
Abstract
Tigecycline is one of the few therapeutic options that are available for treating serious clinical infections. However, tigecycline nonsusceptible Enterobacteriaceae has emerged recently in China. In this study, a total of 28 clinical Klebsiella pneumoniae isolates that were not previously exposed to tigecycline were collected and confirmed for tigecycline minimum inhibitory concentrations (MICs) using standard broth microdilution tests. To elucidate the mechanisms underlying molecular resistance to tigecycline, the expression levels of efflux pumps AcrAB and OqxAB and their regulators RamA, MarA, RarA, and SoxS were determined by quantitative polymerase chain reaction. The expression levels of the genes acrB, ramA, marA, and soxS were statistically different in different MIC groups (p < 0.05). Sequence analysis of the acrR and ramR genes revealed several nonsynonymous mutations in the nine resistance isolates. The values of MIC in these isolated strains with ramR mutations were significantly higher than those without ramR mutation (p = 0.029). Moreover, mutations in the ramR gene led to the overexpression of RamA. These results indicated that the mutation of the ramR gene through the upregulated expression of RamA contributed to tigecycline resistance and that several of the newly identified types of mutations in ramR and acrR were not previously reported in K. pneumoniae clinical isolates.
Introduction
T
The mechanisms underlying tigecycline resistance are not well understood. Previous studies demonstrated that the reduced susceptibility observed in Enterobacter spp. may be due to the overexpression of resistance-nodulation-division (RND) family multidrug efflux pumps, such as AcrAB-TolC.7,10 OqxAB and KpgABC, which are recently identified efflux pump systems in K. pneumoniae, may represent supplementary nonsusceptibility mechanisms for RND-type efflux pump.11,12
The expression of AcrAB-TolC efflux pump is activated by several AraC-type transcriptional regulators, such as RamA, MarA, RarA, and SoxS, which interact with the promoter in the operon region, thereby conferring low-level resistance to a variety of antibiotics and disinfectants.10,13–15 In addition, AcrAB expression is regulated by its local repressor AcrR, and mutations in these genes may participate in tigecycline resistance. ramR, marR, and soxR code for the local transcriptional repressors RamA, MarA, and SoxS, respectively. Mutations in these genes can lead to excessive transcriptional activation, resulting in the upregulation of the efflux pump in Enterobacteriaceae.16–18
In this study, we collected clinical K. pneumoniae strains isolated in patients who had not previously been exposed to tigecycline and identified their tigecycline susceptibility profiles. The purpose of this study was to investigate whether similar or other molecular resistance mechanisms existed in these strains.
Material and Methods
Bacterial strains and growth conditions
From January 2012 to January 2014, 582 nonduplicate K. pneumoniae isolates from various clinically infected samples that had not previously been exposed to tigecycline were collected at Dalian Medical University Affiliated Hospitals (First Affiliated Hospital of Dalian Medical University, Second Affiliated Hospital of Dalian Medical University, and Dalian Municipal Central Hospital Affiliated of Dalian Medical University). All isolates were identified using the VITEK-2 antibiotic susceptibility testing system (BioMerieux). E. coli ATCC 25922 and K. pneumoniae ATCC 13883 were used as reference strains. The propagation of all strains was carried out in the Luria-Bertani (LB) medium at 37°C.
Tigecycline susceptibility test
Susceptibility of the isolates was determined using the Kirby–Bauer method (tigecycline [TGC], Oxoid; breakpoint for TGC: disk diffusion zone diameters ≥19 mm susceptible, 15–18 mm intermediate, and ≤14 mm resistant) and minimum inhibitory concentrations (MICs) of tigecycline (Sigma-Aldrich). The MIC of tigecycline was analyzed using the standard broth microdilution method in accordance with the Clinical and Laboratory Standards Institute (CLSI) guidelines (MIC ≤2.0 μg/mL is susceptible, 4.0 μg/mL is intermediate, and ≥8 μg/mL is resistant). 19 E. coli ATCC 25922 was used as the control strain.
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qRT-PCR) was performed to determine the expression levels of the efflux pumps AcrAB and OqxAB and their transcriptional regulators RamA, MarA, RarA, and SoxS. The primers used for real-time PCR are listed in Table 1. First, total bacterial RNA was extracted using the Total RNA Isolation Purification Extraction Kit (Sangon). The yield and quality of RNA were determined using a NanoDrop 2000C (Thermo). cDNA was synthesized using the PrimeScript RT Reagent Kit (TaKaRa). Then, real-time PCR was performed using the SYBR® Premix Ex Taq™ II Kit (TaKaRa) on the Stratagene Mx3005P qPCR System (Agilent Technologies), with 40 cycles of 5 s at 95°C and 34 s at 60°C. The experiments were carried out in triplicate.
bp, base pair; qPCR, quantitative reversion transcription polymerase chain reaction.
The expression of each target gene was normalized to that of a housekeeping gene (16S rRNA, rrsE). The expression levels of AcrAB, OqxAB, RamA, MarA, RarA, and SoxS were determined and found to be onefold greater than that of K. pneumoniae ATCC 13883 (expression = 1), which served as the reference strain of tigecycline-susceptible strains (MIC = 0.5 μg/mL). Data were analyzed using the Agilent MxPro software based on the 2−ΔΔCt method.
Efflux inhibitory experiment of PAβN
Fresh cation-adjusted Mueller-Hinton Broth (MHB, <12 h old) both with and without 50 μM Phe-Arg-β-Naphthylamide (PAβN, Sigma-Aldrich) was prepared and inoculated with 0.5 McFarland. The effect of efflux inhibitor PAβN on the activity of the efflux pump was determined by MIC assays using the broth microdilution method. 20 The MIC values were established by conducting the experiments in triplicate. A significant inhibition effect on the efflux pump was observed because the MIC values decreased fourfold or greater in the presence of PAβN.
Identification of mutations in acrR, ramR, and rpsJ
PCR and sequence analyses were performed to identify the mutations in the acrR, ramR, and rpsJ codons of the S10 ribosomal protein gene. The gene fragments of acrR, ramR, and rpsJ in clinical K. pneumoniae were amplified and compared with the reference sequence of K. pneumoniae subsp. pneumoniae MGH 78578 (CP000647). The primers designed for PCR are listed in Table 1.
Statistical analysis
The relationship between the tigecycline MICs and gene expression levels was analyzed using the SPSS 13.0 Statistics software. The gene expression levels of different MIC groups were described using the mean and standard deviation (SD). According to the normality test, the expression levels of RamA appeared to be a normal distribution, so an analysis of variance (ANOVA) was performed. Because the expression levels of AcrA, AcrB, OqxA, OqxB, RarA, MarA, and SosX showed non-normal distributions, the Kruskal–Wallis test was adopted as the post-hoc test. The relationship between mutations of the local repressor genes ramR, acrR, and tigecycline MICs was analyzed by Student's t-test. A p-value less than 0.05 was considered statistically significant.
Results
Tigecycline susceptibility
Of the 582 K. pneumoniae isolates that were not exposed to tigecycline during the study period, only 20 isolates were not susceptible to tigecycline (nonsusceptibility rate is 3.44%). Eight ordinary clinical strains that were susceptible to tigecycline were designated as the negative control. Therefore, in total, 28 clinical K. pneumoniae isolates were examined in this study. Among these isolates, the range of tigecycline MICs was 0.50–32 μg/mL, with a MIC50 of 8 μg/mL and a MIC90 of 16 μg/mL. The tigecycline susceptibility profiles of the 28 clinical K. pneumoniae isolates are presented in Table 2.
MIC: Determined by the broth microdilution method. ATCC25922: quality control strain, TGC+PAβN: tigecycline with PAβN.
K-B, Kirby–Bauer; MIC, minimum inhibitory concentration; ND, not detected.
Gene expression and relationship with tigecycline MICs
Target gene expression analysis was performed for 28 clinical K. pneumoniae isolates and compared with K. pneumoniae ATCC 13883 (tigecycline susceptible). Real-time PCR analysis showed a uniformly high gene expression of acrB (1.48 to 257.21 fold) and ramA (1.30 to 212.48 fold) of the RamA/AcrAB pathway in seven isolates (K860, K898, K1087, K1186, K1352, K1411, K1531), whereas eight isolates (K634, K746, K803, K898, K1087, K1186, K1411, K1531) exhibited simultaneous overexpression of the genes oqxB (1.74 to 16.36 fold) and rarA (1.36 to 4.04 fold). Among them, K1186 and K1531 showed upregulation of the oqxA gene (2.30 fold, 15.18 fold). In addition, the marA and soxS genes were differentially expressed among the 28 clinical strains (Figs. 1 and 2).
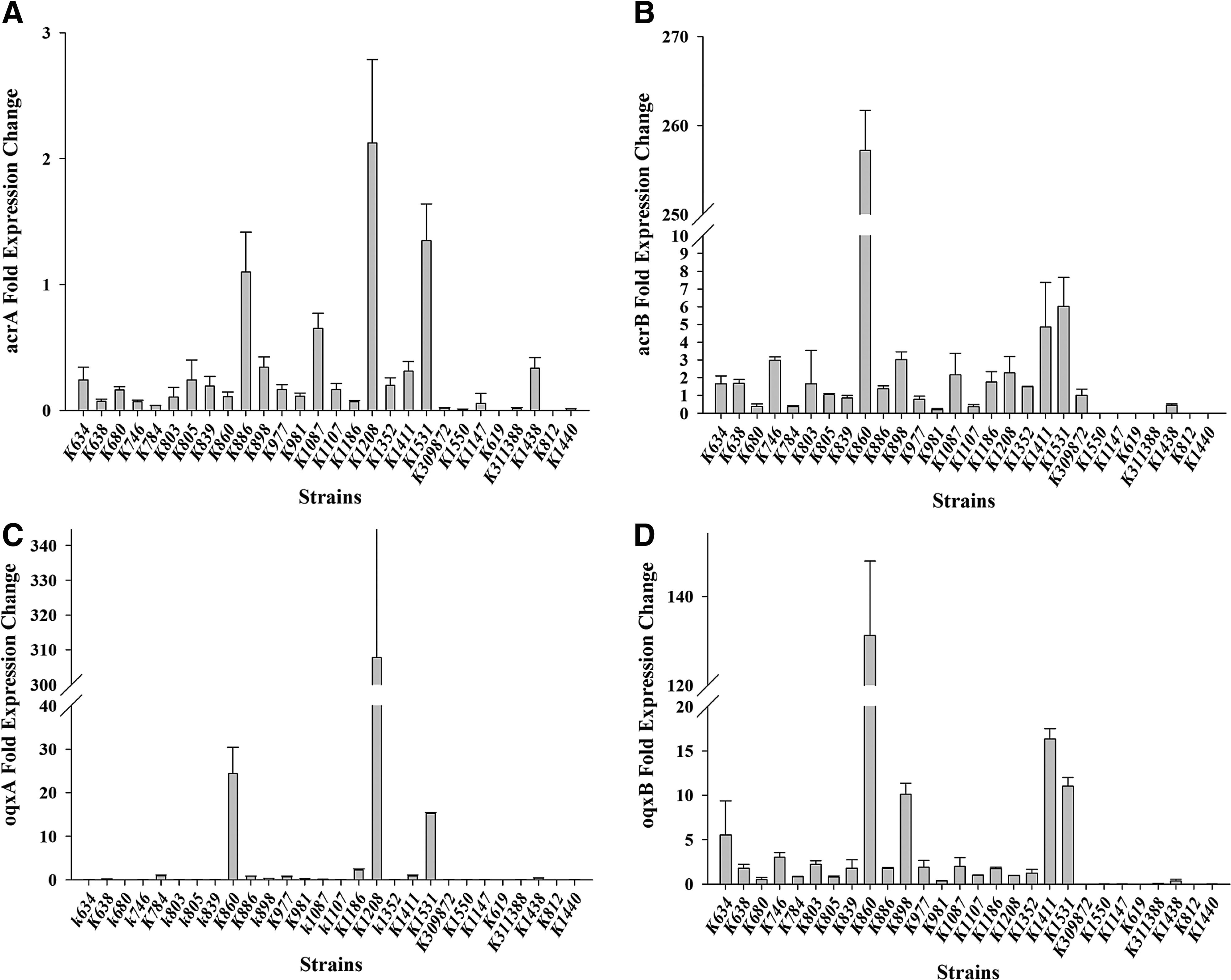
Relative fold gene expressions of efflux pumps AcrAB and OqxAB in 28 clinical Klebsiella pneumoniae isolates.

Relative fold gene expressions of transcriptional regulators RamA, RarA, MarA, and SoxS in 28 clinical K. pneumoniae isolates.
The gene expression levels of different MIC groups are presented in Table 3. According to the ANOVA results, there was a statistically significant association between ramA expression and tigecycline MICs (p = 0.033). The Kruskal–Wallis test indicated that the expression of the acrB, marA, and soxS genes was statistically significantly different in different MIC groups (p = 0.005, p = 0.016, and p = 0.023, respectively). However, the levels of acrB, marA, soxS, and ramA gene expression were not significantly different between tigecycline-resistant isolates (MIC ≥4 μg/mL) and tigecycline-susceptible isolates (MIC <4 μg/mL, p > 0.05, data not shown).
Relative expression compared with ATCC13883 (expression = 1), results are means of 3 runs ± SDs. MIC of tigecycline.
Expression levels of ramA appear to be a normal distribution, expression levels of acrA, acrB, oqxA, oqxB, rarA, marA, and sosX appear to be non-normal distribution.
p < 0.05.
SD, standard deviation.
Efflux pump inhibition assay
The tigecycline MIC values of 28 clinical K. pneumoniae isolates both with and without the efflux pump inhibitor (EPI) PAβN are presented in Table 2. Exposure to tigecycline with PAβN showed a fourfold reduction in MIC in four isolates (K638, K805, K1186, K1208), a twofold reduction in three strains (K634, K977, K981), and no change was observed in the eight tigecycline-sensitive isolates.
Mutations of ramR, acrR, and rpsJ and the relationship with tigecycline MICs
The amplified acrR, ramR, and rpsJ sequences in 28 clinical K. pneumoniae isolates were compared with the reference sequence of K. pneumoniae subsp. pneumoniae MGH 78578 (GenBank Accession No. CP000647).
Mutations in the acrR and ramR genes were observed in nine of the nonsusceptible isolates (Table 2). Mutations in the ramR gene were observed in six isolates. Two isolates (K977, K981) harbored the sequence deletion “TGATGCGCTCGC,” which led to a frameshift mutation in the ramR gene. K803 harbored four substitutions, which led to amino acid exchange in the coding regions of ramR (E175D, I177T, A178D, and A193S [protein]). The synthesis of RamR protein was prematurely terminated by a stop codon in the other three strains (K898, K860, K1186). The overexpression of ramA could have been due to the mutations of ramR in these strains. The range of tigecycline MICs among strains with ramR mutation was 8–32 μg/mL, with an MIC50 of 12 μg/mL and an MIC90 of 32 μg/mL. The MIC values in these strains with ramR mutation were significantly higher than those of other strains without ramR mutation (p = 0.029) (Table 4).
MIC of tigecycline.
p < 0.05.
Moreover, mutations in the acrR gene were observed in five resistant isolates (K638, K680, K746, K803, K860). Four isolates (K680, K746, K803, K860) harbored point mutations and a premature stop codon (G121V+E122K+M123Stop), which led to amino acid exchanges in the coding region of acrR. One isolate (K638) harbored a point mutation leading to a premature stop codon, which resulted in a predicted truncated AcrR. The MIC value in these isolates harboring the mutant acrR gene was higher than that in nonmutant isolates, but the difference was not significant (p = 0.066).
The tigecycline MICs of the two resistance strains (K803 and K860) with mutations both in ramR and acrR were 32 and 8 μg/mL, respectively. However, no mutation was detected in the rpsJ gene that codes for ribosomal protein S10.
Discussion
K. pneumoniae is an important pathogen that causes nosocomial infections, urinary tract infections, pneumonia, bacteremia, and liver abscess. 21 On exposure to several antibiotics, these strains rapidly acquire resistance through different molecular mechanisms. Recently, tigecycline has been reserved for use in situations when alternative treatments are not suitable. 3 As reported previously, susceptible strains of K. pneumoniae with decreasing susceptibility to tigecycline are commonly isolated during tigecycline therapy.12,22–24 Cross-resistance to tigecycline could be indirectly attributed to the use of other antibiotics, which are transported by the same chromosomally encoded efflux pumps in clinical isolates of K. pneumoniae. 25 However, the pathway and molecular-resistant mechanisms underlying cross-resistance are not yet clear.
In this study, we identified the susceptibility profile of tigecycline in 28 clinical K. pneumoniae isolates that had not previously been exposed to tigecycline in China. Based on CLSI guidelines, 20 isolates were not susceptible to tigecycline, and these were determined to be multidrug resistant (data not shown). RND-type efflux pump and transcriptional activators have been linked to the nonsusceptibility of tigecycline in a variety of Enterobacteriaceae. However, our real-time PCR results demonstrated that the differences in the expression of AcrAB, OqxAB, and RamA, RarA, SoxS, RarA between tigecycline-resistant and tigecycline-susceptible isolates were not significant, suggesting that the resistance might be due to efflux pumps other than AcrAB/RamA and OqxAB/RarA, such as KpgABC.
EPIs have been used to reduce resistance patterns due to efflux pump activation. 26 To evaluate the role of efflux pumps in tigecycline resistance of K. pneumoniae, we used a pump inhibition assay. We found that PAβN significantly restored tigecycline susceptibility in four strains. However, no significant reduction of tigecycline MICs was observed in the other 24 isolates, suggesting that PAβN was not effective. One possible explanation is that EPIs have different specificities, which interfere with efflux pump activity through different modes of action. 27 Our finding also indicated that mechanisms other than active efflux systems contribute to tigecycline resistance in clinical K. pneumoniae isolates.
The current study hypothesized that the use of other antibiotics might cause molecular-resistant mechanisms attributed to RND-type or other transporters that mediate a decrease in tigecycline activity. Studies by Hentschke et al. reported that mutations in the ramR gene led to the overexpression of RamA, causing a reduced susceptibility to tigecycline in K. pneumoniae and S. enterica.16,28 In K. pneumoniae, the ramR gene is located upstream of the ramA gene, in a head-to-head arrangement; because ramR is a negative regulator of ramA, mutations in ramR result in the overexpression of ramA. The entire genome sequence for five K. pneumoniae strains belonging to the sequence type 512 (ST512) was determined, and it was found that a mutation or insertion of mobile elements in the ramR gene occurred in the tigecycline-resistant strain. 18
Sheng et al. recently underscored the key role of RamA in tigecycline non-susceptible isolates by identifying several mutations in the acrR gene that led to amino acid substitutions in the AcrR protein. 24 Sequencing of ramR and acrR among the 28 clinical K. pneumoniae isolates revealed that frameshift deletions of fragment TGATGCGCTCGC spanning positions 254 to 265 in ramR gene (located in strains K977, K981), E175→D, I177→T, A178→D, and A193→S, were harbored in the ramR gene (located in strain K803), G121→V, E122→K of coding region of acrR gene or harbored premature stop codon associated with producing an amino acid substitution in AcrR protein.
Based on the real-time PCR result, K860, K898, and K1186 caused the overexpression of acrB by upregulating ramA. Moreover, strain K860 harbored missense and nonsense mutations in acrR, which led to the efflux pump AcrB-mediated resistance to tigecycline. To the best of our knowledge, this report is the first to identify these mutations in ramR and acrR in tigecycline resistance. Further experiments are ongoing to determine whether all of the mutation types reported here are detrimental to RamR or AcrR function.
Nonetheless, the above resistant regulatory pathways failed to explain clearly in three strains (K784, K805, K1107) of the tigecycline-nonsusceptible clinical samples, where the MICs of K805 and K1107 strains were significantly inhibited by PAβN and exhibited a low-level expression of efflux pump genes that was similar to other mechanisms contributing to the development of tigecycline resistance. Therefore, further studies need to elucidate the mechanisms underlying tigecycline resistance in these isolates, including determining the expression level of the efflux pump KpgABC. In addition, we need to identify other possible regulatory mechanisms of tigecycline resistance through transcriptome sequencing.
In conclusion, we identified a key role of RamA, through mutations of ramR and acrR, in strains that were not susceptible to tigecycline. Moreover, several types of mutations in ramR and acrR were identified, which were not reported previously, thus revealing possible molecular mechanisms that contribute to the development of tigecycline nonsusceptibility.
Footnotes
Acknowledgments
This work was supported by grant No. 81401697 from the National Natural Science Foundation of China, Grant no. 2015 M571314 from the China Postdoctoral Science Foundation, and Grant no.201501159 from the Doctoral Scientific Research Foundation of Liaoning Province.
Disclosure Statement
No competing financial interests exist.