
Abstract
A lack of an effective way to eliminate pathogenic bacteria hidden in the biofilm is a major problem in the treatment of chronic bacterial infections. Iminosugar derivatives are potential candidates for inhibitors of enzymes taking part in the biosynthesis of exopolysaccharides, which are forming bacterial biofilm. Investigated iminosugars were studied either at an early stage of biofilm formation or later on when the mature biofilm of Pseudomonas aeruginosa was already formed. A series of diverse iminosugar structures significantly inhibited biofilm formation, whereas they showed no influence on already formed biofilm. This indicates a possible mechanism of their action based on inhibition of exopolysaccharide backbone synthesis in the early stages of biofilm formation. Moreover, iminosugar derivatives did not show significant effect on the viable bacterial numbers in both early and mature biofilm forms. Importantly, they were not cytotoxic against human Caco-2 cells in vitro, which may be to their advantage in case of their medical application in preventing P. aeruginosa biofilm formation.
Introduction
I
Iminosugar derivatives also appear to be interesting candidates as potential inhibitors of glycosyltransferases involved in the biosynthesis of glycan chains 6 or of more specialized enzymes, such as UDP-galactopyranose mutase or UDP-galactofuranose transferases. 7 The latter are involved in the formation of galactofuranose units and production of Galf-containing polysaccharides.8,9 This unusual form of galactose is found in the exopolysaccharides of several bacteria, such as not only in Escherichia coli K-12 and Streptococcus pneumoniae but also in Lactobacillus species.10,11 So far, very few studies have been dedicated to the effect of iminosugars on bacterial biofilms: A brief report by a Chinese group indicates that iminosugars may be useful in caries prevention by affecting α-glucan biosynthesis, 12 and another recent study has provided evidence for a significant inhibitory action of 1-deoxynojirimycin (from mulberry leaves) on Streptococcus mutans biofilm formation.13,14
Biofilm is usually defined as surface-associated microbial communities, surrounded by an extracellular polymeric substance (EPS) matrix, comprising mainly polysaccharides, proteins, nucleic acids, and metabolites produced by bacteria or fungi. The biofilm produced by microorganisms enables acquisition of new ecological niches, and moreover, bacteria hidden in a biofilm matrix are highly resistant to a variety of antibiotics. Therefore, the so-called pathogenic biofilm is considered as one of the most important and difficult to cure form of the microbial pathogenicity.13,15,16
Pseudomonas aeruginosa (PAR) is a bacterial species causing important human infections. PAR is also responsible for biofilm-related infections, such as pneumonia in cystic fibrosis patients, and it is associated with chronic wounds in patients with chronic diabetic foot infection. Until recently, it has been thought that the main elements of the P. aeruginosa biofilm structures are alginate polysaccharides. However, at present, it is considered that the building of the three-dimensional structure of P. aeruginosa biofilm matrix begins from the synthesis of the extracellular exopolysaccharides composed of chains containing sugars, such as rhamnose, glucuronic acid, and galactosamine, and only the next step is to fill this initial structure with strongly gelling alginate scaffold.16–18
The aim of this study was to evaluate the effect of a series of diverse iminosugar structures to inhibit formation of P. aeruginosa biofilm. Additionally, the iminosugars were tested for their ability to influence P. aeruginosa populations in early and mature biofilm forms and cause apoptosis and/or necrosis of the human cells. In our in vitro study, the early and mature biofilm forms of P. aeruginosa were treated with the investigated iminosugars, which were designed to mimic on one hand hexopyranose structures having a pseudo-gluco (iminoxylitol) or a pseudo-galacto configuration (imino-L-arabinitol) and on the other hand two other compounds having a configuration resembling that of a galactofuranose.
Materials and Methods
Tested compounds
Investigated iminosugar derivatives have been synthesized in the Institute of Organic and Analytical Chemistry at the University of Orleans: HDIX, α-1-C-hexyl-1,5-dideoxy-1,5-imino-D-xylitol, a powerful inhibitor of lysosomal β-glucocerebrosidase;19,20 DDIA, β-1-C-(2,3-dihydroxypropyl)-1,5-dideoxy-1,5-imino-L-arabinitol, 21 has a galacto-like configuration and carries a polar aglycone; DHMBI, (1S,2S,3S,5S,9R)-1,2-dihydroxy-3-hydroxymethyl-5-p-methoxybenzylindolizidine, 22 a close analogue of the natural product steviamine, and -PDIA, β-1-C-propyl-1,4-dideoxy-1,4-imino-L-arabinitol, a pyrrolidine analogue of HDIX. Chemical structures of the compounds are presented in Figure 1.
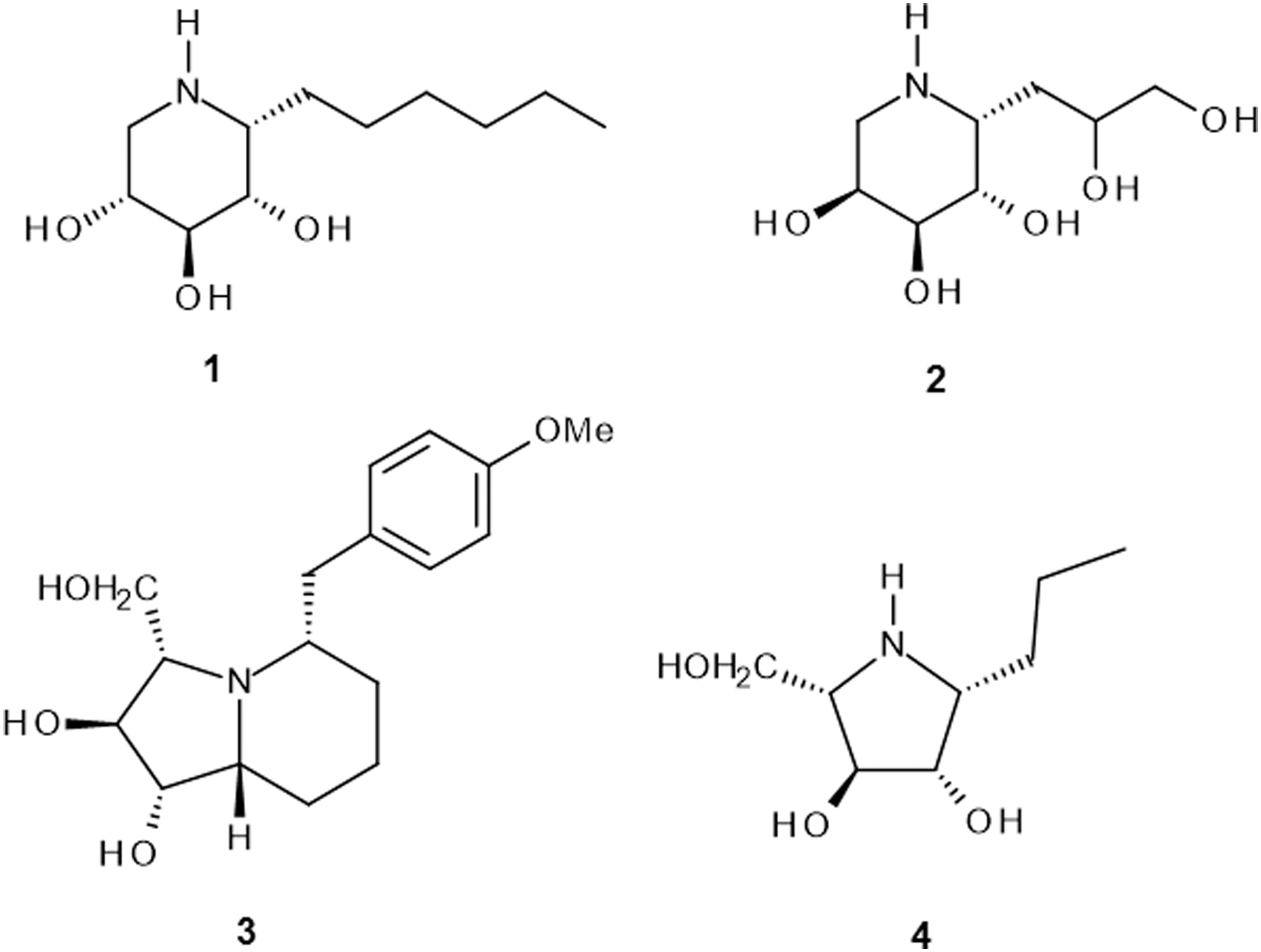
Chemical structures of the compounds:
The details of the synthesis of HDIX and DDIA have been recently published. 1 The synthesis of DHMBI has also been recently reported, 22 and PDIA was obtained by hydrogenation of the precursor of DHMBI (Fig. 2).
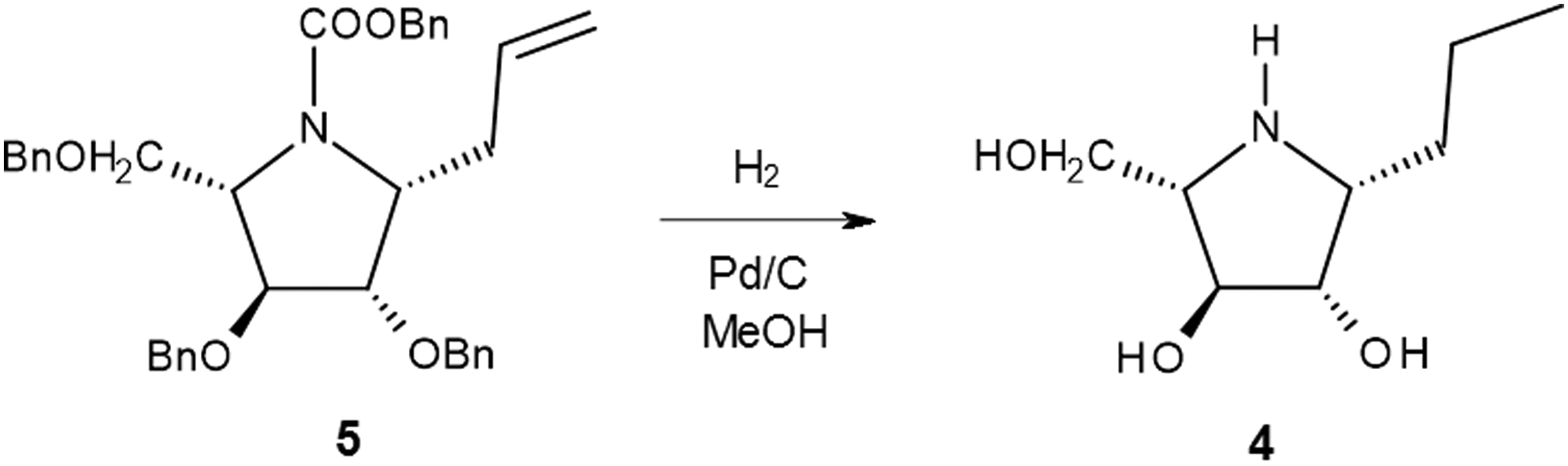
The synthesis pathway of PDIA
(1R)-PDIA: To a solution of compound 5 (302 mg, 0.52 mmol) 13 in isopropyl alcohol (iPrOH, 5 ml) were added 1 mol l−1HCl (2 ml) and 10% Pd on charcoal (78 mg). The mixture was placed under an atmosphere of hydrogen and vigorously stirred for 18 hours; the catalyst was then removed by filtration through a membrane and washed with iPrOH, and the solution was concentrated under reduced pressure. The crude product was dissolved in water (5 ml) and treated with Amberlite IRA-400 (OH−) ion-exchange resin; the resin was filtered, and the solvent evaporated to provide homogeneous PDIA (79 mg, 86%). Data for PDIA: Rf = 0.12 (petroleum ether: ethyl acetate 7: 3); [α]D −30.1 (c 0.92, MeOH). 1 H NMR (Bruker Avance II NMR spectrometer, 400 MHz, CD3OD): δ 3.84 (d, J = 3.3 Hz, 1H, H3), 3.77 (d, J = 3.6 Hz, 1H, H2), 3.67 (d, J = 4.8 Hz, 2H, H5), 3.08 (dt, J = 3.6 Hz, 6.8 Hz, 1H, H1), 2.94–2.91 (m, 1H, H4), 1.62–1.39 (m, 4H, H6, H7), 0.97 (t, J = 7.2, 3H, H8). 13 C NMR (100 MHz, CD3OD): δ 81.53 (C3), 79.27 (C2), 68.68 (C4), 63.30 (C5), 62.65 (C1), 31.67 (C6), 21.36 (C7), 14.66 (C8). ESI-HRMS: calculated for C8H18NO3 [M+H]+: 176.12812; found: 176.12819.
Bacterial cultures
All tests were performed on three P. aeruginosa strains: PAR 5, PAR 20, and PAR 50. The strains were randomly selected from a larger collection of PAR from clinical materials diagnosed in Chair of Microbiology, Jagiellonian University Medical College in Krakow. The samples for microbiological analyses were taken from wounds of patients with chronic diabetic foot infections. The strains were propagated in 10 ml of trypticase soy broth (TSB; Difco) at 37°C for 24 hours under aerobic conditions. Then, the cultures were centrifuged (400 g for 10 min) and washed with 10 ml of saline buffer. Initial suspensions of bacteria containing 1 × 108 colony-forming units (CFU)/ml were prepared by making serial dilutions in saline using the MacFarland's scale. Then, numbers of bacteria were confirmed using a standard viable count method on MacConkey agar (Oxoid) using decimal dilutions. 23
Growth conditions and measurement of biofilm formation by P. aeruginosa
P. aeruginosa biofilm was set up in sterile plastic 96-well plates with adherent surface (Greiner Bio-One). Twenty microliter quantities of the bacterial stock suspensions, prepared as described above, were added to each well, followed by 180 μl of sterile TSB. The final concentration of the bacteria was 1 × 108 CFU/ml. The plates were centrifuged for 10 minutes at 400 g to sediment bacteria on the bottom of each well and then incubated at different times at 37°C under aerobic conditions.
Biofilm quantity was determined using Congo red dye according to a modified procedure described by Reuter et al. 24 Briefly, at different time points of the culture, the plates were centrifuged; the culture medium was gently removed from wells, and immediately, 200 μl of 0.1% Congo red solution was added. The plates were left for 30 minutes at room temperature and then 10 minutes before the end of staining; the plates were centrifuged for 10 minutes at 400 g and then washed twice with buffered saline to remove unbound dye. Absorbance was measured at λ = 492 nm wavelength using spectrophotometer (Awareness Technology, Inc.). All measurements were performed in triplicates, and mean ± standard deviation (SD) values were calculated.
Effects of the iminosugars on early and mature P. aeruginosa biofilm and numbers of P. aeruginosa cells embedded in biofilm structure
Influence of the iminosugars on Pseudomonas cells forming biofilm was observed by filling wells of a 96-well plate with 20 μl of freshly made standard suspensions of P. aeruginosa strains containing 1 × 108 CFU/ml and 180 μl of TSB broth (Oxoid), as described above. Tested substances (HDIX, DDIA, DHMBI, and PDIA) were added immediately after setting up the bacteria at a final concentration of 0.9 mM. Wells filled with bacterial suspensions only were used as a control. Then, the plates were gently rotated on a table to distribute an iminosugar and incubated for 48 hours (early biofilm) and 72 hours (mature biofilm) at 37°C in aerobic conditions.
Biofilm formation and the number of viable bacteria during early biofilm development under influence of the iminosugars or in control were checked in parallel rows of the wells in the following time intervals: 0, 24, and 48 hours. Biofilm quantity was measured in one row using the Congo red method, as described above. At the same time intervals, biofilm content in each well of the second row was removed and mixed by multiple pipetting and transferred to sterile tubes. Then, decimal dilutions of the bacterial suspension were made in TSB broth, plated on MacConkey agar (Oxoid), and incubated at 37°C for 24 hours. Numbers of colonies grown on plates were counted, and total numbers of the viable bacteria were calculated as CFU/ml.
To determine inhibitory properties of iminosugars against mature biofilm, the substances were added 48 hours after adherence of bacteria to the wells in a 96-well plate. The growth conditions of P. aeruginosa were exactly the same, as described above. After the addition of iminosugars, the biofilm formation and the number of viable bacteria were checked in the following time intervals: 48, 48 + 8, and 48 + 24 hours. Biofilm quantity and numbers of viable Pseudomonas cells embedded in mature biofilm were estimated as described above. All measurements were performed in triplicates, and mean ± SD values were calculated.
Detection of apoptosis and necrosis caused by tested compounds by fluorescence microscopy
The Annexin-V-FLUOS Staining Kit (Roche) was used for the detection of apoptotic and necrotic cells according to the manufacturer's protocol. This kit uses a dual-staining protocol, in which the apoptotic cells are stained with annexin-V (green fluorescence), and the necrotic cells are stained with propidium iodide (PI; red fluorescence). Caco-2 cells (human intestinal epithelial cell line) were grown for 24 hours in Dulbecco's Modified Eagle Medium (Life Technologies) supplemented with fetal bovine serum (Sigma-Aldrich) (10% CO2, 37°C, and 90% humidity) to about 70% confluence. Then, the cells were treated with tested iminosugars with a final concentration of 0.9 mM for 24 hours. At the beginning of the experiment and after 8 and 24 hours of incubation, fluorescence was observed under fluorescence microscope (BX51, Olympus). The cells were viewed under magnification of 400× using filters: TRITC (tetramethylrhodamine) intended for viewing necrotic cells (550–570 nm) and FITC (fluorescein isothiocyanate) intended for viewing apoptotic cells (495–519 nm).
Statistical analyses
Statistical analyses were performed using STATISTICA version 10 software. Statistically significant values of the group means for three strains of PAR were evaluated using Student's t-test. The statistical analysis performance was considered significant when p < 0.05 and highly significant when p < 0.01. Since significant differences among the tested strains for their susceptibility to iminosugars activity to their biofilm formation were observed, activities of these substances versus control were calculated for mean values for all tested strains.
Results
While examining the effect of the compounds on biofilm quantity produced by three P. aeruginosa strains PAR 5, PAR 20, and PAR 50, we observed differences between two phases of biofilm formation. When iminosugars were added directly to suspensions of P. aeruginosa, which were then allowed to sediment, a distinct inhibition of the early biofilm development was observed for all tested compounds, although at different degrees (Fig. 3).
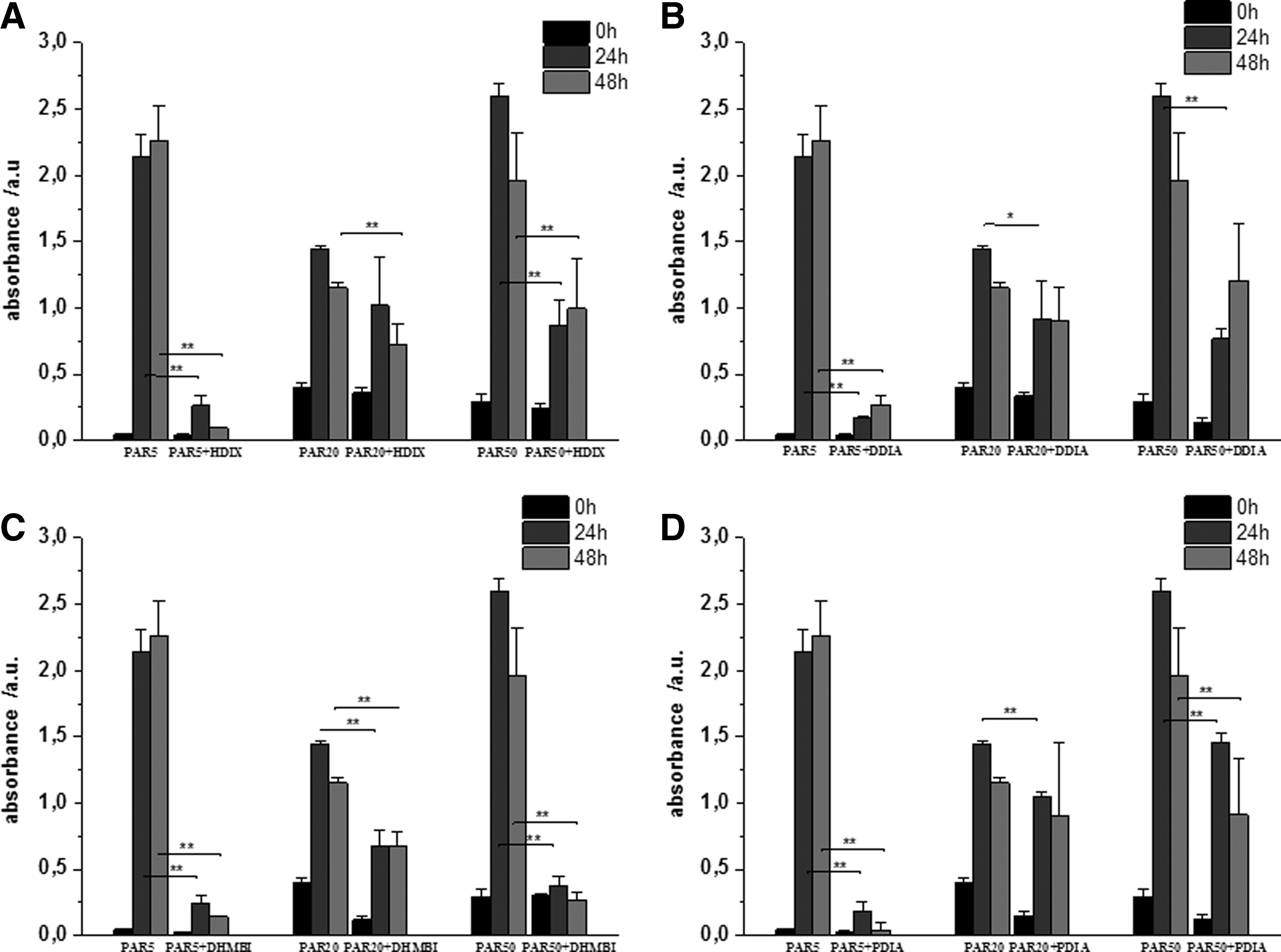
Effect of the tested iminosugars on PAR cultures in early phase of biofilm formation.
It appears also that the tested strains of P. aeruginosa, cultured without the iminosugar, differed in the production of the biofilm. The most active were the strains PAR 50 and PAR 5, which after 24 hours liberated the largest quantities of biofilm expressed as absorbance equal to 2.6 and 2.15, respectively, while the absorbance value of strain PAR 20 was only 1.45.
The compound DHMBI appeared to be the most active, followed by HDIX and PDIA, while DDIA was less inhibitory. Moreover, the latter compound was able to inhibit biofilm production for only 24 hours of incubation.
In addition, such iminosugars, HDX, DHMBI, and PDIA, showed the most potent antibiofilm activity, particularly to such strains of P. aeruginosa (PAR 5 and PAR 50), in which, after 24 hours, the rate of formation of EPS was the quickest.
Generally, the biofilm produced by PAR 5 strain was more strongly inhibited than those produced by strain PAR 50 and then by strain PAR 20.
In contrast, no significant effect on changes in the viable bacterial numbers was observed during 48 hours of incubation of the tested PAR with all investigated iminosugars (Fig. 4). There were also no significant differences between the tested strains.
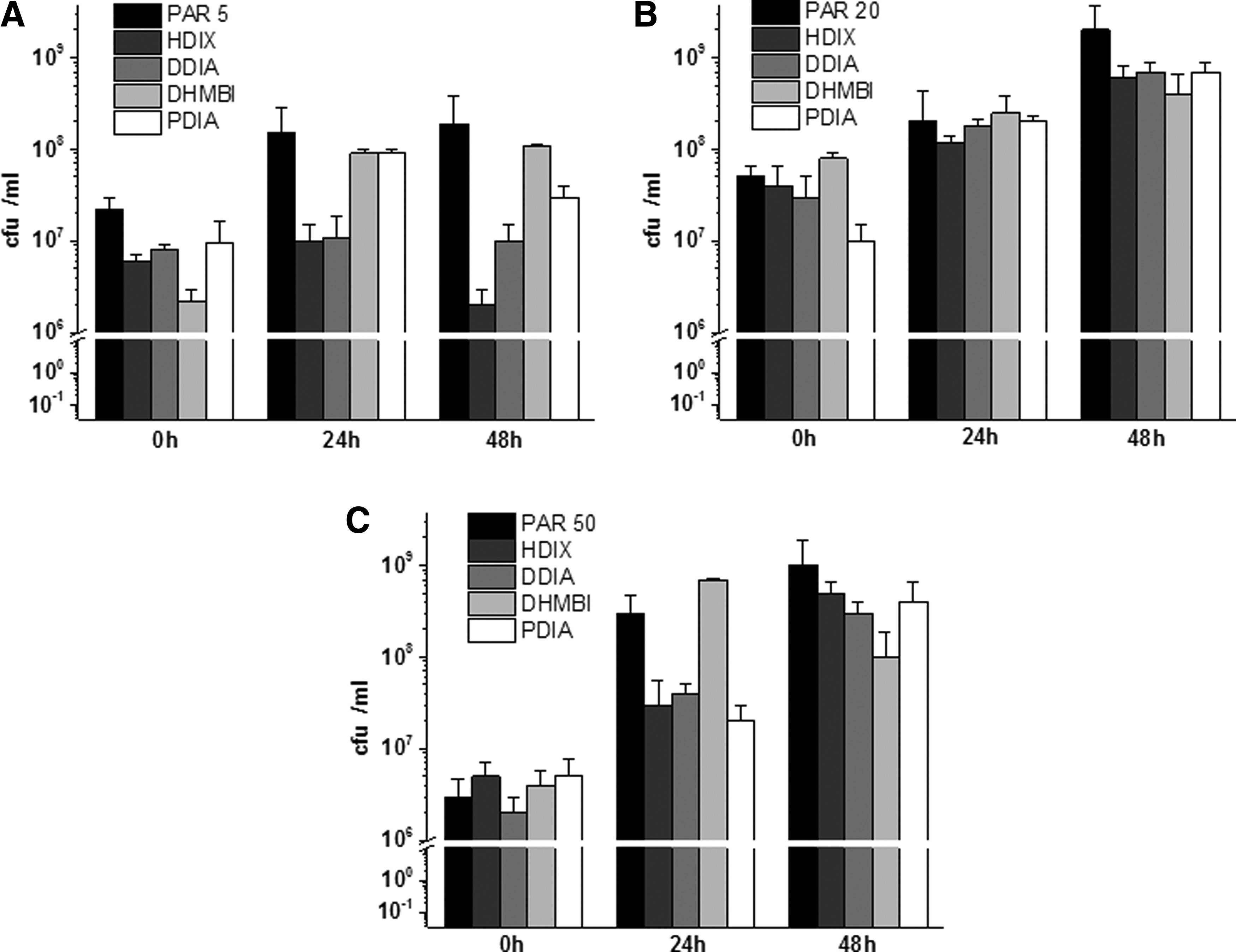
Effect of the tested compounds on numbers of viable Pseudomonas aeruginosa populations in early biofilm phase formation. Results depict logs of CFU/ml for three P. aeruginosa strains
A relatively weak influence on biofilm formation was exerted by the iminosugars added to mature biofilm of all tested PAR strains (Fig. 5). The studied compounds were able to inhibit production of P. aeruginosa biofilm only up to 8 hours of incubation with already formed biofilm or decompose its already formed structure. The biofilm thickness measured as absorbance after Congo red staining was almost identical when the strains were incubated for 48 + 24 hours in the presence of the tested substances or without them. What is noteworthy, only compound HDIX showed higher absorbance after staining with Congo red, that is, caused by the increased biofilm amount before the end of the experiment. This is in opposition to three remaining iminosugars for which a slight decrease in the biofilm quantity was observed (Fig. 5)

Effects of the tested iminosugar compounds on mature P. aeruginosa biofilm:
No significant effect of the substances was observed when numbers of viable PAR cells were measured in mature biofilm (Fig. 6). Although some differences were noted, for example, growth of PAR5 and PAR20 strains was slightly inhibited (<1 log reduction) by some compounds, these were not significant.
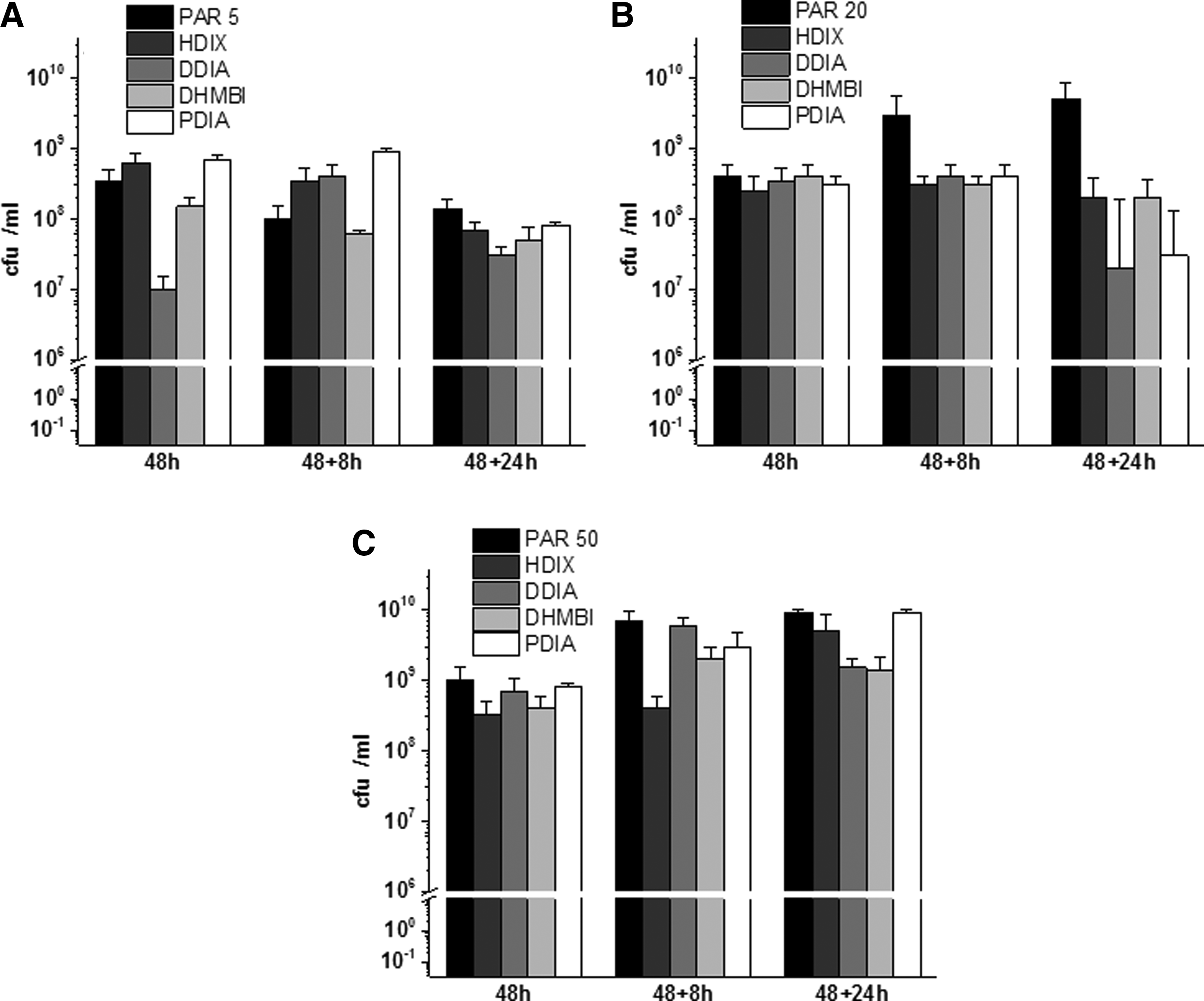
Effect of the compounds on numbers of viable P. aeruginosa populations in 48-hour-old P. aeruginosa mature biofilm. Results depict logs of CFU/ml for three P. aeruginosa strains
Except for investigating interactions of the iminosugar compounds with P. aeruginosa bacteria and biofilm formation, they were checked for their effects on human intestinal epithelium cells of the Caco-2 line. A discrete necrosis but not apoptosis was observed. As can be seen in Figure 7, cells stained entirely in red represent the necrotic cells after disruption of the cell membrane, while those stained in blue represent undamaged cells.
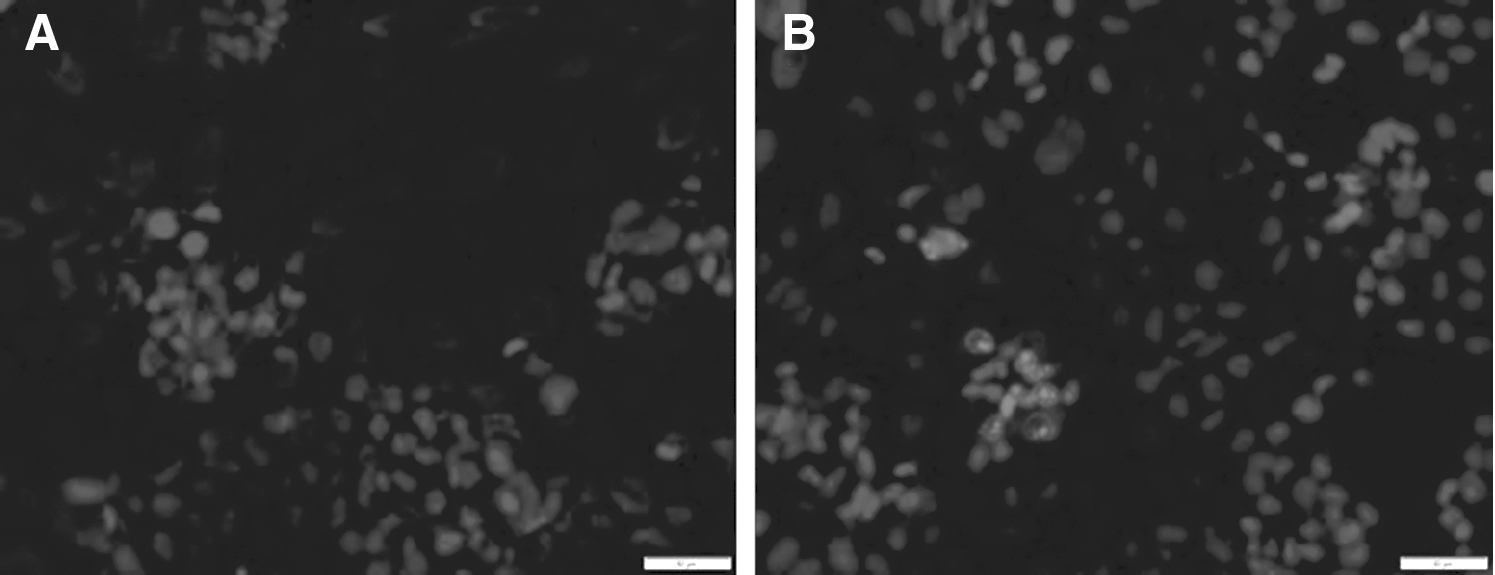
The percentage of live, necrotic, and apoptotic cells after 24 hours of incubation with compounds (0.9 mM) is presented in Table 1.
DDIA, β-1-C-(2,3-dihydroxypropyl)-1,5-dideoxy-1,5-imino-L-arabinitol; DHMBI, (1S,2S,3S,5S,9R)-1,2-dihydroxy-3-hydroxymethyl-5-p-methoxybenzylindolizidine; DMEM, Dulbecco's Modified Eagle Medium; FBS, fetal bovine serum; HDIX, α-1-C-hexyl-1,5-dideoxy-1,5-imino-D-xylitol; PDIA, 1-C-propyl-1,4-dideoxy-1,4-imino-L-arabinitol.
Discussion
P. aeruginosa is a well-recognized opportunistic pathogen responsible for various infections, from acute pyogenic soft tissue infections and life-threatening ventilator-related pneumonias in intubated patients and cystic fibrosis patients up to chronic wound infections in diabetic patients. All these infections are related to formation of biofilm by the pathogen. Since P. aeruginosa cells embedded in biofilm matrix are much more resistant to antibiotics, disinfectants, and surfactants than planktonic cells, treatment of P. aeruginosa infections is extremely difficult and often unsuccessful.25,26 Therefore, many efforts have been made to prevent P. aeruginosa biofilm formation in vitro and in vivo. Most of the studies on inhibition of biofilm formation by P. aeruginosa concentrated on substances, which might interact with cell-to-cell signaling (quorum sensing), or gene regulation of biofilm synthesizing enzymes.27,28
The presented approach was based on chemical compounds that might act as inhibitors of enzymes and/or receptors involved in the biosynthesis of EPS. Extremely important is the fact that iminosugars tested here are not present in human cells and that they may be considered as carbohydrate mimics. Apparently, since iminosugars are known to interact strongly with various cellular processes by inhibition of cellular glycosidases and glycosyltransferases, they may also interfere with bacterial biofilm formation. 29 Accordingly, iminosugars may be suitable compounds to interfere with the biosynthesis of EPS.
Our results suggest that, indeed, it is possible to inhibit synthesis of the biofilm at an early stage using new iminosugars, although there are differences in this activity among four compounds studied as well as differences among individual P. aeruginosa in their sensitivity to this activity displayed by the same compound. This sensitivity may be related to the amount and chemical composition of the biofilm produced by individual strains. In contrast, the iminosugars have shown a slight influence on mature biofilm (Fig. 5), which may also indicate their mechanism of action based on inhibition of exopolysaccharide backbone synthesis only in the early stages of biofilm formation. It is of interest that only one of the compounds (HDIX) showed their ability to promote biofilm synthesis in the late phase of the bacterial growth (Fig. 5). Since P. aeruginosa biofilm thickness measured at the same time was declining in the control well even without any chemical added, it may indicate that iminosugars added served as a source of nutrients for the bacteria. Alternatively, it is possible that the compound exerted its activity on 3D structure of the late biofilm in 24 hours, as shown on SEM pictures, which corresponded with increased optical density. It is also of importance that the compounds do not inhibit the growth of the bacteria forming the biofilm (Figs. 4 and 6). Our data suggest that the iminosugars interfere only with biofilm biosynthesis but not interact with reproduction cycles of the tested P. aeruginosa strains. Moreover, the compounds did not cause cytotoxicity toward the tested human cell line (Fig. 7 and Table 1).
Currently, it seems that synthesis and selection of the appropriate iminosugars will bring new perspectives into development of compounds, which will be able to effectively inhibit the formation of the successive stages of the exopolysaccharide chains and in consequence may increase the permeability of the biofilm to antibiotics, disinfectants, or nanocompounds. These iminosugars may interfere not only with the transferases but also with other carbohydrate-active enzymes. Undoubtedly, more studies are needed to elucidate the molecular mechanism of the inhibitory activity of the iminosugars and define specificity and antibiofilm spectrum of the compounds against a broader range of bacterial species.
In summary, it appeared that all iminosugars were able to cause a decrease in P. aeruginosa biofilm formation in the early stage of its biosynthesis. This may indicate that synthesis of exopolysaccharides plays an important role in early biofilm formation by this pathogen. Moreover, it is plausible that inhibition of glucosyltransferases and other enzymes, which are involved in exopolysaccharide synthesis, has a direct impact on P. aeruginosa biofilm formation and development.
On the basis of the presented results, it may be speculated that, showed here for the first time, iminosugars inhibiting early biofilm formation will appear helpful in preventing P. aeruginosa biofilm formation in case of the infected tissues or on the surfaces of the contaminated implants. Successful reduction or inhibition of biofilm formation will potentiate antibacterial activity of the antibiotics and other antibacterials.
Footnotes
Acknowledgments
This work was supported by the grant N N401 547040(2012–2014) and statutory grant K/ZDS/002861 (2012).
Disclosure Statement
No competing financial interests exist.