
Abstract
The emergence and dissemination of NDM-1 (New Delhi metallo-β-lactamase-1)-producing Enterobacteriaceae have resulted in a worldwide public health risk. This study described a high incidence and endemic spread of NDM-1-producing extensively drug-resistant Escherichia coli in a teaching hospital in Zhejiang province, China. We recovered six nonduplicated NDM-1-producing E. coli isolates from May 2014 to August 2014 with positive modified Hodge test and EDTA synergistic test. These isolates were highly resistant to β-lactam antimicrobials, aminoglycosides, and quinolones. PCR and DNA sequences analysis showed that all isolates carried the blaNDM-1, blaSHV-11, aac(6′)-ib-cr, and qnrB. Several isolates also harbored blaCTX-M-1, blaCTX-M-9, rmtB, and qnrA. Southern blot confirmed that blaNDM-1 was located on the same ∼55 kb plasmid and conjugation experiments further proved the contransferable characteristic of blaNDM-1. The ompC sequences showed various mutations, which was related to multidrug resistance in E. coli. Pulsed-field gel electrophoresis identified four of six isolates that belonged to the same genotype. Multilocus sequence typing assigned them to ST2, except one strain that belonged to ST594. Our study demonstrated that the resistance-associated genes and the loss of the outer membrane proteins could account for high resistance of NDM-1-producing E. coli to multiple antimicrobial drugs. Both horizontal transfer of IncN and transmission of ST2 were responsible for the spread of drug resistance. These findings highlighted an urgent need to limit the further dissemination of NDM-1-producing E. coli in this region.
Introduction
E
NDM-1 (New Delhi metallo-β-lactamase-1), a carbapenemase that can hydrolyze all β-lactams except aztreonam, was first identified in India and now worldwide. 4 In China, the presence of blaNDM-1-positive E. coli was first reported in HongKong, which was isolated from a 1-year-old infant and his mother. 5 More recently, the epidemic dissemination of NDM-1-producing E. coli has been confirmed in many cities in Henan, Guangdong, Hunan and Zhejiang provinces,6,7 causing a public health threat.
Plasmid location was one of the primary cause for the rapid dissemination of the blaNDM-1 gene. In the United Kingdom and India, NDM-1 was found on plasmids with sizes from 50 to 300 kb, which belonged to different plasmid incompatibility (Inc) groups such as A/C and FI/FII. 8 In our study, IncN plasmids were the most common plasmid types, which was different from other countries.
Moreover, loss or alteration of outer membrane proteins (OMPs), including OmpF and OmpC, were related to multidrug resistance in E. coli, particularly when combined with production of extended-spectrum β-lactamases (ESBLs) or AmpC enzymes.9,10
In this study, we explored the molecular characteristics and epidemiology of six NDM-1-producing extensively drug-resistant (XDR) E. coli isolates recovered from a teaching hospital in China during May 2014 to August 2014.
Materials and Methods
Bacterial isolates
Six nonrepetitive XDR E. coli clinical isolates were recovered from the First Affiliated Hospital of Wenzhou Medical University in China during May 2014 to August 2014, which were detected to harbor blaNDM-1 by PCR and sequencing. All isolates were identified by a VITEK-2 compact system (bioM áerieux).
Antimicrobial susceptibility testing
Antimicrobial susceptibility of each isolate to cefepime (FEP), ceftriaxone (CRO), ceftazidime (CAZ), cefotaxime (CXM), tobramycin (TOB), gentamicin (GEN), imipenem (IPM), meropenem (MEM), ertapenem (ETP), levofloxacin (LEV), ciprofloxacin (CIP), amikacin (AK), sulphamethoxazole/trimethoprim (SXT), sulbactam/ampicillin (SAM), and piperacillin/tazobactam (TZP) was assayed by the agar dilution method and interpreted according to the Clinical and Laboratory Standard Institute (CLSI) 2014 recommendations. 11
Phenotypic screening and confirmation of NDM-1 producers
The metallo-β-lactamase (MBL) phenotype test was determined by the modified Hodge test (MHT) and EDTA-disk synergy test according to Lee et al. 12
PCR amplification and DNA sequence analysis of resistance genes
Total DNAs of all isolates were extracted using the AxyPrep Bacterial Genomic DNA Miniprep kit (Axygen Scientific) and were used as templates for PCR amplification. The β-lactamase genes (blaTEM, blaSHV, blaCTX-M-1, blaCTX-M-9, blaKPC, blaGES, blaSPM, blaIMP, blaVIM, blaOXA-23, and blaOXA-48), 13 plasmid-mediated quinolone resistance (PMQR) genes (qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr), 14 chromosome-mediated quinolone resistance genes (gyrA and parC) 15 and 16S rRNA methylase genes (armA, rmtA, rmtB, rmtC, and rmtD) 16 were amplified by PCR methods as previously described. The positive PCR products were sequenced in a DNA analyzer (ABI PRISM 3700) by Shanghai Majorbio Bio-Pharm Technology Co. Nucleotide sequences were analyzed and compared using BLAST (www.ncbi.nl m.nih.gov/BLAST).
Conjugation experiments
The transferability of blaNDM-1 mediated by plasmids was confirmed by broth mate conjugation assays, using azide-resistant E. coli J53 as the recipient strain. Transconjugants were selected on Mueller–Hinton agar plates containing sodium azide (100 μg/ml) and meropenem (0.5 μg/ml). MICs of several representative agents for transconjugants were measured and transconjugants were screened for MBL by MHT and EDTA-disk synergy test. Then, the presence of resistance genes were verified via PCR analysis.
Plasmid analysis and Southern blot
The total bacterial DNAs of all donor isolates and transconjugants were first digested with S1 nuclease (Takara Bio, Inc.) and analyzed in a CHEF-Mapper XA PFGE system (Bio-Rad) with the following conditions: temperature 14°C, voltage 6 V/cm, pulse angle 120°, and pulse duration of 5–35 sec for 18 hr. Then, the DNA in gel were transferred to a nylon membrane (Roche Diagnostics) by the capillary method, fixed by a Stratalinker UV cross-linker (Stratagene). Eventually, the membranes hybridized with digoxigenin-labeled blaNDM-1 probes and detected with the Detection Starter Kit II (Roche). Salmonella H9812 served as a size marker.
Incompatibility typing of the blaNDM-1 plasmid
To classify the plasmids, 18 pairs of primers (FIA, FIB, FIC, HI1, HI2, I1-Ig, L/M, N, P, W, T, A/C,K, B/O, X, Y, F, and FIIA) were designed to perform five multiplex- and three simplex-PCRs by a PCR-based replicon typing method according to Carattoli et al. 17
Analysis of OMPs
The outer membrane proteins OmpA, OmpF, and OmpC were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) by membrane extracts, using E. coli K12 as a control for porin profiles. ompA, ompF, and ompC structural genes were amplified by PCR methods 3 and the predicted amino acid sequences were compared with reference protein sequences of E. coli K12.
Pulsed-field gel electrophoresis
Clonal relatedness among the NDM-1-producing isolates was performed using the XbaI restriction enzyme (Takara Bio, Inc.) and a CHEF-Mapper XA PFGE system, with the following conditions: temperature 14°C; voltage 6 V/cm; pulse angle 120°, and pulse duration of 5–35 sec for 20 hr. Patterns were analyzed using Quantity One software to generate a dendrogram and interpreted according to the criteria proposed by Tenover et al. 18
Multilocus sequence typing
Multilocus sequence typing (MLST) of E. coli isolates harboring blaNDM-1 were implemented using eight conserved housekeeping genes (dinB, icdA, pabB, polB, putP, trpA, trpB, and uidA) according to protocols available at a MLST website (http://bigsdb.web.pasteur.fr/ecoli/primers_used.html).
Results
Clinical characteristics of the patients
A total of six individual E. coli were isolated from clinical specimens, including blood, urine, sputum, and pus during May 2014 to August 2014 (Table 1). Patients were mainly hospitalized in the comprehensive ward and all of them had been submitted to invasive procedures. In addtion, they were primarily elderly (range 45–66 years old) and had long-term hospitalization (range 12–75 days), which probably associated with wide prevalence of these isolates. Nevertheless, we noticed that six patients harboring NDM-1-producing E. coli had never traveled abroad.
Antimicrobial susceptibility testing, phenotypic detection, and detection of resistance genes
The results of antimicrobial susceptibility testing for six E. coli isolates were shown in Table 2. Six isolates were highly resistant to all categories of β-lactam antimicrobials, aminoglycosides, quinolones, and other clinical antibacterial agents, which were defined as XDR. 19 The MHT and EDTA-disk synergy tests were both positive for six isolates. PCR and sequencing indicated that all isolates harbored blaSHV-11, aac(6′)-ib-cr, and qnrB. Furthermore, several isolates also harbored qnrA, rmtB, blaCTX-M-1, and blaCTX-M-9 (Table 2). In addition, sequencing of the amplified DNA revealed that all isolates possessed two quinolone resistance-determining region (QRDR) mutations in gyrA and at least one QRDR mutation in parC (Table 2).
J2303, J2504, J2543, J2516, J2547, J917 representing 6 transconjugants of XDR isolates, respectively.
NDM, New Delhi metallo-β-lactamase-1; PMQR, plasmid-mediated quinolone resistance; XDR, extensively drug-resistant; FEP, cefepime; CRO, ceftriaxone; CAZ, ceftazidime; CXM, cefotaxime; TOB, tobramycin; GEN, gentamicin; AK, amikacin; MEM, meropenem; ETP, ertapenem; IPM, imipenem; LEV, levofloxacin; CIP, ciprofloxacin; SXT, sulphamethoxazole/trimethoprim; TZP, piperacillin/tazobactam.
Conjugation experiments
Transfer of blaNDM-1 gene from six isolates to E. coli J53 by conjugation was successful. Transconjugants showed resistance or decreased susceptibility to all β-lactams, β-lactam/inhibitor combinations, aminoglycosides, quinolones, and carbapenems (Table 2). PCR and sequencing for transconjugants further confirmed the successful transfer of blaNDM-1 and blaCTX-M-9 in these isolates.
Plasmid analysis and Southern blot
The results of S1-pulsed-field gel electrophoresis (PFGE) showed that six isolates carried a least two plasmids with sizes from 33.3 to 244.4 kb (Fig. 1A). Southern blot confirmed that the blaNDM-1 gene was located on plasmids of ∼55 kb in five clinical isolates and their transconjugants, whereas in the remainder it was detected on a ∼45 kb plasmid (Fig. 1B).
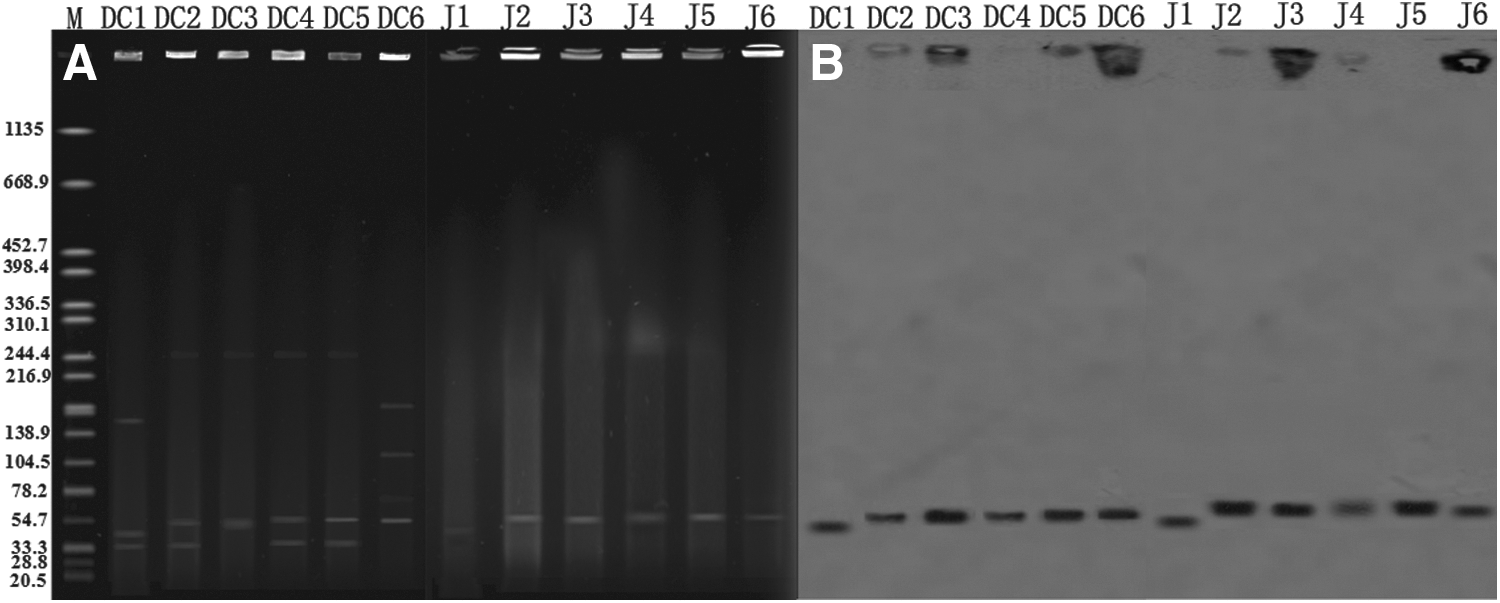
Incompatibility typing of the blaNDM-1 plasmid
Among six NDM-1-producing isolates, amplification successfully recognized four isolates and their transconjugants carrying IncN plasmids. In addition, DC2303 and its transconjugant carried IncF I, DC917, and its transconjugant carrying IncY-F I.
Examination of OMPs and porin genes
The OMP profiles of six XDR isolates showed the expression of OmpA and either OmpF or OmpC, whereas the wild-type strain E. coli K12 expressed OmpA, OmpF, and OmpC. The results of SDS-PAGE are shown in Fig. 2. PCR and sequence analysis of OmpF revealed that there were no changes between E. coli K12 and the clinical isolates, whereas the ompC sequences showed various mutations and deletions that were indicated to lead to changes of amino acid (details seen in Fig. 3).
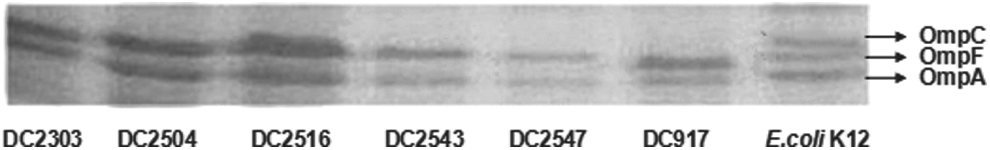
Omp profiles of E. coli K12 and six XDR E. coli. Omp, outer membrane proteins.

Comparison of predicted amino acid sequences of OmpC with reference protein sequences. Alignment of regions of deduced OmpC sequences, which exhibited variation relative to that of CQR81715.1. The region of consensus sequences were marked by the black boxes, substitutions are shaded gray, and deleted amino acid residues are shown as dashes.
Bacterial clonal relatedness
PFGE analysis revealed that four of six NDM-1-producing isolates belonged to the same genotype, whereas the DC2543 and DC917 showed unique restriction banding patterns (Fig. 4). MLST confirmed that DC917 and these four isolates with closely related banding patterns belonged to ST2 (allelic profile 8-2-7-3-7-1-4-2), except DC2303, which belonged to ST594 (allelic profile 31-195-11-36-27-67-26-125).

The pulsed-field typing of six NDM-1-producing XDR E. coli.
Discussion
E. coli, one of the most common enteric microorganisms, is an opportunistic pathogen associated with both community-acquired and nosocomial infections, including pneumonia, urinary tract infections, and wound infections. 20 The increasing prevalence of XDR E. coli isolates has been associated with higher morbidity and mortality rates, posing a considerable threat to public health. In this study, we provide the detailed data on the molecular characterization and epidemiologic study of XDR E. coli isolates during May 2014 to August 2014 in a teaching hospital. It's worth noting that all six XDR isolates carried blaNDM-1. As previously reported, NDM-1-producing isolates were resistant to all the β-lactamase antibiotics including some highly potent antibiotics like carbapenems. 21 In our present study, the NDM-1-positive isolates were also highly resistant to all antibiotics frequently used in the clinic, which may present a serious problem because tigecycline and colistin would be the only useful therapies against the NDM-1-producing isolates.
To better characterize the resistance mechanism of NDM-1-producing E. coli, we analyzed the antibiotic resistance genes. Our study revealed a wide distribution of diverse resistance determinants among the NDM-1-producing isolates, possibly interpreting why NDM-1-producing isolates are highly resistant to various kinds of antimicrobial agents. In this study, five isolates carried rmtB, one of the 16S rRNA methylase, which can confer high level resistance to the majority of clinically important aminoglycosides. 22 In particular, DC917, detected negative for the known 16S rRNA methylase genes, was resistant to amikacin, tobramycin, and gentamicin, indicating the presence of other mechanisms of aminoglycoside resistance in this isolate. In studies, six isolates also harbored blaCTX-M, in accordance with previous studys that most of 16S rRNA methylase gene-positive isolates were found to be ESBL producers, especially CTX-M type. 23 As previously reported, CTX-M production was significantly associated with resistance to quinolones and aminoglycosides, leading to a high prevalence of multiresistant isolates. 24 Moreover, all isolates carried double gyrA mutations at codon 83 and 87, also a least one parC mutation at codon 80, which were considered to be an important step in the acquisition of fluoroquinolone resistance. Two PMQR genes, including aac(6′)-Ib-cr and qnrB, were identified among all six isolates in our investigation, causing low-level quinolones resistance. 25
DNA hybridization showed that blaNDM-1 was located on a plasmid of 55kb. As expected, resistant plasmids carrying blaNDM-1 for six clinical isolates were successfully transferred into the recipient E. coli J53 through conjugation, further proving the transferable characteristic of the resistance determinants. Typing of plasmid incompatibility groups showed that IncN plasmids were the most common plasmid types in our study. Previous reports indicated that the IncN plasmid has been shown to encode clinically important resistance determinants among E. coli and also in Klebsiella pneumoniae isolates, such as blaCTX-M, blaNDM-1, and qnr.26–28 Highly efficient transmission of the IncN plasmids appeared to account for the diversity and worldwide spread of blaNDM-1-carrying Enterobacteriaceae. 29
We also characterized the two porin proteins in E. coli isolates, which were associated with resistance or reduced susceptibility to carbapenems. Only one band was detected by SDS-PAGE, which convinced to be OmpF. ompC sequences showed that all six isolates possessed four deletions of the amino acid within the conserved L4 loop-borne and various substitutions. We further analyzed envZ and ompR, which were the main regulator of ompC 30 and no mutations in both envZ and ompR. There was no mutation detected in the promoter of the ompC gene either. Thus, we supposed that these substitutions and deletions of the ompC sequences might modify the molecular weight of the OmpC proteins and cause OmpC concealed by OmpF, which were indicated with the changes of the permeability in E.coli cells, contributing to multidrug resistance in E. coli. 9
MLST manifested that five of six isolates belonged to ST2, which differs from the international high-risk clone ST131 of carbapenems resistance E. coli. 1 ST2 was closely related to Shiga toxin-producing E. coli O157: H7, while our study did not draw such a conclusion.31,32 Furthermore, PFGE revealed that the four of six isolates shared the same genotype, which may create the dissemination of NDM-1 in healthcare facilities.
In conclusion, the resistance-associated genes and the loss of the outer membrane are contributing to the complex phenotype of these NDM-1-producing E. coli isolates. Both horizontal transfer of IncN and transmission of ST2 were responsible for the spread of drug resistance. Thus, our study emphasized an urgent enhancement of infection control directives to control the transmission of XDR E. coli, and the epidemiological surveillance is indispensable for implementing appropriate intervention measures.
Footnotes
Acknowledgments
This work was supported by research grants from the National Natural Science Foundation of China (no. 81171614), the Health Department of Zhejiang Province of the People's Republic of China (no. 2011KYA106), and the Zhejiang Provincial Program for the Cultivation of High-Level Innovative Health talents.
Disclosure Statement
No competing financial interests exist.