
Abstract
Penicillin-resistant Streptococcus pneumoniae is a serious concern worldwide. In this study, we analyzed the cause of β-lactam resistance in pandemic multidrug-resistant clones. A total of 41 penicillin-nonsusceptible clinical isolates were collected from 1996 to 2012. Sero- and molecular typing confirmed that these isolates were clonal types of Taiwan19F-14, Taiwan23F-15, and Spain23F-1. Sero-switching was found in four isolates. All isolates were multidrug resistant. Sequencing analysis of the penicillin binding proteins (PBPs) was performed on PBP1a, 2b, and 2x, and a large number of mutations were identified in comparing to clinical penicillin-susceptible isolates and the recipient strain R6 used for homologous recombination. The T451A substitution was the key amino acid in PBP2b that contributed to penicillin resistance. T338A in PBP2x played a role in resistance and reached the highest level of resistance when combined with other mutations in PBP2x. High-level penicillin resistance could not be obtained without the combination of mutations in PBP1a with PBP2b and 2x. The amino acid substitutions in PBP1a, 2b, and 2x were the crucial factors for β-lactam resistance.
Introduction
S
Homologous recombination, which mediates the exchange of mosaic penicillin binding proteins (PBPs) among pandemic multiple drug-resistant pneumococci or highly homologous viridian streptococci, is one reason for the resistance.7,8 Mutations in PBPs [especially mutated residues encompassing the three active motifs SXXK, SXN, and KT(S)G] affect the reactivity of the active site serine recognized by penicillin. 9 Alternatively, mutations affect the structural integrity of PBP and deform the catalytic cleft required for efficient penicillin binding, resulting in low affinity or null affinity for penicillin.7,9
Among the six PBPs in S. pneumoniae, mutations across the triad of PBPs (PBP2x, 2b, and 1a) are critical steps in the development of penicillin-nonsusceptible Streptococcus pneumoniae (PNSSP).2,10,11 Among the 97 serotypes, 12 pneumococci in serotypes 19F and 23F are more frequently isolated and have a higher likelihood of having a PNSSP phenotype. 13
In this study, PNSSP clones of serotypes 19F and 23F, which have spread worldwide, were selected to investigate the causative mutations or sequences in the PBPs. Three relevant genes (pbp1a, 2b, and 2x) that were postulated to be involved in penicillin susceptibility were investigated.2,10,11
Materials and Methods
Bacterial isolates selected in this study
Clinical PNSSP were collected from 13 hospitals located from Northern, Central, and Southern Taiwan between 1996 and 2012. A laboratory penicillin-susceptible strain (R6) and two randomly selected penicillin-susceptible Streptococcus pneumoniae (PSSP) from each serotype (19F/23F) were incorporated in our study as controls. All pneumococci were stored in a −80°C freezer and grown on Columbia sheep blood agar plates at 35°C in an atmosphere with 5% CO2 for 20–24 hr. A pneumolysin-positive PCR was used to confirm that the bacteria were pneumococci.
Antimicrobial susceptibility testing
Empiric antibiotics against pneumococci, including penicillin, cefuroxime, cefotaxime, ceftriaxone, cefepime, vancomycin, chloramphenicol, erythromycin, meropenem, levofloxacin, moxifloxacin, tetracycline, and trimethoprim/sulfamethoxazole, were tested. The broth dilution method using SENSITITRE susceptibility plates for STP6F or STP3F (Thermo Scientific) operated by an automated inoculation delivery system under the revised guidelines of the Clinical and Laboratory Standards Institute (CLSI) 14 was adopted to determine their minimal inhibitory concentrations (MICs) toward S. pneumoniae. All runs were guarded by the control strain ATCC 49619.
Selection of serotype19F and 23F pneumococci
Capsular serotype typing of PNSSP isolates was confirmed through the Quellung reaction 15 using sera purchased from Statens Serum Institut (Copenhagen, Denmark).
Multilocus sequence typing
Multilocus sequence typing (MLST) was performed as described by Enright and Spratt. 16 The internal fragments of seven housekeeping genes (aroE, gdh, gki, recP, spi, xpt, and ddl) were amplified from our isolates using the MLST primers listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr). Sequencing of each amplicon and analysis were implemented according to the instructions in the pneumococcal MLST database. 17 Strains with the same genotype or identical allelic profiles or sequence type (ST) were considered to be the same clonal type. A single-locus variant was grouped as a clonal complex. The relatedness of associated allelic profiles was analyzed with the e-BURST V3 program. 18 The evolutionary descent was illustrated.
Genomic fingerprinting by pulsed field gel electrophoresis
PNSSP and three pandemic clones (Spain23F-1, Taiwan23F-15, and Taiwan19F-14) were incorporated in this study. Solidified pneumococcal plugs cast by mixing equal volumes of cell suspension buffer inoculated with pneumococci and 2% pulsed field gel electrophoresis (PFGE) agarose (Bio-Rad) plus 500 μg proteinase K (Sigma-Aldrich) were lysed with 3 ml of cell lysis buffer with the same amount of proteinase K at 37°C for 2 hr. Following two washes with 55°C ddH2O and TE buffer, the plugs were digested with 40 U of SmaI (New England Biolabs) in compliance with the instructions provided by the manufacturer. The DNA was separated in 1% PFGE agarose in 0.5× TBE buffer at 14°C for 22.5 hr at 200 V with a ramping time of 1.79–35.8 sec using the Bio-Rad CHEF-DRII apparatus (Bio-Rad Laboratories). The resulting genomic DNA fingerprints were analyzed by the Molecular Analyst Finger-printing DST version 1.6 software program (Bio-Rad).
Sequence analysis of pbp1a, 2b, and 2x
Three pbps (pbp1a, 2b, and 2x) were amplified in a 25 μl reaction mixture using 0.6 μM of the forward and reverse primer pairs of pbp1a-F-up227/pbp1a-R-dn219, pbp2b-F-up330/pbp2b-R-dn134, and pbp2x-F-up110/pbp2x-R-dn379 (Supplementary Table S1) with Phusion Hotstart Taq polymerase (Thermo Scientific), 1× Phusion HF buffer, and 250 μM deoxynucleotide triphosphates (GeneTeks BioScience). The program included a hot start of 98°C for 30 sec, followed by 35 cycles of 98°C for 10 sec, annealing for 10 sec, 72°C for 1.5 min (30 sec/kb), and finally 72°C for 10 min for extension. Amplicons were sequenced using the corresponding sequencing primers listed in Supplementary Table S1. An alignment between the three PBPs from R6, PSSP, and PNSSP was constructed for preliminary analysis.
Construction of penicillin-nonsusceptible mutants by homologous recombination of pbps
As penicillin resistance was introduced through the stepwise exchange of individual pbps or in mutuality in R6, the region encoding transpeptidase (TP) domain across the three pbps (2b, 2x, and 1a) was amplified individually from the three clinical PNSSP representatives using proprietary primer pairs (serotype specific): pbp1a-F1-PstI and pbp1a-R2160-BamHI, pbp2b-F118-BamHI and pbp2b-R-dn15-EcoRI, and pbp2x-F52-BamHI/pbp2x-F145-BamHI and pbp2x-R2248-EcoRV (listed in Supplementary Table S1). The PCR products were simultaneously digested with the chloramphenicol-resistant suicide vector pPCR-Script (Stratagene) using the restriction endonucleases (Fermentas; Thermo Scientific, Inc.) designated with their primers in Supplementary Table S1. The digested product was cloned into the dephosphorylated vector using FastAP Thermosensitive Alkaline Phosphatase (Fermentas; Thermo Scientific, Inc.) through the T4 DNA ligase (Fermentas; Thermo Scientific, Inc.) in accordance with the instructions of the manufacturer. Successful ligates (pPCR-Script-T238-pbp1a/-2b/-2x, pPCR-Script-T22-pbp1a/-2b/-2x, and pPCR-Script-T33-pbp1a/-2b/-2x) verified to possess the correct size and sequence were heat-shocked into Top10 competent cells. After transformation, derivative clones that were white in color, resistant to chloramphenicol, and had the correct sized insert-embedded plasmid were used for further homologous recombination with the PSSP R6.
Before the homologous recombination of pbps, R6 was primed with the competence stimulating peptide (240 ng/μl) according to the method of Chen et al., 19 using pPCR-Script-TP-pbp1a, -2b, or -2x exchange plasmids. MICs of penicillin and cefotaxime of the homologous recombinants (HRs) selected from CAT agar plates 20 as the most penicillin resistant (PR) were measured using the same antimicrobial susceptibility testing system.
Site-directed mutagenesis of an isogenic variant
Mutations correlated with penicillin susceptibility were evaluated by site-directed mutagenesis (SDM). Reversion of PBP2b A451T and PBP2x A338T in the constructs pPCR-Script-T238-pbp2b and -2x was generated using the primer sets pbp2b-F1331-SDM-A451T and pbp2b-R1373-SDM-A451T and pbp2x-F992-SDM-A338T and pbp2x-R1034-SDM-A338T, respectively (Supplementary Table S1), with the QuikChange® II Site-Directed Mutagenesis kit (Stratagene). The chimeric plasmids underwent homologous recombination with triple-pbp exchange. Using the replica methodology, the daughter clones that exhibited the largest drop in their penicillin MICs compared to the parental HR were selected for further testing. The degree of nonsusceptibility to penicillin induced by a specific amino acid (a.a.) substitution was noted.
Statistical analysis
Significant differences in the susceptibility rate among various antibiotics between serotypes 19F and 23F were calculated using Fisher's exact test. A p-value smaller than 0.05 represented a significant difference, while a p-value smaller than 0.001 indicated a strongly significant difference.
Results
Bacterial isolates and serotype confirmation of 19F and 23F
A total of 41 PNSSP, collected between 1996 and 2012, were isolated from 7 different sample types, including 23 blood isolates, 7 pus isolates, 4 pleural fluid isolates, 3 CSF isolates, 2 urine isolates, and 1 each from ascites and sputum. Among the 41 cases, 4 patients were confirmed with meningitis. The serotype confirmation test showed that 20 isolates belonged to serotype 19F, and 21 isolates were identified as serotype 23F.
Molecular epidemiological typing of PNSSP
In the MLST, ST81, 236, and 242 were characterized and referred to three different pandemic circulating types: the clonal types Spain23F-1, Taiwan19F-14, and Taiwan23F-15. From our clinical PNSSP, three predominant clonal types, belonging to 19F and 23F, with eight STs were obtained (Table 1).
Bold, unmatched results between genotype and serotype.
Clones represented international antibiotic-resistant clones.
New variant or ST type found in this study.
MLST, multilocus sequence typing; PNSSP, penicillin-nonsusceptible Streptococcus pneumoniae; ST, sequence type in MLST.
Four STs (ST236, 271, 320, and 1464) that were Taiwan19F-like isolates were identified. According to the e-BURST V3 analysis, the isolates that formed a clonal complex of Taiwan 19F and ST271 were the predicted founders, suggesting that ST236, 320, and 1464 were derived from ST271. The isolates differed in the aroE or ddl allele. One ST1464 isolate had a genotype that was reported to be specific to serotype 19F but was confirmed to be serotype 23F in this isolate. This capsular serotype switching phenomenon was also found in the clonal types Taiwan23F-15 and Spain23F-1, in which one and two isolates, respectively, with clonal types of Taiwan23F-15 and Spain23F-1 were found to have the 19F serotype (Table 1).
Clonal type 23F isolates, including Taiwan23F-15 clonal type and Spain23F-1 clonal type, comprised four STs; ST242 belonged to the Taiwan23F-15 clonal type, while the other three STs (ST81, 83, and 2085) belonged to the Spain23F-1 clonal type. ST2085 was a novel ST identified in this study (Table 1).
PFGE phylogenetic correlation analysis of our isolates confirmed that the MLST types and clonal types were Taiwan23F-15, Taiwan19F-14, and Spain23F-1 (Fig. 1). Eleven small clusters with PFGE similarities >80% were classified as clonally related strains and designated groups I to XI. When the PFGE results were combined with the molecular typing results, the first six clonally related PFGE clusters were categorized as Spain23F-1. The clonally related PFGE VII cluster was categorized as Taiwan23F-15. The remainder of the clonally related PFGE VIII to XI clusters belonged to the Taiwan19F-14 clonal type.
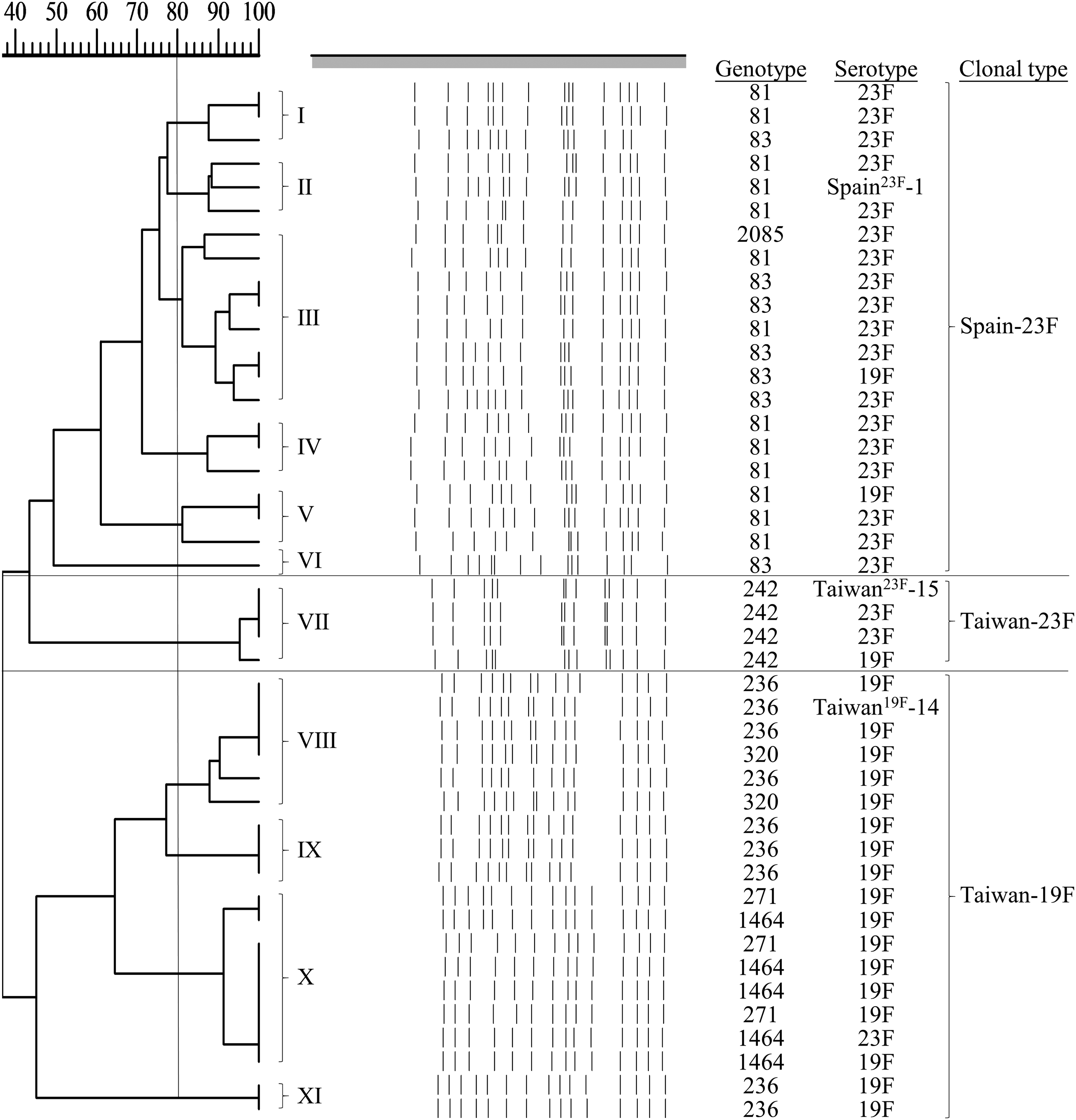
Molecular typing among 41 PNSSP. PNSSP, penicillin-nonsusceptible Streptococcus pneumoniae.
Antimicrobial susceptibility testing
The MIC50, MIC90, ranges and the percentage of susceptibility of the pneumococci to various antibiotics are listed in Table 2. The MIC for penicillin among the selected isolates ranged from 4 to ≥8 μg/ml. According to the CLSI guidelines, none of the isolates was susceptible to penicillin. Moreover, 10 isolates were defined as PR, including 4 cases that originated from meningitis, with penicillin MICs of 4 μg/ml. The remainder of the isolates were from other isolation sites and had MICs for penicillin that were ≥8 μg/ml. Of these 10 penicillin-resistant Streptococcus pneumoniae isolates (PRSP), 7 belonged to serotype 19F (70%) and 3 belonged to serotype 23F (30%). A total of 40% of the pneumococci were cefotaxime resistant (n = 4). All of the isolates were susceptible to vancomycin, levofloxacin, and moxifloxacin. None of the isolates was susceptible to cefuroxime, erythromycin, meropenem, and tetracycline. No isolate with serotype 23F was susceptible to cefotaxime, ceftriaxone, and cefepime, while 20%, 5%, and 40% of the isolates with serotype 19F were susceptible to cefotaxime, ceftriaxone, and cefepime, respectively. Significant differences in susceptibility toward chloramphenicol and cefepime were observed between the 19F and 23F isolates (Table 2). Isolates with serotype 19F were more susceptible to cefepime than isolates with serotype 23F (p = 0.001, Fisher's exact test). In contrast, isolates with serotype 19F were significantly more susceptible to chloramphenicol than isolates with serotype 23F (p < 0.001, Fisher's exact test).
Significant differences in the susceptibility rate between serotype-23F and serotype-19F by Fisher's exact test (p < 0.05).
MIC, minimal inhibitory concentration (μg/ml); SXT, trimethoprim/sulfamethoxazole (19:1).
Sequence analysis of PBP1a, PBP2b, and PBP2x
Three penicillin-susceptible strains, including two randomly selected clinical isolates with serotype 19F or 23F and the recipient R6 used for homologous recombination, were used as control sequences for comparison of the a.a. sequences of PBP1a, 2b, and 2x between the 41 PNSSP (Supplementary Figs. S1–S3). Hot spot mutations in PNSSP that may contribute to penicillin susceptibility were identified. The PBP1a and 2b sequences of two randomly selected clinical PSSPs were concordant to R6 with the exception of a few sporadic mutations. One PSSP isolate did not resemble R6 PBP2x (Supplementary Fig. S3).
For PBP1a, two mutations at positions 388 and 533 were not related to penicillin resistance because they occurred in clinical PSSP isolates. A total of 36–41 mutations (11.1–12.7%) were located in this TP (Table 3). Twenty-nine consensus mutations were found in all three serotypes (Supplementary Fig. S1). Two patterns of mutations were observed among the 41 PNSSP. Forty-one (12.7%) and 36 (11.1%) consensus mutations were found in Taiwan 19F/23F and Spain 23F, respectively (Table 3). Around the three active motifs, the STMK370–374, mutation at position 371 with SA/SMK (T371A or S) was observed in all of our PNSSP. The T371S substitution (SSMK) was specifically found in Taiwan-19F/23F, while the T371A substitution (SAMK) was only found in Spain 23F. The mutation located near the second active motif at a.a. position 432 (P432T) was concomitantly found in all of the PNSSP with the SRN428–430 active site mutation. No change was observed in the third motif KTG557–559.
Bold, active motif sites.
Clonal group, including all clonally related types in MLST.
Number of amino acids mutated in TP of PNSSP minus the mutations that occurred in PSSP.
PSSP, penicillin-susceptible Streptococcus pneumoniae; TP, transpeptidase.
For PBP2b, the clinical PSSP possessed amino acid sequences identical to R6 with the exception of one mutation at G602E (Supplementary Fig. S2). Among the 41 PNSSP, 7 consensus mutations (2.1%) were found. Amino acid changes ranged from 11 to 39 (3.4–11.9%) in TP (Table 3). The only a.a. change that was near the active motifs was SSNT448–451 to SSNA (T451A). The other two active motifs remained unchanged in all 41 PNSSP (Table 3). Among the Taiwan19F clonal type, the lowest number of mutations occurred in ST236. This ST was collected during the earliest period, suggesting that substitutions arising from mosaicity accumulated over time. Taiwan23F and Spain23F possessed much less variation in terms of their number of mutations. Genetic variants in Taiwan23F had 15 or 16 mutations, while variants in Spain23F had 11 or 12 mutations (Supplementary Fig. S2). Q443E and T451A were unique mutations close to the SSNT motif and were prevalent in our PNSSP strains (Supplementary Fig. S2 and Table 3). Some distinct point mutations were observed, such as A521S in the Spain23F variants, S362A, D369N, and K371E in the Taiwan23F variants, and I366L in the Taiwan19F variants. The E338G and E374D substitutions were distributed nonspecifically among Spain23F and Taiwan19F and 23F (Supplementary Fig. S2).
For PBP2x, 8 consensus mutations (2.5%) with a range of 10–15 mutations (3.1–4.7%) were found in the TP domain of PBP2x when the mutations in PSSP were eliminated (Supplementary Fig. S3 and Table 3). PBP2x was very similar between Taiwan23F and Spain23F (Table 3). The mutation T338A, which was demonstrated to be associated with penicillin resistance, was found in all PNSSP (Table 3). I371T, which was important for penicillin and cefotaxime nonsusceptibility, was present in all of our PNSSP. Y595F, 21 which was associated with a fourfold increase in the MICs of penicillin and cefotaxime, was identified in two of our strains; however, these isolates did not show a higher MIC for penicillin or cefotaxime than isolates without this mutation. M339F22,23 and M400T,7,22 which had been observed to be specific for cefotaxime resistance, were also identified in our isolates and associated with resistance to cefotaxime (data not shown). N514H, which had been reported to be associated with penicillin resistance, 24 was also found in both penicillin intermediate or resistant isolates, indicating that the concomitance of this mutation with other penicillin-related mutations did not play a significant role in penicillin nonsusceptibility. L546V 22 and D567N 22 were excluded from the analysis of the relationship to penicillin resistance because these mutations were also found in the PSSP strains. N605T 22 was present in all of our strains despite the fact that it was previously reported to be unrelated to penicillin resistance. 21
Homologous recombination of pbp1a, 2b, and 2x from PNSSP into the recipient R6
A two- to fourfold increase in penicillin resistance was obtained following a single TP pbp switch into R6 (Table 4). The HRs of pbp1a from Taiwan19F, Taiwan23F, and Spain23F could not be selected in the experiment. Random selection of colonies from the agar plate with or without antibiotic selection did not show evidence of pbp1a recombination, indicating no significant change in the penicillin MIC for the selection of the recombinant or a low frequency of recombination (Table 4). Recombinants of pbp2b were selected from serotype 19F and showed no or an insignificant change in the penicillin MIC (0.06 μg/ml). A significant increase in the MIC (fourfold increase from 0.12 μg/ml) for cefotaxime was found in the pbp2x recombinants. The pbp2x alteration was the only pbp involved in cefotaxime susceptibility.
Bold, significant change in the MIC in PEN or FOT after homologous recombination of different PBPs relative to R6.
FOT, cefotaxime; NS, unable to select recombinant; PBP, penicillin binding protein; PEN, penicillin.
The recombinant with the donor sequence from Taiwan19F was selected for the double recombination of pbp1a/2b. This recombinant only exhibited a twofold change in the penicillin MIC (to 0.06 μg/ml). No change in the cefotaxime MIC was detected in the recipient (Table 4). The pbp2x/1a or pbp2b/2x recombinants exhibited a significantly increased MIC (4- to 16-fold increase) for both penicillin and cefotaxime. pbp2b/2x and pbp1a/2x donated from Taiwan23F showed more resistance to penicillin and cefotaxime than genes from Taiwan19F and Spain23F (Table 4).
The highest MICs of penicillin and cefotaxime (2–4 μg/ml, respectively) were obtained for the triple recombinant pbp1a/2b/2x compared to all other types of recombinants, indicating that all three pbps were involved in penicillin and cefotaxime resistance.
Restoration of mutated amino acids close to or within the active motif in pbp1a, 2b, and 2x
Restoration of the a.a. sequence in pbp2b at A451T or pbp2x at A338T in the triple recombinant with the sequence from Taiwan19F resulted in a fourfold drop in the original MIC of penicillin from 2 to 0.5 μg/ml (Table 5). The restoration of pbp2b at A451T resulted in a penicillin MIC that was identical to the double mutant recombinant pbp1a/2x, indicating that A451T was the key mutation contributing to penicillin resistance in pbp2b (Table 5). Restoration of A338T in pbp2x was unable to cause the MIC to drop back to a level identical to the double recombinant pbp1a/2b, indicating that other mutations in pbp2x played a role in penicillin resistance. Restoration of pbp1a at A or S371T and T432P could not be selected, indicating that the drop in the penicillin MIC was insignificant (Table 5).
NS, unable to select complementary strain.
Discussion
Community-acquired S. pneumoniae infection with penicillin resistance is a serious concern worldwide, especially in children younger than 5 years old. Although a 13-valent conjugate vaccine was recently launched in the market, not all children in nondeveloped countries were vaccinated and serotypes of escaped strains from the nonvaccine coverage type have been documented with the increasing incidences. 25 In Asia, multidrug-resistant strains could account for up to 60%. Serotypes 19F and 23F were on the top of the list with high prevalence rates (23.5% in 19F and 10% in 23F). 26 In this study, we analyzed the cause of penicillin nonsusceptibility in these two serotypes.
Among the 41 isolates with PNSSP that were collected in this study, three pandemic clonal types were identified (Taiwan19F-14, Taiwan23F-15, and Spain23F-1). According to the MLST analysis, the variant types found in these serotypes indicated evolutionary change. Sero-switching was found in all three clonal types (Taiwan19F-14, Taiwan23F-15, and Spain23F-1). Although the phenomenon occurred in all three clones, PFGE analysis showed that these three pandemic strains were highly distinct (Fig. 1). The phenomenon of sero-switching may contribute to the emergence of nonvaccine coverage types with a multidrug-resistant background in the future.27–30 The application of molecular typing and serotyping together in the same epidemiology study should be used to explore the above possibility. 31
In this study, we have selected strains with high penicillin MICs (4 or 8 μg/ml) to ensure mutation/s of PBPs that could be identified in sequence analysis. Although there are six PBPs in S. pneumoniae, three have been demonstrated to be involving in penicillin resistance. The sequence analysis revealed that mutations were sprinkled among PBP1a, 2b, and 2x compared to clinical susceptible isolates and the recipient R6. Several studies have documented different mutations that may be related to penicillin susceptibility. We confirmed the presence of most of these previously described mutations in our strains (Table 3). These previous studies primarily used enzyme kinetics to determine the contribution of the mutations to penicillin susceptibility, with the exception of Smith et al.21,32
Among these previously documented mutations involved in penicillin susceptibility, mutations in or close to the three active motifs were mostly evaluated using epidemiology or kinetic studies; these mutations were also identified in our isolates. Single gene recombinants except pbp1a and different combinations of pbps were obtained in the homologous recombination of pbp1a, 2b, and 2x from the Taiwan19F, 23F and Spain23F isolates. The recombinants showed susceptibility differences compared to the recipient R6. The MIC of penicillin for these recombinants confirmed that all of the pbps were involved and contributed different degrees of resistance to penicillin and cefotaxime (Table 4). Single pbp2b and 2x HRs had only twofold differences in penicillin susceptibility, indicating that the single gene mutation in any one of these pbps was unable to produce a significant change in the MIC of penicillin. This observation has also been reported by other groups. 33 Moreover, pbp2x recombinants had a significant effect on cefotaxime, achieving a fourfold or more difference in the MIC. Unlike pbp1a and 2b, pbp2x had a relatively more specific contribution to penicillin and cefotaxime resistance. The double HR pbp1a/2b could only be selected from the donor sequence of Taiwan19F-14. There was only a twofold difference in the MIC for penicillin and no MIC difference for cefotaxime. These MIC results were identical to those obtained for the single pbp2b recombinant, suggesting that the additional pbp1a recombination did not have an additive or synergic effect on penicillin susceptibility. 34 In contrast, the addition of pbp2x into the pbp1a or pbp2b recombinants caused a significant change in both penicillin and cefotaxime susceptibility compared to the pbp1a or pbp2b recombinants. This observation suggests that pbp2x is the most important contributor among all of the pbps to penicillin and cefotaxime resistance. In our triple pbp1a/2b/2x recombinant, penicillin MICs were remarkably increased (≥64-fold), indicating the synergic effect on these three pbps on resistance.
The restoration of amino acid sequences in or close to the three active motifs in the triple recombinant revealed that we were able to select complementary strains with the exception of pbp1a restoration. However, the conversion at TSQF574–577NTGY in pbp1a that conferred a contribution to penicillin resistance using site-specific mutagenesis had been previously reported. 32 These mutations were also found in clinical isolates. Because we are unable to select the restoration strain with S371T and T432P in the active motif, we could not exclude the possibility that the mutation at a.a. 371 and/or 432 had an effect on penicillin susceptibility. TSQF574–577NTGY in pbp1a may be one of the most important point mutations in our resistant isolates. T451A in pbp2b has been demonstrated to be involved in penicillin susceptibility in kinetic studies. The restoration of A451T in pbp2b in the triple recombinant, resulting in a strain that was identical to the pbp1a/2x recombinant, had the same penicillin MIC as the pbp1a/2x recombinant. Although there were different mutations in pbp2b in the clinical penicillin-resistant isolates, the T451A mutation located close to the active site motif was the fundamental amino acid that contributed to penicillin resistance in our study. Unlike pbp2b, the restoration of A338T in pbp2x showed a drop in the penicillin MIC from 4 to 0.5 μg/ml but did not reach the same level (0.06 μg/ml) as the pbp1a/2b recombinant with the identical sequence. This result suggested that the T338A mutation was not the only amino acid that contributed to penicillin susceptibility; therefore, other mutations in pbp2x should be involved. In addition to T338A, S389L, M339F, I371T, R384G, M440T, N514H, D567N, and N605T that were associated with penicillin susceptibility in previous kinetic studies were also identified in our clinical isolates. Whether any of these mutations in combination with T338A in PBP2x will contribute to resistance needs further study.
Taken together, our results demonstrate that all three of these proteins are important and contribute to penicillin resistance to differing degrees. T451A is the key mutation in PBP2b contributing to penicillin resistance. T338A combined with other mutations in PBP2x should play a role in our clinical penicillin-resistant isolates. PBP1a acts together with PBP2b and 2x to reach a high-level penicillin resistance. These results highlighted the important information for the development of penicillin class of antibiotic.
Footnotes
Acknowledgments
We express our sincere thanks to Taiwan Centres for Disease Control and National Health Research Institutes for providing us serotype-19F and serotype-23F penicillin-resistant pneumococci strains in this study. This work was partly supported by the grants of Ministry of Science and Technology of Taiwan (103-2314-B-010-052-MY2), and partly from the grants MOHW103-CDC-C-114-135404-07, MOHW104-CDC-C-114-144406-07 from Taiwan Centres for Disease Control and National Health Research Institutes.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.