
Abstract
We investigated the epidemiology and clonality of 175 nonrepetitive methicillin-resistant Staphylococcus aureus (MRSA) isolates from clinical specimens collected between 2011 and 2012 in Kinta Valley in Malaysia. Molecular tools such as polymerase chain reaction, pulsed-field gel electrophoresis, and staphylococcal protein A (spa) typing were used. Our study revealed the predominance of three closely related ermA+ SCCmec type III pulsotypes belonging to spa type t037 (Brazilian–Hungarian clone), which were deficient in the locus F, but positive for the ccrC gene in majority (65.7%) of the MRSA infections in this region. The first evidence of SCCmec type II MRSA in the country, belonging to spa type t2460, was also noted. Although the carriage of pvl gene was uncommon (8.6%) and mostly confined to either SCCmec type IV or SCCmec type V isolates, most of these isolates belonged to spa types t345 or t657, which are associated with the Bengal-Bay CA-MRSA clone. Interestingly, spa t304 and t690 SCCmec type IV pvl+ were also detected among the MRSA isolates. Data from this study show the rise of uncommon clones among MRSA isolates in Malaysia.
Introduction
S
The prevalence of MRSA remains high in East Asia. It is one of the most frequently isolated opportunistic bacteria in clinical settings, but epidemiological data on the genetic background of MRSA in many parts of South East Asia, including Malaysia, remain elusive.4,5 Multiple studies have shown that a limited number of MRSA clones are in circulation in various parts of the world with the major pandemic MRSA clones being the Iberian, Brazilian–Hungarian, New York/Japan, and pediatric clones. 6
Furthermore, the emergence of community-acquired MRSA in the last two decades threatens to elevate the status of MRSA from an opportunistic pathogen to an active pathogen.7,8 Of interest is the spread of Panton–Valentine Leukocidin (pvl)-positive strains. PVL is an exotoxin specific to S. aureus belonging to the bicomponent synergohymenotropic toxins that kill leukocytes by creating pores in the cell membrane. 9 Carriage of pvl and SCCmec type IV has been linked to community-acquired MRSA infections, including primary skin infections and severe necrotizing pneumonia.10,11
In addition, macrolide and lincosamide resistance was reported to be high among staphylococci. Macrolide resistance among staphylococci is mainly attributed to target modification by methylase enzymes (Erm) and less commonly to efflux (Msr). 12 While the prevalence of these genetic elements is well studied, the relationship of the carriage of these genetic elements with specific MRSA genetic lineages remains largely unexplored. In light of this, the present study was undertaken to investigate the clonal relatedness and the genetic background of the MRSA isolates in Kinta Valley, Malaysia, in conjunction with the distribution of several genetic determinants, including the macrolide resistance genes (ermA, ermB, ermC, msrA, msrB) and the pvl gene.
Materials and Methods
Bacterial strains
One hundred seventy-five MRSA isolates from different patients were obtained from Raja Permaisuri Bainun Hospital, the state's referral hospital (142/175), KPJ Ipoh Specialist Hospital, a leading private referral hospital (28/175), and Gribbles Pathology Ipoh (5/175) in Kinta Valley, Malaysia, from 2011 to 2012. Species confirmation of S. aureus and screening of oxacillin resistance were performed by the individual participating hospitals. A total of six isolates were from confirmed cases of community acquired MRSA infection. The six samples were isolated from skin lesions of healthy individuals in the community with no catheterization. Classification as CA-MRSA is based on fulfillment of several criteria outlined by the CDCs Active Bacterial Core Surveillance Program, including isolation from outpatient or hospitalized patients within 48 hours of hospitalization and the lack of well-established healthcare-associated MRSA risk factors: hemodialysis, surgery, as well as residence in long-term care facility. 13
Total DNA extraction
A volume of 2.4 ml of the overnight bacterial suspension was pelleted by centrifugation at 13,000 rpm for 5 minutes, resuspended in 180 μl sterile distilled water, and subjected to treatment with 20 μl of 50 mg/ml lysozyme chloride (Nacalai Tesque) at 37°C for 1.5 hours. This was followed by addition of three volumes of 6 M guanidine hydrochloride (Merck) before incubation at 65°C for 1 hour. The cell debris was pelleted by centrifugation at 13,000 rpm for 5 minutes. Supernatant was transferred to a fresh microcentrifuge tube and 0.6 volume of isopropanol (Merck) was added to it. The mixture was then transferred into a silica minicolumn (Epoch Life Science). A series of washes were performed, and the DNA was eluted with sterile distilled water. The eluate was used as template DNA for polymerase chain reaction (PCR) or stored at −20°C until required.
Susceptibility testing
The MRSA isolates were tested against a panel of 10 antibiotics, including oxacillin (1 μg), penicillin (10 U), erythromycin (15 μg), clindamycin (2 μg), gentamicin (10 μg), trimethoprim–sulfamethoxazole (25 μg), vancomycin (30 μg), rifampicin (5 μg), fusidic acid (10 μg), and linezolid (30 μg), using the Kirby–Bauer disk diffusion test and interpreted based on the criteria outlined by the Clinical and Laboratory Standards Institute. 14
PCRs detection of target genes
SCCmec typing of the isolates was performed using a multiplex PCR assay described by Milheirico et al. 15 The SCCmec types identified in the study were further validated by determining the mec class and ccr complexes carried by representative strains of each of the amplification patterns obtained. The determination of mec class and detection of ccrC gene were accomplished using the primers described by Chen et al., 16 and ccrAB allotyping was performed using the PCR assay described by Boyle-Vavra et al. 17 The results were interpreted using the guidelines drawn by the International Working Group on the Staphylococcal Cassette Chromosome elements (IWG-SCC). 18 PCR detection of ermA, ermB, ermC, msrA, msrB, and pvl gene was carried out using the primer sequences and PCR cycling conditions described previously.10,12 For each pair of the primers, PCR was performed in a total reaction volume of 25 μl consisting of the following: 1X PCR Green GoTaq® Buffer (Promega), 1.5 μM MgCl2 (Promega), 0.2 mM dNTPs (Promega), 0.4 μM of each primer, 1.25 U Taq Polymerase (Promega), and 100 ng of template DNA. The amplicons were resolved in agarose gel and visualized under the UV transilluminator.
Pulse-field gel electrophoresis
Chromosomal DNA plug preparation and SmaI restriction digest were performed using the protocol recommended by PulseNet USA. 19 Pulse-field gel electrophoresis (PFGE) of the SmaI (New England Biolabs) digested chromosomal DNA of MRSA isolates was performed in the CHEF Mapper® XA PFGE System (BioRad) using the following parameters: Voltage of 200 V, temperature of 14°C, initial switch time of 5 seconds, final switch time of 40 seconds, and a total run time of 20 hours and 30 minutes. The gels were stained with GelRed (Biotium) and photographed under UV light. The Dice coefficient was used to calculate the similarities, and the unweighted pair group with arithmetic mean (UPGMA) was used for cluster analysis with BioNumerics software version 7.1 (Applied Maths). A tolerance setting of 1.8% and optimization of 1.5% were used for analysis. The isolates were grouped into the same clusters if they share 80% or more similarity.
Staphylococcal Protein A typing
Clonal spreading is defined as the isolation of the MRSA isolates demonstrating a particular pulsotype in multiple patients. 20 In this study, one representative sample of each pulsotype that demonstrated clonal dissemination as well as the sole SCCmec type II MRSA isolate was selected for staphylococcal protein A (spa) typing using the primer sequences described previously. 21 The sequences were queried using the spaType finder software available on http://spatyper.fortinbras.us
Results
Antibiogram
All 175 MRSA isolates were resistant to oxacillin (100.0%, 175/175) and penicillin (100.0%, 175/175). Most of the isolates were also resistant to erythromycin (93.7%, 164/175), clindamycin (88.0%, 154/175), trimethoprim–sulfamethoxazole (81.1%, 142/175), and gentamicin (88.0%, 154/175). The MRSA isolates, however, demonstrated low or no resistance toward the following antibiotics: rifampicin (3.4%, 6/175), fusidic acid (5.7%, 10/175), vancomycin (0.0%), and linezolid (0.0%).
Prevalence of erm and msr genes
Macrolide resistance was mainly mediated by the erm genes. Out of the 175 isolates, 143 (81.7%) carried the ermA gene and 17 (9.7%) carried the ermC (Table 1). None of the isolates were found to be carrying ermB. The carriage of msr genes (msrA and msrB) were infrequent. Only 9 (5.1%) isolates harbored the msr genes. Eight of the nine msr+ isolates carried both msrA and msrB genes, while only one carried msrA gene. The majority of ermA+ (96.5%, 138/143) and ermC+ (82.4%, 14/17) isolates were resistant to both erythromycin and clindamycin. In contrast, only one msr+ isolate (11.1%, 1/9) was resistant to both of the antibiotics, showing that msr+ isolates were less likely to be resistant to both erythromycin and clindamycin. Four msr+ isolates (44.4%, 4/9) demonstrated resistance toward erythromycin, but not clindamycin, while another four (44.4%, 4/9) were sensitive to both erythromycin and clindamycin.

Distribution of pvl genes
Fifteen (8.6%) of the 175 MRSA isolates were found to carry the pvl gene (Table 1). The carriage of pvl gene was SCCmec types specific (Table 1). Out of the 15 pvl+ strains, eight (53.3%) were isolated from skin lesions. All six confirmed cases of CA-MRSA infections were attributed to pvl+ strains.
Prevalence of SCCmec types
The predominant SCCmec type was SCCmec type III (81.1%, 142/175) followed by SCCmec type IV (12.6%, 22/175), and SCCmec type II (0.6%, 1/175). Ten MRSA isolates (5.7%, 10/175) were positive only for ccrC gene and mecA gene, which were suggestive of SCCmec type V. The SCCmec Type III carried by the MRSA isolates was deficient in the locus F (414 bp) and positive for the ccrC gene. The amplification patterns of representative isolates are shown in Fig. 1. The identities of all SCCmec types detected in this study were verified by the determination of their respective mec classes and ccr complexes. The carriage of mec class A-ccr gene complex 2 and mec class C2-ccr complex 5 confirmed the identities of SCCmec type III and SCCmec type V, respectively (data not shown).

SCCmec typing of MRSA by multiplex PCR and analysis by 3.0% agarose gel electrophoresis. Lanes M, VC 100 bp Plus DNA Ladder (Vivantis); Lane 1, Type II; Lane 2, Type IV; Lane 3, Type IV (atypical); Lane 4, Type IV; Lane 5, Type IV; Lane 6, Type V; Lane 7, Type III; Lane 8, Type III (atypical); Lane 9, negative control. MRSA, methicillin-resistant Staphylococcus aureus; PCR, polymerase chain reaction.
Clonal relatedness of MRSA isolates
PFGE segregated the 175 MRSA isolates into four major clusters based on 80% similarity; Clusters A (82.9%, 145/175), B (5.7%, 10/175), C (4.6%, 8/175), and D (6.9%, 12/175) (Table 1; Figs. 2 and 3). A total of 41 distinct MRSA pulsotypes (strains) were identified in this study. Of the 41 strains, 13 demonstrated clonal spreading. Three closely related pulsotypes belonging to Cluster A; Pulsotypes A3 (24.6%, 43/175), A15 (12.0%, 21/175), and A16 (29.1%, 51/175) were responsible for the majority of MRSA infections.

Dendogram generated using the unweighted pair group method with arithmetic mean with a tolerance setting of 1.8% and optimization of 1.5% with a cutoff point of 80% similarity for clustering. Only representative isolates for pulsotypes A3, A15, and A16 are shown in the dendogram to improve its clarity. UPGMA, unweighted pair group method with arithmetic mean.
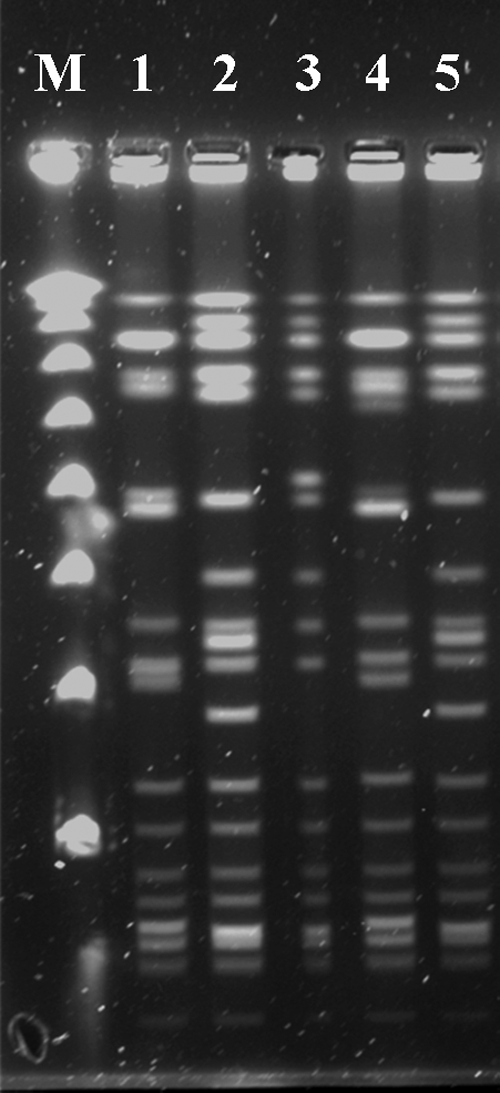
Representative PFGE gel image. Lane M, λ PFGE marker (New England Biolabs); Lane 1, Pulsotype A16; Lanes 2 and 5; Pulsotype A3; Lane 3, Pulsotype A5; Lane 4, Pulsotype A15. PFGE, pulsed-field gel electrophoresis.
In terms of the distribution of SCCmec types, Cluster A isolates were predominantly SCCmec type III (97.9%, 142/145), cluster B isolates were predominantly SCCmec type V (90.0%, 9/10), while clusters C and D were made up exclusively of SCCmec type IV MRSA isolates. The sole SCCmec type II detected in this study was classified under Cluster B and carried spa t2460 (Tables 1 and 2).
In general, there was a good concordance between the PFGE data and those of spa typing and SCCmec typing (Table 2). Spa typing revealed that all five major pulsotypes in cluster A, which demonstrated clonal dissemination, including pulsotypes A3, A15, and A16, carried closely related spa types that are associated with the Brazilian–Hungarian clone (t037 and t234). Pulsotypes B3 (1.7%, 3/175) and B4 (2.3%, 4/175) of PFGE cluster B carried spa types t657 and t345, respectively, which are associated with the Bengal-Bay MRSA clones. Interestingly, four out of the six confirmed community-acquired MRSA isolates included in this study belonged to pulsotypes B3 and B4. The remaining two CA-MRSA cases were mediated by isolates belonging to pulsotypes C3 and C5. Pulsotypes C3 (1.1%, 2/175) and C5 (1.7%, 3/175) of cluster C belonged to unrelated spa types (t304 and t690), despite being classified under the same cluster. Pulsotypes D1 (1.7%, 3/175), D3 (2.3%, 4/175), D4 (1.1%, 2/175), and D5 (1.1%, 2/175) of cluster D belonged to the same spa type t032, which is associated with the UK EMRSA-15 clone (Tables 1 and 2).
The carriage of the macrolide resistance genes and pvl gene was closely associated with the SCCmec types. All SCCmec type II and SCCmec type III isolates harbored the ermA gene. In contrast, ermC gene was only detected among the SCCmec type IV isolates (Table 2). Out of the 22 SCCmec type IV isolates, 17 (77.3%) carried ermC gene. The msr genes, in contrast, were strongly associated with SCCmec type V isolates. Out of the 10 SCCmec type V, eight (80.0%) carried both msrA and msrB genes, while one (10.0%) carried msrA gene. Similar associations were observed for pvl gene. The carriage pvl gene was only confined to SCCmec type IV and SCCmec type V isolates, particularly those that belonged to PFGE clusters B (90.0%, 9/10) and C (75.0%, 6/8).
Discussion
In agreement with previous studies, low rates of resistance toward rifampicin, fusidic acid, vancomycin, and linezolid were observed, which could be attributed to the low prescription rates of these drugs since vancomycin and linezolid are only prescribed as the last resort.22–24 In addition, rifampicin and fusidic acid are often prescribed in tandem, thus increasing the barrier for resistance development. 25
Widespread clonal dissemination of several spa type t037 MRSA seen in this study, inferred to be the Brazilian–Hungarian clone, is in line with several past epidemiological studies conducted in Malaysia and parts of Asia.26–29 In addition, these clones carried a unique SCCmec type III characterized by ccrC positivity and deficiency in locus F. Locus F (414 bp) is usually present in the prototypical SCCmec type III MRSA and is located in the region between Tn554 and the chromosomal right junction (orfx). 15 SCCmec type III positive for ccrC is by far the most prevalent form of SCCmec in several South East Asian countries, including Singapore, Indonesia, Thailand, Vietnam, and the Phillipines. 29 Interestingly, the lack of locus F in SCCmec type III has been reported among MRSA isolates belonging to the Brazilian–Hungarian in Lublin, Poland. 30
Clonal dissemination of several minor clones was also noted in this study, although the extent of their spread was markedly lower. Of particular interest is the spread of pvl+ strains. The pvl gene together with SCCmec Type IV, V, and VII has been linked to community-acquired MRSA. 31 Several pvl+ MRSA isolates, including four CA-MRSA isolates, carried spa types (t657 and t345) consistent with those of SCCmec type V Bengal-Bay CA-MRSA clone, which is widespread in India. 32 The emergence of Bengal-Bay CA-MRSA in Malaysia has also been documented in other hospitals in Malaysia.28,33 In this study, these strains were associated with community-acquired MRSA skin infections. Apart from the Bengal-Bay MRSA clone, the emergences of spa t304 and t690 SCCmec type IV pvl+ MRSA isolates (Pulsotypes C3 and C5) were also observed. Spa t304 MRSA is prevalent in the Caribbean, South Korea, and in certain parts of Europe, while spa t690 MRSA is mostly prevalent in several Middle Eastern countries like Kuwait.34,35
In addition, this study is the first to report the existence of SCCmec type II MRSA in Malaysia. The SCCmec type II isolate was obtained from a surgical wound of a 49-year-old female patient and it was resistant to oxacillin, erythromycin, clindamycin, and gentamicin, but sensitive to trimethoprim–sulfamethoxazole, rifampicin, linezolid, and vancomycin. This particular isolate belongs to spa type t2460 and is endemic in South Korea, which, interestingly, had replaced spa type t037 in intensive care units in Seoul University Hospital. 36 Thus, its emergence in this geographical area should not be taken lightly.
The existence of MRSA clones that originated from foreign countries in this geographical area highlights the role of transmigration in the dissemination of MRSA. However, despite the introduction of newer clones, those MRSA isolates belonging to spa type t037 remain the predominant strains in Malaysian hospitals.26,28,29 Numerous factors could have influenced the selection and propagation of these predominant clones. Plausible contributing factors include the combination of antimicrobial drugs used in clinical settings and the reservoirs of these clones. In addition, all four pulsotypes that demonstrated clonal dissemination in Cluster D carried spa type t032, which is associated with UK EMRSA-15 clone, which was reported in Malaysia in 2010. 26
Table 3 summarizes the distribution, pulsotypes, antibiogram, and pathological conditions of the patients by hospitals. In both of the state and private tertiary referral hospitals, the predominance of strains associated with the Brazilian–Hungarian clone over strains associated with the UK EMRSA-15 clone was elucidated, although different pulsotypes predominated in each hospital. The overwhelming success of strains associated with the Brazilian–Hungarian clone was likely multifactorial, but it is evident that they demonstrated resistance to a wider range of antibiotics compared to the UK EMRSA-15 clone. This could have conferred a selective advantage in the environment where antibiotic usage is high. An analysis of the pathological conditions of the patients revealed that invasive medical procedures appeared to be a strong risk factor for MRSA infection for both Brazilian–Hungarian and UK EMRSA-15 clones. The persistence of these clones suggests that intertransmission between patients was the major contributor to the endemicity of the Brazilian–Hungarian and UK EMRSA-15 clones in these two hospitals.
The predominant antibiotic resistance patterns and the pathological conditions are also shown. The superscripted letter “R” connotes antibiotic resistance. The percentage is presented in parentheses and is expressed as number of isolates over the total number of isolates associated with each clone isolated within each hospital.
The close association between ermA and SCCmec types II and III seen in this study was readily accounted for as it has been elucidated that ermA was part of SCCmec types II and III.2,37 In contrast, the relationship between ermC and msr genes with specific SCCmec types has not been thoroughly investigated, especially in Malaysia. In the present study, ermC gene was associated with SCCmec type IV MRSA, particularly those that carried spa t032 (UK EMRSA-15 clone) while the msr genes were associated with spa t657 and t345 (Bengal-Bay clone). While ermC had been shown to be carried on a plasmid capable of transconjugation in vitro by Lim et al., 38 the result of this study suggests that clonal dissemination plays a greater role in its spread.
Results from this study also showed that CA-MRSA clones were more likely to use an efflux pump mechanism for macrolide resistance than nosocomial MRSA strains. This observation corroborates well with the mechanistic experiment conducted by Gupta et al., 39 who showed that expression of erm genes reduced the competitive fitness of MRSA. This could explain why erm+ MRSA isolates' spread was only limited to hospital settings where antibiotic usage is high. This would have favored the selection of these strains and allowed them to occupy this ecological niche since the growth of beneficial microflora would have been suppressed.
In conclusion, three major SCCmec type III spa t037 strains (pulsotypes A3, A15, and A16) associated with the Brazilian–Hungarian clone were the predominant strains in Kinta Valley hospitals from 2011 to 2012. Sporadic cases of MRSA skin infections caused by pvl+ spa t657 and t345 isolates, associated with the Bengal-Bay MRSA clone, were also reported in the community. The first report of SCCmec type II isolate carrying spa t2460 was also presented in this study, which warrants further molecular characterization and surveillance. This study also confirmed the role of target modification of ribosome by methylase (erm) as the main macrolide resistance mechanism used by MRSA, specifically those of SCCmec types II, III, and IV.
Footnotes
Acknowledgments
This work was supported by the UTAR Research Fund (IPSR/RMC/UTARRF/C2-11/H01) from Universiti Tunku Abdul Rahman. The authors would like to thank Ms. Suganthi of RPBH and Ms. Shree of ISH for providing the bacterial strains.
Ethical Standards
This work was registered under Malaysia's National Medical Research Register, National Institute of Health, with the project number NMRR ID: 11-436-9556. Prior approval for the collection of MRSA clinical isolates and patient data was also obtained from the Medical Research and Ethics Committee (MREC) of the Ministry of Health, Malaysia.
Disclosure Statement
W.Y.H. is the graduate researcher who carried out the research, wrote the manuscript, and analyzed and discussed the results. Q.C.C. plays the cosupervisory role and was involved in the manuscript editing. C.H.C. plays the main supervisory role and was heavily involved in the conception, planning, and supervising the execution of the work to the final draft of this article.