
Abstract
Staphylococcus aureus is a predominant pathogen in keratitis, and the rate of methicillin-resistant S. aureus (MRSA) is increasing. In our previous study, genotypes of MRSA isolates from keratitis cases were classified into ST5 or ST764 lineage by multi-locus sequence typing. In this study, we examined the virulence properties of these MRSA keratitis isolates and its virulence determinants. There was no difference in the prevalence of virulence genes, such as adhesion and toxins, between ST5 and ST764 isolates. All ST5 isolates carried the intact psm-mec gene, which suppresses exotoxin production and colony spreading, but promotes biofilm formation. In contrast, all ST764 isolates had one point mutation in the psm-mec gene. Biofilm production in ST5 isolates was significantly higher than that in ST764 isolates, whereas colony spreading, hemolytic activity, and production of alpha-phenol-soluble modulins were higher in ST764 than in ST5 isolates. The toxicity of ST764 supernatants to corneal epithelial cells was higher than that of ST5 supernatants. These results suggest that the point mutation in the psm-mec gene contributes to the difference in virulence properties between ST5 and ST764 isolates in MRSA keratitis.
Introduction
I
Antibiotic-resistant S. aureus initially developed in hospitals but have since spread to the community, where rates of methicillin-resistant S. aureus (MRSA) are now approaching those of hospitals.3,4 MRSA can cause keratitis as a serious complication of ocular surface disease or surgery.5–9 Khan et al. reported that MRSA isolated from cases of keratitis possesses types II and III SCCmec elements, which are also detected in hospital-acquired MRSA (HA-MRSA). 10 In contrast, Hsiao et al. reported the isolation of both community-acquired MRSA (CA-MRSA) and HA-MRSA from ocular infections, including keratitis. 11
Previously, we examined the genotype of S. aureus isolated from ocular sites of cases in Japan by multi-locus sequence typing (MLST), and ST5 SCCmecII and ST764 SCCmecII clones were predominant among MRSA isolates from keratitis. 12 ST5 and SCCmec type II elements are among the most globally disseminated HA-MRSA lineages.13,14 The ST764 clone, a variant of the ST5 HA-MRSA lineage with the arginine catabolic mobile element (ACME), was isolated from children in a community in Niigata, Japan. 15 There are few reports about the virulence of MRSA isolates from keratitis, and little is known about the differences between ST5 and ST764 in terms of virulence against corneal tissue.
In this study, bacteriological profiles of MRSA isolates (ST5 and ST764) from keratitis cases were examined. Furthermore, bacterial virulence and influences on corneal tissue were investigated.
Materials and Methods
Patients and bacterial isolates
Ten MRSA stock isolates from keratitis cases were used. Five were obtained from Ehime University Hospital, from 2003 to 2011, and the other five were collected from various locations in Japan in 2003 via a Japanese multi-center study. 16 In our previous study, we performed MLST analysis and SCCmec typing and identified five isolates each of ST5 SCCmec type II and ST764 SCCmec type II strains. 12 The clinical records of the MRSA keratitis cases were reviewed, and we collected data on age, gender, and systemic and local predisposing factors. This study was approved by the Ehime University Review Board and conformed to the tenets of the Declaration of Helsinki.
Cell culture
In this study, we used Simian virus 40-immortalized human corneal epithelial cells (HCECs). The cells were maintained in keratinocyte serum-free medium (KSM; Invitrogen, Carlsbad, CA) in a humidified 5% CO2 incubator at 37°C. Before the initiation of the toxicity tests, HCECs (2 × 104 or 1 × 105/well) were cultured to 95% confluence in 24-well plates.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was performed using a broth microdilution assay, as described by the Clinical and Laboratory Standards Institute (CLSI; CLSI M7-A8). 17 The minimum inhibitory concentration (MIC) was determined using dry plates (Kyokuto Pharmaceutical Industrial Co., Ltd, Tokyo, Japan) with Mueller–Hinton broth (cation-adjusted). After a 24-h incubation at 37°C in CO2, an ellipse appeared on the MIC value scale (μg/ml) at the concentration at which the antibiotic tested inhibited bacterial growth. All results were interpreted using cutoff values for susceptibility and resistance according to criteria for Staphylococcus spp. established by the CLSI (CLSI M100-S23). 18 The antimicrobial agents included imipenem (IPM), arbekacin (ABK), vancomycin (VCM), teicoplanin (TEIC), linezolid (LZD), gentamicin (GM), clarithromycin (CAM), clindamycin (CLDM), minocycline (MINO), levofloxacin (LVFX), rifampicin (RFP), and sulfamethoxazole/trimethoprim (SXT). Because there is no established cutoff value for ABK, the value for GM was used for ABK.
Bacterial typing method
Isolates were typed using the DiversiLab typing system (DL; bioMérieux, Inc., Durham, NC), which uses the DNA repetitive elements present in the organism's genome to determine the genetic relatedness of bacterial isolates, 19 as recommended by the manufacturer. Following DNA extraction, repetitive-element polymerase chain reaction (PCR) was performed using the DL Staphylococcus kit, and the amplification products were analyzed using DL with microfluidics chip technology. The DL data for each organism consisted of a dendrogram, graph of fluorescence intensity corresponding to the banding pattern, and similarity matrix. DL strain relatedness was defined as a minimum of >95% similarity with a difference of up to one band in the dendrogram. Along with DL, the POT methodology, which comprises genotyping based on multiplex PCR, was performed. The Cica Geneus Staph POT kit (Kanto Chemical, Tokyo, Japan) was used according to the manufacturer's instructions. 20
Genetic virulence determination
The presence of genes coding for virulence factors, such as adhesion molecules and toxins, were examined by PCR. We evaluated genes encoding enterotoxins (sea and seb), exfoliative toxins (eta and etb), toxic shock syndrome toxin 1 (TSST) (tst), icaA (icaA), fibronectin-binding protein A (fbnA), collagen-binding protein (cna), and the ACME arcA and ACME opp-3C. Each gene was identified by simplex PCR amplification using primers described elsewhere.21–24 Along with the detection of virulence genes, DNA fragments including the psm-mec, which is located in the mobile genetic element SCCmec-II and suppresses production of alpha-phenol-soluble modulins (PSMαs), was sequenced using primers as reported previously. 25
Phenotypic virulence determination
Phenotypic virulence, such as biofilm production, colony spreading, and haemolytic activity, were examined. The degree of biofilm production was determined by microtiter plate assay, as described previously. 26 Bacteria were inoculated into tryptic soy broth (TSB) with 0.25% glucose (Wako Pure Chemical Industries) and incubated overnight. This solution was diluted 1:100 in TSB with glucose, and 200 μL were dispensed into 96-well polystyrene microtiter plates. The plates were incubated overnight at 37°C under aerobic conditions. The plates were then washed thrice with phosphate-buffered saline and stained with 0.1% crystal violet (Wako Pure Chemical Industries). The crystal violet was solubilized using 30% glacial acetic acid for 15 min. Relative biofilm formation was assayed by reading the optical density at 570 nm. The biofilm assay was assessed in three independent experiments, each performed in triplicate.
The colony spreading assay was performed as described previously. 27 S. aureus overnight culture (2 μl) was spotted onto soft TSB agar plates containing 0.24% agar, and incubated for 24 hr at 37°C. The diameter of the giant colony was measured. Experiments were performed in triplicate.
Hemolytic activity was measured as described previously, 28 except that sheep blood was used instead of rabbit blood. Isolates were cultured for 16 hr at 37°C in yeast extract-casamino acid-sodium pyruvate broth. The cultures were then centrifuged (1,509 g, 5 min, room temperature), and the supernatant was filtered using a 0.45-μm pore size membrane filter (DISMIC-25CS; ADVANTEC). The culture supernatant was diluted several fold, mixed with 1.5% v/v erythrocyte solution, and incubated at 37°C for 1 hr. Then, the cells were removed, and the OD450 of the supernatant was measured. Hemolytic activity was assessed in three independent experiments, each performed in triplicate.
The amount of PSMαs was measured as previously described. 18 Briefly, MRSA culture supernatants were evaporated and the remaining solid was dissolved in 40% acetonitrile. The supernatants remaining after subsequent centrifugation were evaporated, and the evaporated products were dissolved in water and subjected to reversed-phase high-performance liquid chromatography. PSMαs (PSMα1 + PSMα2 + PSMα3 + δ-hemolysin) eluted from 12 to 17 min were measured by a photodiode array detecter (5430 Diode Array Detector; Hitachi).
Bacterial challenge to HCECs
Bacterial adhesion and internalization assays were performed essentially as described previously. 29 Briefly, for the determination of adhesion, bacteria (mid-log phase or stationary phase) were washed twice with culture media and then applied to HCEC monolayers (∼108 CFU in 1 ml medium). Following incubation for 1 hr, epithelial cell cultures were washed and disaggregated, and adherent bacteria were counted by serial dilution. To determine internalization, bacteria were added to the cells as above, and following incubation for 1 hr, cells were washed, covered with medium containing 100 μg/ml gentamicin, and incubated for an additional 1 hr to kill residual extracellular bacteria, before epithelial cell disruption and the enumeration of bacteria. The following equations were used for estimating adhesion and internalization.
Growth index = total bacteria at harvest/number of bacteria added initially
% Adhesion = (attached bacteria/number of bacteria added initially) ×100
Relative adhesion = (% experimental adhesion/% adhesion in control)
% Invasion = (number of bacteria internalized/number of bacteria added initially) ×100
Corrected invasion = % invasion/growth index
Relative invasion = (corrected invasion of experimental/corrected invasion of control) ×100
Five samples were run for each group, and experiments were performed in duplicate.
The toxicity of bacterial supernatants to corneal epithelial cells was examined. Aliquots (100 μl) of the supernatant were mixed with 900 μl KSM and added to the HCECs in 24-well microtiter plates. After incubation for 20 hr in a humidified 5% CO2 incubator at 37°C, cell viability was quantified using the LDH assay (Promega). Lysis buffer was used as a positive control of cell damage. Cell killing (%) was expressed as (optical density of tested sample/optical density of positive control) ×100. Experiments were performed thrice independently, each in triplicate.
Statistical analysis
We compared the prevalence of the virulence factors between ST5 and ST764 using the Student's t-test. A p-value of <0.05 was considered to indicate statistical significance. Data are shown as mean ± SD unless otherwise specified.
Results
Patient backgrounds
Patient backgrounds and the ST type of the isolates are shown in Table 1. The age of the patients ranged from 29 to 86 years (mean age, 64.4 ± 19.2 years). There were four males and six females. A predisposing systemic or ocular condition was identified in all patients (Table 1). Seven patients had an ocular predisposing factor; common ocular conditions were keratoplasty (four patients) and bullous keratopathy (two patients). Systemic risk factors were identified in five patients, with diabetes mellitus being present in three. Three patients with keratitis caused by ST5 had hospital associated (HA) factors because they had a history of dialysis or hospitalization due to brain infarction within 1 year of a MRSA culture date. Thus, patients with ST5 MRSA keratitis appeared to have more HA factors.
DALK, deep anterior lamellar keratoplasty; DM, diabetes mellitus; HIV, infection of human immunodeficiency virus; HT, hypertension; LKP, lamellar keratoplasty; PKP, penetarating keratoplasty.
Antimicrobial susceptibility testing
The antimicrobial susceptibilities and MICs of the drugs evaluated for each strain are summarized in Table 2. All strains were susceptible to ABK, VCM, TEIC, LZD, RFP, and ST and were resistant to IPM and CAM. Two of the ST5 strains showed susceptibility to GM and MINO, and one ST5 strain was susceptible to CLDM. All ST764 strains and one ST5 strains showed high resistance to LVFX, with an MIC >256 μg/ml.
Strains 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 were isolated from cases 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10, respectively.
This interpretation was determined according to the Clinical and Laboratory Standards Institute (M100‐S23).
ABK, arbekacin; CAM, clarithromycin CLDM, clindamycin; GM, gentamicin; I, intermediate; IPM, imipenem; LVFX, levofloxacin; LZD, linezolid; MIC, minimum inhibitory concentration; MINO, minocycline; R, resistant; RFP, rifampicin; S, susceptible; SXT, sulfamethoxazole/trimethoprim; TEIC, teicoplanin; VCM, vancomycin.
Bacterial typing method
DL typing demonstrated that all isolates belonged to one cluster, defined as having >95% similarity (Fig. 1). The POT type of seven strains (three of ST5 and four of ST764) and three strains (2 of ST5 and 1 of ST764) were 93-255-127 and 95-255-127, respectively. Neither DL nor POT typing could distinguish ST5 and ST764.
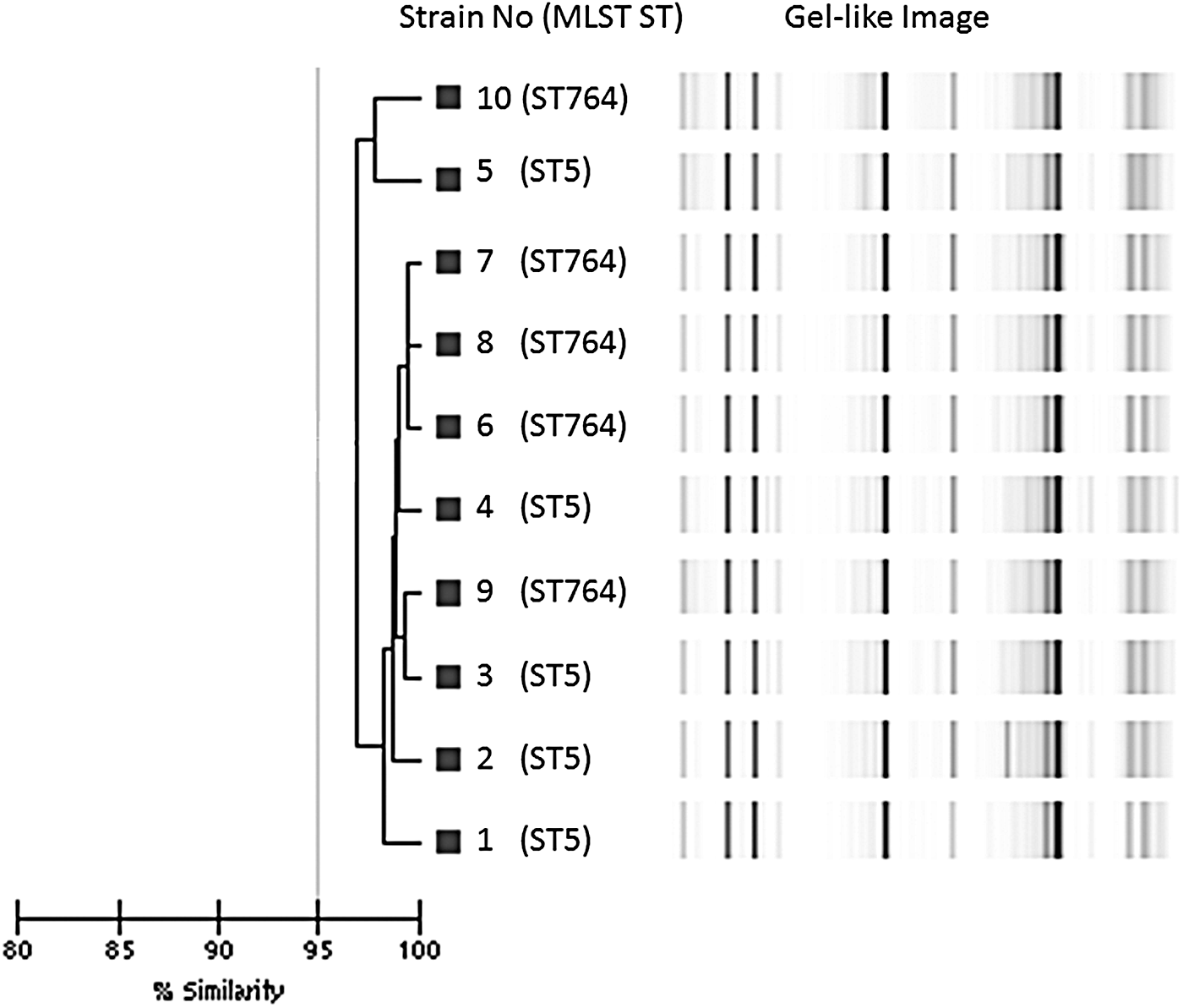
Dendrogram showing ST5 and ST764 isolates. Gel-like images were created using the DiversiLab system. The horizontal bar at the bottom left of the dendrogram indicates the similarity coefficient percentages of the isolates. ST5 and ST764 isolates were clustered together at >95% similarity. MLST, multi-locus sequence typing.
Genetic virulence determination
Genes encoding virulence factors in the 10 isolates are shown in Table 3. All isolates were positive for sea, seb, and fbnA. All ST5 isolates and three ST764 isolates had the tst gene. Two ST5 isolates and three ST764 were positive for eta. Three ST5 isolates and four ST764 were positive for icaA. All isolates were negative for etb, cna, and ACME opp-3. One strain each of ST5 and ST764 had the ACME arc gene. Gene sequencing data in the psm-mec promoter showed that all ST764 isolates had a point mutation (-7T>C). Four ST5 isolates had intact psm-mec promoters, and one had a T inserted between bases -74 and -75 of the psm-mec open reading frame.
Strains 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 were isolated from cases 1, 2, 3, 4, 5, 6, 7, 8, 9, and 10, respectively.
ACME, arginine catabolic mobile element.
Phenotypic virulence determination
Biofilm-forming ability was determined quantitatively by microtiter plate assay (Fig. 2a). The mean ODs of the ST5 and ST764 isolates were 0.2429 ± 0.2781 and 0.0521 ± 0.0375, respectively. ST5 isolates produced significantly more biofilm than did ST764 (p ≤ 0.001). Colony spreading was measured on soft agar (Fig. 2b). The mean colony diameters of ST5 and ST764 isolates were 21 ± 10.4 and 37 ± 9.2 mm, respectively, with a significant difference (p < 0.001). Hemolytic activity of the isolates was determined to be significantly higher in ST764 than ST5 isolates (p < 0.05; Fig. 2c). PSMαs production by ST764 isolates was three-fold higher than that by ST5 isolates (p < 0.01; Fig. 2c)
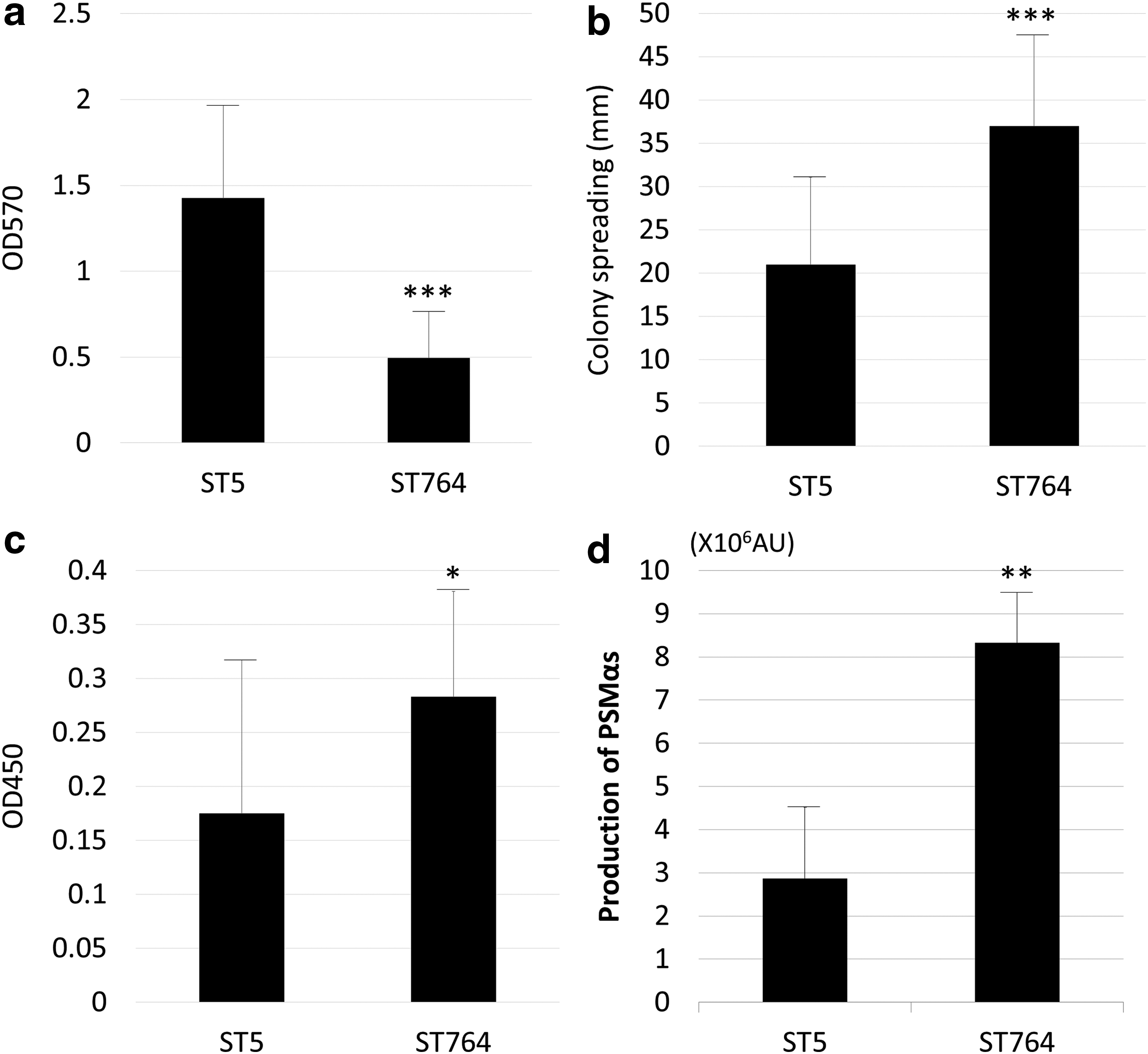
Phenotypic virulence of ST5 and ST764 isolates.
Bacterial adhesion and internalization to HCEC were examined (Fig. 3a, b). There was no difference in adhesion or internalization between ST5 and ST764 isolates. The toxicity of bacterial supernatants to HCEC was examined (Fig. 3c). The mean toxicity (%) of ST764 isolate supernatant was significantly higher than that of ST5 isolate supernatant (p < 0.05).

Bacterial challenge to HCECs.
Discussion
MRSA is not a common pathogen causing keratitis but could become more common due to the increasing rate of MRSA in the ophthalmic field in general. 30 Furthermore, antibiotics are used for ocular surface disease and ocular surgery, so commensal MRSA could be selected in eye lids and conjunctival sacs and increase the chance of keratitis. MRSA was traditionally associated with healthcare facilities, but its prevalence has reportedly increased in otherwise healthy patients without identified risk factors. Methicillin resistance in S. aureus is on the rise in the community, in large part due to proliferation of the infectious USA300 lineage. 4 The virulence of CA-MRSA against human tissues is higher than that of HA-MRSA, because CA-MRSA have virulence determinants such as alpha-toxin and PSMαs, the acquisition of PVL, which can destroy white blood cells and cause extensive tissue necrosis, and ACME, which facilitates stable skin colonization.21,31
In this study, we focused on ST5 and ST764 isolated from keratitis, which are predominant strains in keratitis. In a previous study, we demonstrated that MRSA isolates from keratitis did not possess the PVL gene, but we did not investigate other virulence factors. 12 As described previously, ST5 SCCmec type II is a worldwide-prevalent HA-MRSA clone and was dominant in Tokyo in 1992. 32
Recent studies have demonstrated that ST764 was found in hospitals and in the community in Japan.15,33–36 First, strains with ST764 SCCmecII were isolated from children in the community.15,35 Moreover, strains with ST764 clones were isolated from outpatients in Northern Japan. 33 Nakaminami et al. reported that ST764 clones were isolated from tertiary care hospitals in Tokyo, Japan. 34 Although ST764 is a hybrid variant of ST5, ST764 has been reported to have characteristics different from those of ST5.
Takano et al. demonstrated that ST764 strains from the community carry the ACME arc cluster next to their type II SCCmec element. 36 However, ACME-negative ST764 clones were isolated from hospitals in Tokyo. 34 Moreover, Kawaguchiya et al. showed that the ACME-arc gene was detected in not only ST764 but also ST5 isolates from outpatients. 33 Thus, the prevalence of the ACME gene does not seem to be a unique characteristic of ST764. Indeed the ACME-arc gene was detected in one ST5 and one ST764 isolate in this study. Takano et al. used ST764 isolates including isolates from healthy hands of medical students. 37 Different background of isolates may be related to differences between their and our results. We assessed MRSA keratitis isolates from various areas in Japan. Although all patients had ocular and systemic predisposing factors, patients with ST5 MRSA keratitis appeared to have more HA factors, rather than patients with ST764 MRSA keratitis. ST764 isolates may spread to not only hospitals but also the community.
We examined and compared antibiotic susceptibility of both ST5 and ST764 strains. Two ST5 but no ST764 strains were sensitive to MINO. All ST764 strains and one ST5 strain showed high resistance to LVFX, with a MIC >256 μg/ml. The susceptibility of other antibiotics against ST5 and ST764 were similar. High resistance to fluoroquinolones could be one characteristic of ST764 isolated from keratitis. In the ophthalmic field, fluoroquinolone eye drops have been widely used for the treatment and prevention of ocular infection. 37 Thus, it is possible that ST764 could be selected after fluoroquinolone eye drop use and could cause keratitis.
The presence of genes coding virulence factors was evaluated. The TSST-1 gene, which had been frequently found in ST5 strains, was found in all ST5 and three ST764 strains. There was no gene found only in ST5 or ST764 strains. However, all ST764 strains, but no ST5, had a mutation in the psm-mec promoter (-7T<C). The psm-mec gene is located in the mobile genetic elements SCCmec-II and–III of HA-MRSA, and transcription products of psm-mec suppress the production of PSMαs, which is a cytolytic toxin that contributes to CA-MRSA virulence in vitro.25,38 PSMαs consist of 20–25 amino acids and play a key role in MRSA evading the innate immune system. 39 PSMαs could contribute to infections caused by CA-MRSA. Furthermore, psm-mec RNA inhibits translation of the agrA gene, encoding a quorum sensing and positive transcription factor for the PSMα gene, via specific binding to agrA mRNA. 38 Both the transcription and translation products of the psm-mec promoted biofilm formation and suppressed colony-spreading activity, which is suspected to play a role in its virulence. 25
Recent studies demonstrated that HA-MRSA isolates with a same mutation in the psm-mec promoter (-7T<C) produced high amounts of PSMα, comparable to that of CA-MRSA.38,40 Thus, a mutation in the psm-mec promoter apparently could increase virulence in HA-MRSA. Indeed, our results demonstrated that colony spreading, hemolytic activities, and PSMαs production of ST764 isolates were higher than those of ST5 isolates, and that the toxicity of ST764 supernatants to corneal cells was stronger than that of ST5. Furthermore, biofilm production in ST5 isolates was higher than that in ST764 isolates.
Regardless of the presence or absence of virulence factor genes, biofilm production decreased while cell toxicity increased in ST764. This indicates that a mutation in the psm-mec promoter attenuated its function and increased the virulence of MRSA isolated from keratitis. One ST5 strain showed insertion of a T between bases -74 and -75 in the psm-mec gene, and this insertion apparently did not influence the function of the psm-mec gene. There was no difference in bacterial adhesion or internalization between ST5 and ST764 isolates, such that the mutation in the psm-mec promoter apparently also did not affect bacterial adhesion or internalization in corneal cells. Many factors, such as wall teichoic acids and fibronectin-binding protein, play key roles in the adhesion and internalization of S. aureus in corneal epithelial cells,29,41 and these factors might not be regulated by the psm-mec gene.
There could be differences in virulence and antibiotic susceptibilities between ST764 and ST5 strains. Thus, it is important to identify ST764 and ST5 strains using a simple method. Commercial bacterial typing methods, such as DL and POT methods, could not differentiate between ST764 and ST5. Sequencing of the psm-mec promoter could provide information about virulence.
The results of our study should be interpreted with caution, considering its limitations. First, the sample number was small because MRSA keratitis is rare. Second, clinical manifestations of keratitis caused by ST5 and ST764 strains were retrospectively examined and they were not documented. But clinical records described 4 cases with keratitis caused by ST5 and one case with keratitis caused by ST764 as “mild.” All cases with keratitis caused by ST764 were treated with vancomycin or arbekacin ophthalmic solution, but two cases with keratitis caused by ST5 needed vancomycin ophthalmic solution. Since outcomes in 5 patients were not followed, the differences in outcomes of keratitis caused by ST5 and ST764 isolates were not known. Thus, further investigations involving many cases are needed to understand the relationships between MRSA virulence and clinical features.
In conclusion, ST764 isolates may have higher fluoroquinolone resistance and virulence compared with ST5 isolates. Mutation of the psm-mec promoter may contribute to the virulence of ST764 isolates in MRSA keratitis.
Footnotes
Acknowledgments
We thank Shinobu Murakami for their valuable technical assistance with our experiments and providing isolates. This study was supported by the Department of Bioscience, INCS, Ehime University.
Disclosure Statement
No competing financial interests exist.