strains.
Coinfection in a mouse model
All in vivo studies were undertaken following institutional and national standards for animal care and experimentation. The protocol was approved for use by the Laboratory Animal Welfare and Ethics Committee of the Third Military Medical University.
One representative t037/t030 strain pair from each location was randomly selected (CQ40/CQ29 from Chongqing, SH31/SH50 from Shanghai, and GZ62/GZ01 from Guangzhou) to test the capability of paired strains in coinfection in a mouse model. The overnight culture of each strain was diluted 500-fold in fresh BHIB and was incubated at 37°C with shaking until an OD600 of 0.8 was achieved. The bacterial cells were pelleted and washed once with 10 mM phosphate-buffered saline (PBS, pH 7.2) and then adjusted to 1 × 108 colony-forming units per milliliter (CFU/ml). Afterward, 300 μl of this suspension was mixed with an equal volume of suspension of the counterpart strain in a strain pair to make a 600-μl mixture and then was intravenously injected into five 8-week-old female BALB/c mice (100 μl for each). The mice were killed by cervical dislocation 3 days after the challenge. The kidneys were removed and homogenized separately in 1 ml of PBS with 1% Triton X-100 as described. 2 The number of viable bacteria in the suspension was determined by dilution plating in triplicate onto selective and nonselective BHIA as described above.
Detection of genetic determinants associated with rifampicin and SXT resistance
To identify the genetic determinants that were associated with the distinct resistance phenotypes of t030/t037 strain pairs, several sets of primers were designed (Table 2) and used to amplify the candidate genes of interest. As for rifampicin resistance, an internal gene fragment containing rifampicin resistance–determining region (RRDR) of staphylococcal rpoB (nucleotides 1,256 to 1,648) was amplified and sequenced. The candidate genes associated with S. aureus resistance to SXT, including dhps (encodes dihydropteroate synthase, DHPS), dfrB (encodes dihydrofolate reductase, DHFR), and dfrG (encodes a variant DHFR), were also amplified by polymerase chain reaction and sequenced to identify the possible SXT-resistant determinants.15,23,28 The target sequences of the tested strains were then compared to MRSA N315 (GenBank accession no. NC_002745), a reference strain with susceptible phenotypes to rifampicin and SXT, using the Clustalw software as previously described. 32
Sequences were designed based on the genome of MRSA N315 (GenBank accession no. NC_002745).
Statistical analysis
Data were analyzed using SPSS statistical software version 18. Fisher's exact test or t test was used to compare categorical variables. All analyses were two-tailed, and a p-value <0.05 was considered statistically significant.
Results and Discussion
Changes in MRSA predominant clones have been increasingly observed over the last two decades.4,5,14 The Portuguese clone ST239-MRSA-III-t421 predominant until the early 1990s in Portugal was replaced by ST247-IA. Later, the Brazilian clone ST239-MRSA-III/IIIA-t037 emerged in 1995 and was replaced by EMRSA-15. 1 In Argentina, the Brazilian clone was displaced by the Cordobes clone. 27 In Hungary, the Hungarian clone was replaced by the southern German and New York/Japan clones. 7 However, the possible mechanisms underlying the MRSA clonal dynamics may be sophisticated. The capability to adapt to environmental conditions is a major physiological determinant of bacterial growth and survival. A previous study investigated the replacement of CC30 by CC22 in the United Kingdom and demonstrated that CC22 MRSA is fitter than CC30 in the independent growth and competition assays. 17 The fitness cost can be considered as a bacteriological explanation for the demise of the first international MRSA epidemic in Denmark. 9
To address the replacement of ST239-MRSA-III-t037 by ST239-MRSA-III-t030 in China, the independent growth curves and DTs of the ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains in BHIB were compared (Fig. 1 and Table 1). As shown in Fig. 1, the ST239-MRSA-III-t030 strains produced higher biomasses compared with their counterparts by achieving high OD600 levels after 24 hr of culture. The ST239-MRSA-III-t030 strains displayed short DTs (mean DT = 26.96 ± 0.27 min), whereas the ST239-MRSA-III-t037 strains presented long DTs (mean DT = 33.50 ± 0.62 min). The difference between the DTs of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 was highly significant (p < 0.001). Furthermore, the identical experiments were performed in TSB and LBB growth media with all strains of interest. The analysis of the DTs obtained with the two media also revealed that ST239-MRSA-III-t030 and ST239-MRSA-III-t037 can be distinguished on the basis of the DTs (p < 0.001). In the competitive experiments, while the ST239-MRSA-III-t037 and ST239-MRSA-III-t030 strains were initially inoculated in almost equal numbers [initial ratio Nt037(0)/Nt030(0) = 1.33 ± 0.43], the ST239-MRSA-III-t037 strains represented 25–44% of the population after a 24-hr incubation period [final ratio Nt037(24)/Nt030(024) = 0.59 ± 0.14, Table 3]. The ST239-MRSA-III-t037 strains presented slight-to-moderate fitness cost (3–13%) compared with that of the ST239-MRSA-III-t030 strains (Table 3). Together, these data demonstrated that the ST239-MRSA-III-t030 strains can outgrow the ST239-MRSA-III-t037 strains in the mixed culture. The decrease in ratio resulted from the relatively severe fitness cost of ST239-MRSA-III-t037 strains over the ST239-MRSA-III-t030 strains.
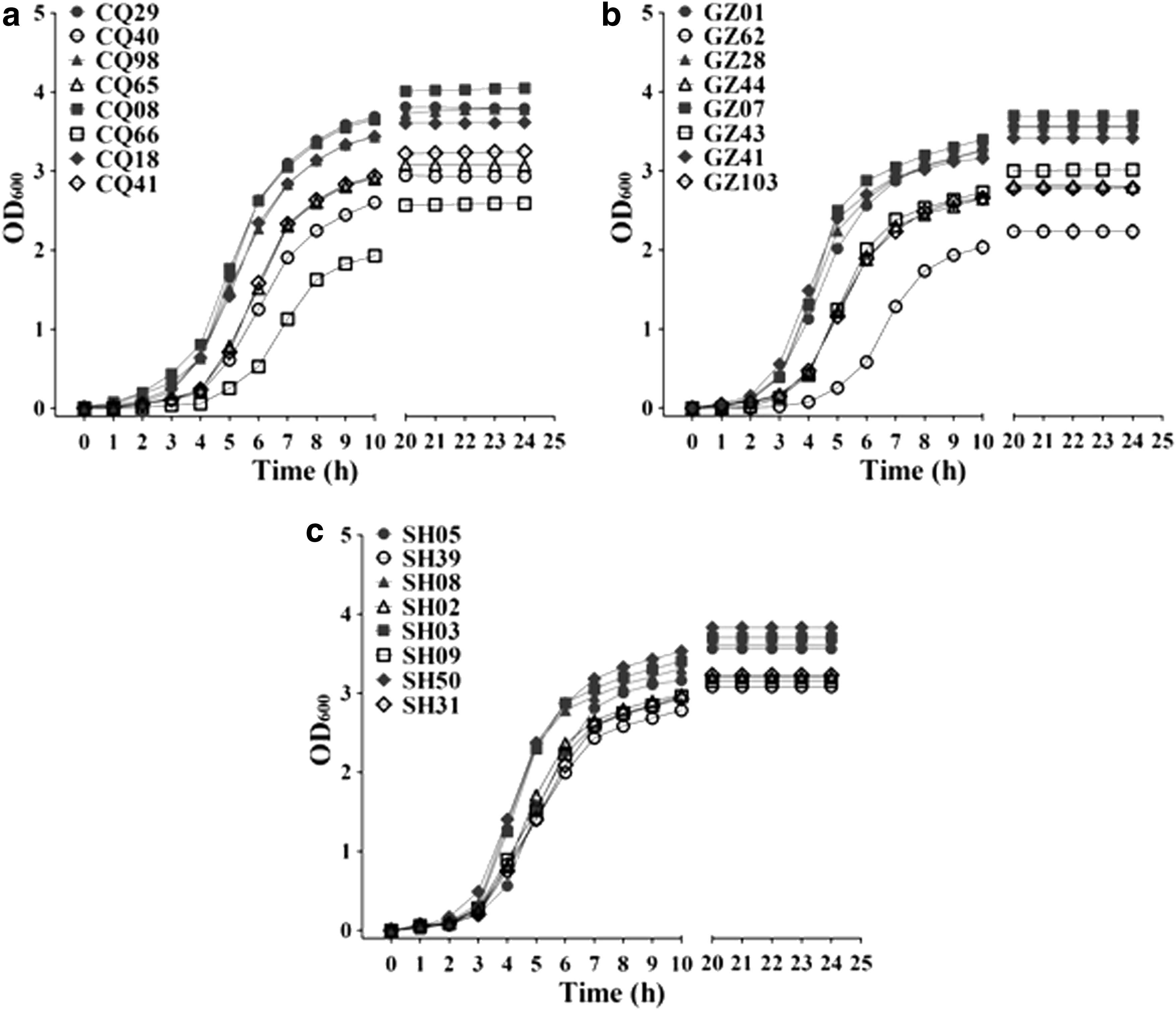
Growth curves of the selected MRSA strains. Independent growth curves of ST239-MRSA-III-t030 (filled symbols) and ST239-MRSA-III-t037 (empty symbols) strains recovered from
At the start of inoculation (t = 0 hr).
At the end of inoculation (t = 24 hr).
Fitness reduction is significant, as determined by one-sample t-test (p < 0.05).
SD, standard deviation.
We also demonstrated that both the ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains can establish infections in the mouse model. However, in the coinfection experiments, the average numbers of the viable bacteria of ST239-MRSA-III-t030 in the kidneys determined 3 days after the challenge were (9.06 ± 0.40) × 106, (8.62 ± 0.45) × 106, and (8.33 ± 0.58) × 106 CFU per kidney for CQ29, SH50, and GZ01, respectively. The values were significantly higher than those of ST239-MRSA-III-t037 strains with (3.70 ± 0.80) × 106, (4.75 ± 0.56) × 106, and (4.36 ± 0.58) × 106 CFU per kidney for CQ40, SH31, and GZ62, respectively (p < 0.05). These data provide a rational explanation, at least a part, for currently maintaining a low proportion of ST239-MRSA-III-t037 infections in Chinese hospitals.
Antibiotic resistance may contribute to the clonal dynamics by increasing the fitness burden of bacteria. 22 Laurent et al. 18 found that gentamicin-susceptible MRSA (GS-MRSA) has a competitive advantage over gentamicin-resistant MRSA (GR-MRSA), which facilitates the progressive replacement of GR-MRSA clone by GS-MRSA in France. Horváth et al. 14 reported that the MRSA strains cause severe fitness costs when these strains develop resistance to fluoroquinolone-type antibiotics (ciprofloxacin), and this fitness cost is diverse across MRSA isolates and may govern the clonal dynamics of MRSA in Hungary. The selected ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains in this study were all resistant to gentamicin and fluoroquinolone-type antibiotics (levofloxacin and ciprofloxacin). To address the distinct resistance phenotypes of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains for rifampicin and SXT, we amplified and sequenced the RRDRs of all strains of interest and found that all the ST239-MRSA-III-t030 strains exhibited H481N+L466S double substitutions compared with those of the rifampicin-susceptible strain N315, whereas all the ST239-MRSA-III-t037 strains contain RRDRs identical to those of N315. These data revealed that the nonsynonymous double substitutions in RpoB (H481N+L466S) confer resistance to rifampicin in ST239-MRSA-III-t030 strains. The observation is consistent with a previous study conducted in Anhui province, China, which showed that H481N+L466S in MRSA RpoB is the most common genotype that confers rifampicin resistance (77/83, 92.8%). 32
Trimethoprim and sulfamethoxazole are synthetic antibacterial agents and have been used in combination (termed SXT) in clinics since 1968. 15 Trimethoprim acts as a competitive inhibitor of bacterial DHFR (encoded by the chromosomal dfrB gene). Two genetic mechanisms confer trimethoprim resistance in S. aureus, which are mutations in the dfrB gene and acquisition of extrachromosomal resistance genes that encode variant DHFRs, such as dfrA, dfrG, and dfrK.16,18,23 No mutations were found in the dfrB gene of the SXT-resistant ST239-MRSA-III-t037 strains. In 2005, Sekiguchi et al. 25 for the first time reported on dfrG in an MRSA clone in a hospital in Thailand. In 2014, Nurjadi et al. 23 reported that 94% of trimethoprim resistance in S. aureus from sub-Saharan Africa is mediated by the dfrG gene. Therefore, the presence of dfrG gene was assessed in ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains. Importantly, all the tested ST239-MRSA-III-t037 strains carried the dfrG gene, but none of the ST239-MRSA-III-t030 strains carried the gene. A recently described ST239-MRSA-III-t030 strain from Russia (16K) is also sensitive to trimethoprim and resistant to rifampicin with H481N+L466S mutations in its RpoB. 30 Compared with the TW20 genome (belonging to t037 subclone), the 16K genome presents a Tn5801-like transposon that lacks the dfrG gene, whereas the TW20 genome has a dfrG-carrying Tn5801-like transposon. 30 Sulfonamide drugs are competitive inhibitors of DHPS (encoded by the chromosomal dhps gene); as a result, folate biosynthesis in the bacterial cell is inhibited. 15 The dhps gene of all t030/t037 paired strains was amplified and sequenced, and no mutations were found. The minimum inhibitory concentrations (MICs) of ST239-MRSA-III-t037 strains against sulfamethoxazole alone were ≥512 mg/L, indicating that the presence of genetic determinants other than dhps in ST239-MRSA-III-t037 strains for the resistance to sulfamethoxazole exists and needs further investigation.
It has been reported that a single substitution in MRSA RpoB (i.e., H481Y) is sufficient to mitigate the binding affinity of rifampicin to RNA polymerase, resulting in a low-level rifampicin resistance. 11 In this case, a negative correlation could exist between rifampicin resistance and relative fitness. O'Neill et al. 24 found that the most common genotype mutations in British S. aureus (RpoB H481N+S529L) do not exhibit any fitness cost in vitro and are often associated with a high-level rifampicin resistance. They proposed that an additional intragenic mutation may compensate for the fitness costs associated with rifampicin resistance in clinical S. aureus strains. Although the laboratory-prepared double mutations (RpoB H481N+L466S, similar to our clinical ST239-MRSA-III-t030 strains) resulted in a slight fitness cost, the second mutation (L466S) could compensate ∼3% of the fitness cost compared with that observed with the H481N genotype alone. 24 The difference in the DTs of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 subclonal strains observed in this study could be ascribed to the relatively severe fitness costs of the SXT-resistant ST239-MRSA-III-t037 strains compared with those of the rifampicin-resistant ST239-MRSA-III-t030 strains, which may contribute to explain the current predominance of ST239-MRSA-III-t030 strains in Chinese hospitals by showing their potential to outgrow ST239-MRSA-III-t037 strains.
In conclusion, ST239-MRSA-III-t030 strains in Chinese hospitals displayed short DTs and exhibited their potential to outgrow ST239-MRSA-III-t037 strains in the mixed culture. Although the difference in bacterial loads in the kidneys of coinfected mice was observed, both the ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains can successfully establish infections in the mouse model, which is in agreement that both the ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains can be isolated from patients with MRSA infections, but the subclonal dynamics exists. The determinants for characteristic drug resistance in ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains were characterized in this study. The different resistance profiles found in t030 and t037 strains could be an important factor that promotes the quick dissemination of ST239-MRSA-III-t030 strains with relatively low fitness costs in China. Our future studies aim to determine the degree of bacterial fitness cost associated with the characteristic drug resistance of ST239-MRSA-III-t030 and ST239-MRSA-III-t037 strains.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81471993 and 31170159), New Drug Development grant from China (2012ZX09103301-038), and the Natural Science Foundation of Chongqing City (CSTC2011jjA10070).
Disclosure Statement
No competing financial interests exist.