
Abstract
Candida spp. cause significant health problems, inducing various types of superficial and deep-seated mycoses in humans. As a result of the increasing antibiotic resistance among pathogenic yeasts, the interest in alternative agents of antifungal activity is growing. This study evaluated the antimicrobial activity of selected essential oils (EOs) against Candida clinical and food-borne strains, including antibiotic-resistant isolates, in relation to yeast cell surface hydrophobicity (CSH). Candida strains showed different range of susceptibility to tea tree, thyme, peppermint, and clove oils, and peppermint oil demonstrated the lowest anticandidal activity with minimal inhibitory concentrations (MICs) of 0.03–8.0% v/v. MIC values for thyme and clove oils ranged from 0.03% to 0.25% v/v, and for tea tree oil—from 0.12% to 2.0% v/v. The exception was Candida tropicalis food-borne strain, the growth of which was inhibited after application of EOs at concentration of 8% v/v. Due to diverse yeast susceptibility to EOs, isolates were divided into five clusters in a principal component analysis model, each containing both clinical and food-borne strains. Hydrophobic properties of yeast were also diversified, and 37% of clinical and 50% of food-borne strains exhibited high hydrophobicity. The study indicates high homology of clinical and food-borne Candida isolates in relation to their susceptibility to anticandidal agents and hydrophobic properties. The susceptibility of yeasts to EOs could be partially related to their CSH. High antifungal activity of examined EOs, also against antibiotic-resistant isolates, indicates their usefulness as agents preventing the development of Candida strains of different origin.
Introduction
D
Currently, there are three classes of antifungal antibiotics available for treatment of serious Candida infections: the azoles (fluconazole, itraconazole, voriconazole, and posaconazole), the echinocandins (anidulafungin, caspofungin, and micafungin), and the polyenes (nystatin and amphotericin B). However, these antifungal agents have some limitations, such as low susceptibility of some Candida species or strains, Candida resistance and cross-resistance developing during prolonged and prophylactic treatment, significant cost, toxicity, drug interactions, or lack of an oral formulation.5,6
Resistance to antibiotics varies widely by country and species, and overall reported resistance is highest in South Africa (50%) and lowest in the Republic of Korea (0.9%). 7 Based on published data and projections, costs associated with prolonged length of hospitalization of patients with Candida infections and healthcare expenditures are estimated at 8 billion US dollars every year in US.7,8
The term candidiasis refers to both more superficial and milder diseases, and serious infections, including blood stream infections and disseminated candidiasis, whereas the description of invasive candidiasis concerns severe diseases such as candidemia, endocarditis, disseminated infections, central nervous system infections, endophthalmitis, and osteomyelitis. 4 The list of factors increasing the risk of candidiasis encompasses underlying malignancies, immunosuppressive diseases, the use of wide-spectrum antibiotics or corticosteroids, invasive interventions and transplantations, internal prosthetic devices, aggressive chemotherapy, and parenteral alimentation.2,4,8
However, one of the determinants of Candida pathogenicity is cell surface hydrophobicity (CSH), a factor closely related to the adhesion properties of yeast.9,10 A positive correlation between the CSH and candidal adhesion to buccal epithelial cells and acrylic surfaces has been previously reported.9,11 CSH is also considered a feature associated with biofilm formation and biofilm biomass. 10
The list of Candida species causing serious, life-threatening infection increases every year. Besides C. albicans, at least 14 Candida species were isolated from patients as infectious agents, namely C. glabrata, C. tropicalis, C. parapsilosis, C. krusei, C. guilliermondii, C. lusitaniae, C. dubliniensis, C. pelliculosa, C. kefyr, C. lipolytica, C. famata, C. inconspicua, C. rugosa, and C. norvegensis.7,10 Interestingly, it has been reported that the incidence of C. albicans has decreased, while that of the non-albicans Candida has increased.4,12
The aim of this research was to evaluate Candida spp. susceptibility to selected essential oils (EOs), natural substances of plants origin with known antifungal activity, in relation to yeast CSH.
Materials and Methods
Yeast
In this study, 45 clinical and food-borne Candida isolates were tested, including Candida albicans strain ATCC 10231 used as a reference for analysis of antifungal agents (Table 1).
The clinical isolates were obtained from Department of Laboratory Diagnostics of Polish Mother's Memorial Hospital Research Institute in Lodz, Poland. All yeast isolates were identified using the API 20 C AUX tests (BioMerieux) as described previously. 13
Essential oils
The susceptibility of Candida strains was estimated for EOs of Melaleuca alternifolia (Maiden & Betche) Cheel (tea tree oil), Mentha piperita L. (peppermint oil), Thymus vulgaris L. (thyme oil), and Syzygium aromaticum (L.) Merr. & L.M. Perry (clove oil).
Chemical composition of EOs was analyzed by GC-MS-FID using Trace GC Ultra (Thermo Scientific) combined with DSQ II mass spectrometer and with flame ionization detector (FID) throughout MS-FID splitter (SGE; Analytical Science). Analysis was provided using nonpolar capillary column Rtx-1 ms (60 m × 0.25 mm, film thickness 0.25 μm; Restek). The oven temperature was programmed as follows: 50°C–300°C at 4°C/min; injector temperature 280°C; detector temperature 310°C; carrier gas helium with regular pressure 200 kPa; and ionization energy 70 eV, ion source temperature 200°C. Identification of components was based on the comparison of their mass spectra with those in a laboratory-made MS library, commercial libraries (NIST 09, Wiley 275.1, Mass Finder 4) along with the retention indices associated with a series of alkanes with linear interpolation (C8–C26). A quantitative analysis (expressed as percentages of each component) was carried out by peak area normalization measurements without correction factors. The chemical composition of EOs is presented in Table 2.
RI, retention index.
Determination of minimal inhibitory concentration of EOs
The antifungal activity of EOs was determined with the use of broth macrodilution method, according to CLSI reference method. 14 In detail, a range of twofold dilutions of EOs from 16% to 0.0075% (v/v) in the Sabouraud broth was prepared. To enhance the EO solubility, Tween 80 was included at a final concentration of 0.1% (v/v). Tubes containing only Tween 80 (without EO) were used as control. Yeast inocula were prepared by growing isolates on Sabouraud agar for 24–48 hr at 35°C and then suspending growth in sterile saline (0.85% NaCl solution). The number of cells was determined by counting in a hemacytometer. Each tube was inoculated with suspensions of Candida spp. cells (106 CFU/ml) in saline. Following 24 hr of incubation at 37°C, the tubes were examined for visible yeast growth as evidenced by turbidity. The minimal inhibitory concentration (MIC) was assessed as the lowest concentration of the EO inhibiting visible growth of yeast compared to positive control cultures without the addition of EOs.
Cell surface hydrophobicity
The hydrophobicity of Candida strains was determined by microbial adhesion with a hydrocarbon method, previously described.
15
The tested isolates were grown overnight in the Sabouraud dextrose broth at 28°C and washed with phosphate-buffered saline (NaCl 8 g/L, KCl 0.2 g/L, Na2HPO4 1.44 g/L, KH2PO4 0.24 g/L; pH 7.4). For adhesion assay, 2.5 ml of the cell suspensions (OD520 = 1.0) were mixed with 0.5 ml of p-xylene (Sigma-Aldrich) in an acid-washed glass tube. The samples were incubated at 37°C for 10 min and vigorously mixed for 30 sec. After 45 min of incubation at 37°C, the absorbance of aqueous phase was measured at 520 nm. The percentage of cells in the xylene layer was used to estimate the hydrophobicity index (HI), according to the following formula:
where A1, absorbance of inoculum; A2, absorbance of the aqueous phase.
Statistical analysis
A principal component analysis (PCA) model was generated based on the data of MIC values of EOs for 45 clinical and food-borne Candida isolates, to group the tested yeasts in a statistically significant manner (Statistica 10; Stat Soft, Inc.). Hierarchical cluster analysis was used to estimate the similarity of Candida strains according to their CSH and the results were presented as HIs of three replicates. The algorithm for hierarchical clustering was calculated by Ward's method and cluster hierarchy was presented using a tree structure (dendrogram) with horizontal agglomeration distance (Statistica 10; Stat Soft, Inc.).
Results
Candida clinical and food-borne isolates were evaluated for their susceptibility to four EOs (tea tree, thyme, peppermint, and clove) and grouped on the basis of MIC values, using the PCA model. Due to diverse yeast sensitivity to EOs, isolates were divided into five clusters (Fig. 1). Cluster I contained two clinical C. albicans and four food-borne strains comprising C. lusitaniae, C. krusei, and C. parapsilosis, characterized by low sensitivity to the tested EOs, and highest MIC value for peppermint oil equaled 8.0%. However, the lowest sensitivity to the EOs tested showed C. tropicalis fo/BM/01 with all MIC values of 8.0%. In cluster II, five clinical C. albicans, one C. famata food-borne, and two C. lusitaniae (one clinical and one food-borne) isolates were grouped, all more sensitive to thyme, clove, and peppermint oils than yeasts in cluster I. Strains in cluster III (two clinical C. albicans, one clinical C. glabrata, two food-borne C. boidinii, one food-borne C. krusei) demonstrated the highest sensitivity to tea tree (MICs 0.12–0.5%) and thyme (MICs 0.03–0.12%) oils among all the tested yeasts (Fig. 1). However, isolates in cluster IV (three clinical C. albicans, and six food-borne isolates: C. krusei, C. lusitaniae, C. pelliculosa, C. famata, C. rugosa, C. boidinii) were the most sensitive to peppermint and clove oils, with MIC ranges 0.03–0.25% and 0.03–0.12%, respectively.

Spatial scattering of Candida spp. isolates on the basis of their susceptibility to different essential oils; PCA model. PCA, principal component analysis.
Cluster V exhibited the greatest differentiation of yeast susceptibility to tea tree oil (MICs in the range 0.12–1.0%) and thyme oil (0.03–0.25%). This cluster encompassed most of all C. albicans clinical isolates (n = 10), but also two food-borne isolates, identified as C. krusei and C. colliculosa. Between clusters II, III, IV, and V were located C. albicans ATCC 10231, cl/MP/8K, and cl/MP/2K; isolates demonstrated rather low sensitivity to tea tree oil with MIC ranged from 1.0% to 2.0%.
In our previous study of all tested strains, 7% was resistant to nystatin (C. albicans cl/MP/01, C. tropicalis fo/BM/01, C. pelliculosa LOCK 0007), 32% to fluconazole (C. albicans cl/MP/1M, cl/MP/01, cl/MP/04, cl/MP/05, cl/MP/08, cl/MP/09, C. glabrata cl/KL/01, C. boidinii fo/MP/03, C. rugosa fo/BG/05, C. tropicalis fo/BM/01), 23% to voriconazole (C. albicans cl/MP/1M, cl/MP/01, cl/MP/04, cl/MP/05, cl/MP/08, C. glabrata cl/KL/01, C. tropicalis fo/BM/01), and no strains grew in the presence of caspofungin. 13 Moreover, the group of strains showing resistance to antibiotics included both clinical and food-borne Candida isolates to be inhibited. The susceptibility of these antibiotic-resistant strains to EOs also varied (Table 3). Generally, the more antibiotics to which the strains were resistant, the lower the sensitivity of these isolates to EOs. Peppermint oil was found to be the one with the lowest antifungal activity against antibiotic-resistant yeasts and in the case of three strains (fluconazole- and voriconazole-resistant C. albicans, fluconazole-, voriconazole-, and nystatin-resistant C. albicans, and C. tropicalis) inhibited their growth at the concentration of 8.0%.
Flu, fluconazole; MIC, minimal inhibitory concentration; nys, nystatin; R, resistant; S, susceptible; vor, voriconazole.
Furthermore, antibiotic-resistant food-borne strains required lower concentrations of EOs than antibiotic-resistant clinical isolates. The specific exception was food-borne C. tropicalis fo/BM/01, which showed resistance to fluconazole, voriconazole, and nystatin and was susceptible to the EOs at concentration of 8.0% (Table 3). Moreover, in the group of antibiotic-sensitive yeasts, non-albicans isolates were more sensitive to tea tree oil than C. albicans with MIC ranges of 0.12–1.0% and 0.25–2.0%, respectively.
Hierarchical cluster analysis applied for similarity assessment of Candida isolates according to their CSH showed differentiation of the strains in the five main clusters (Fig. 2). Although this differentiation did not coincide exactly with the spatial distribution of strains in the PCA analysis, the C. tropicalis fo/BM/01 was similarly separated from the other isolates. This strain was characterized by resistance to fluconazole, voriconazole, and nystatin and the lowest sensitivity to the EOs among all Candida strains tested. Furthermore, the strain showed the lowest CSH with an HI of 22.4% ± 2.6%.
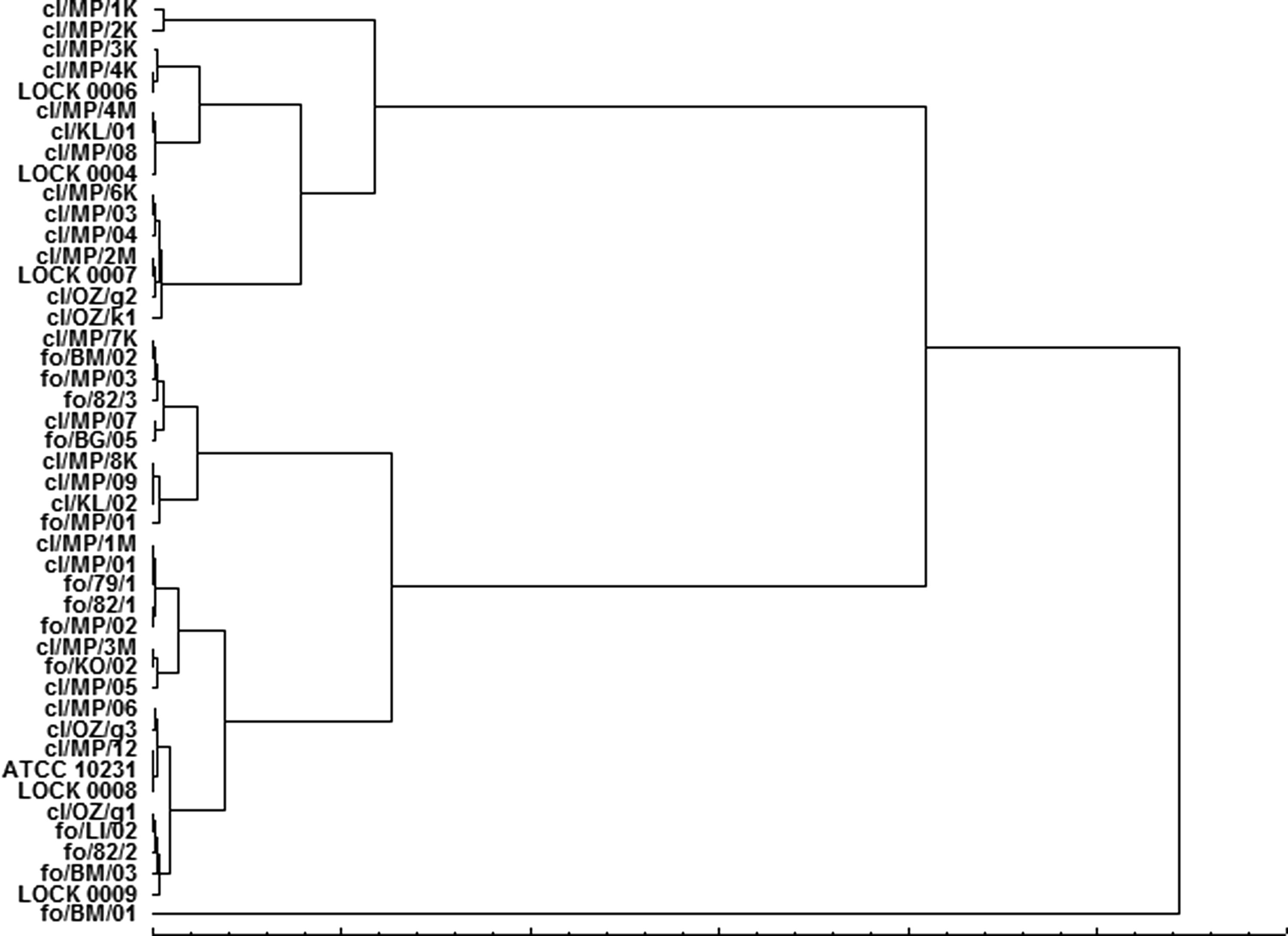
Dendrogram of similarity of hydrophobicity indexes determined for Candida spp. isolates.
Comparable to the PCA model, strains of cluster I (beside C. parapsilosis fo/82/3) were grouped together and their HIs ranged from 70.0% to 72.9%. According to the shown moderate levels of CSH, yeast sensitivity to EOs was also rather low. Sequentially distributed isolates from cl/MP/7K to fo/MP/01 and cl/OZ/g3 to LOCK 0009 exhibited high hydrophobicity (75.2–92.3% clinical and 75.9–91.2% food-borne isolates) and diverse susceptibility to the EOs. All the clusters included both, the clinical and food-borne Candida isolates, which indicates a high homology of these strains, not only with regard to their susceptibility to antimycotics but also related to their hydrophobic properties.
Discussion
EOs are complex mixtures, which can comprise more than sixty compounds, however, major components can constitute up to 85% of the EO. 16 Due to the great number of constituents, EOs seem to have no specific cellular targets. 17 The antimicrobial activity of EOs results from their lipophilic properties and is associated with damage of cell wall and cytoplasmic membrane structures, and their increased permeabilization. 18 In eukaryotic cells, EOs can provoke depolarization of the mitochondrial membranes, affect ionic channels, and reduce the pH gradient. Moreover, permeabilization of outer and inner mitochondrial membranes leads to cell death by apoptosis and necrosis. 19 It have been demonstrated that various EOs or their main components can induce nuclear mutations and ultrastructural alterations in nucleus.17,20,21
Generally, the major components are found to reflect the biological features of the EOs, however, its activity is modulated by other minor molecules. 18 In addition, it seems that several components of EOs play a role in cell penetration, fixation on cell wall and membranes, and cellular distribution. 22 It is generally assumed that the mechanisms by which the constituents of EOs inhibit the growth of microorganisms may be partially dependent on their hydrophobicity.16,18 In this context, it cannot be excluded that the interaction between the EOs and the surface of yeast cell wall may also depend on the hydrophobic properties of yeasts. Our results partially confirm this hypothesis, proving the relationship between the lower HI and the lower sensitivity to EOs. However, the mechanism could be more complex than just the hydrophobic interactions. In the structure of C. albicans cell wall, several carbohydrates and proteins have been identified, which contribute to hydrophobic interactions and are potentially involved in the CSH of the organism. 23 These interactions include the ability of the yeast to adhere to epithelial and endothelial cell surfaces and plastic devices, to evade the action of phagocytic cells, and also to enhance the uptake of substances from the medium. 24 This phenomenon of increased intracellular incorporation of compounds in C. albicans cells of high hydrophobicity has been previously demonstrated for corticosterone, 24 and this mechanism may also act in the case of EOs or their components. It was also reported that Candida spp. surface hydrophobicity may be linked to coaggregation properties, 25 and the ability of yeasts to perform coaggregation and excrete an extracellular matrix contribute to cellular adherence to acrylic and polymeric surfaces and to higher antifungal resistance. 26 On the other hand, crucial can also be the liposoluble nature of EOs and their constituents, which facilitates their interaction with cellular structures containing lipid components. 27
To our best knowledge, no particular resistance or adaptation to EOs has been reported because of their mode of action affecting several targets at the same time. In accordance with that assumption, resistance of Candida strains tested to tea tree, thyme, peppermint, and clove oils has not been observed in this study. The lowest anticandidal activity was demonstrated for peppermint oil, which inhibited the growth of 15.6% of the strains, equally clinical and food-borne isolates, at a concentration of 8% v/v. The high homology between clinical and food-borne Candida strains concerns not only their sensitivity to EOs but also biochemical profiles and resistance to antibiotics, as we have demonstrated previously. 13
The development of resistance to conventional chemotherapeutic agents by Candida spp. seems to be a continuous process.3,5–7 Candida strains may show resistance to all groups of antibiotics, even to these belonging to a new class of echinocandin drugs. 28 In our study, all examined strains were sensitive to caspofungin, but seven isolates showed cross-resistance to fluconazole and voriconazole. 13 The scientific publications are focused on clinical strains rather, and we did not find reports of antibiotic resistance among food-borne Candida isolates. Among those examined by us, 20% food-borne strains showed resistance to antibiotics currently used in the treatment of candidiasis, and one strain was voriconazole-, fluconazole-, and nystatin-resistant. It should be emphasized that the occurrence of antibiotic-resistant Candida yeast in foods and in production environment may pose a threat to human health, as has been shown for clinical isolates.2,3,5–8,28 To date, antidrug resistance has been reported for food-borne bacterial pathogens and this finding is considered a public health issue. 29 There is increasing evidence that antimicrobials used in agriculture and livestock farming may act as select factors for resistant food-borne microorganisms.
The EOs have demonstrated potential as antifungal agents both when used solely and in combination with an antibiotic.30,31 Moreover, combining EOs with conventional antimycotics enables lower doses of antifungal agents to be administered. This finding opens perspectives not only in increasing activity of antibiotics when applied with EOs but also in their extended suitability due to lower rates of yeast resistance.
The broad spectrum of antimicrobial activity and relative lack of toxicity of EOs indicate numerous possible formulations for their topical use and for the preservation of crops or food products.18,32 EOs may constitute effective alternatives or complements to synthetic compounds, without showing the same collateral effects. Previously, in vivo studies have demonstrated the safety of the topical use of eugenol and carvacrol, components of clove and thyme oils, for the treatment of vaginal and oral candidosis in rats.33,34 Although the application of EOs may carry the risk of allergy and skin irritation, it has been reported that topical use of even 100% EO may be efficacious with no side effects for patient.35,36 Also, EO oral solutions appear to be safe and effective as alternative agents for patients with oropharyngeal candidiasis or in reducing plaque and gingivitis.37,38
This study indicates high homology of clinical and food-borne Candida isolates in relation to their susceptibility to anticandidal agents and hydrophobic properties. All Candida strains examined were susceptible to EOs tested (tea tree, thyme, peppermint, and clove), although peppermint oil demonstrated the lowest anticandidal activity. For majority of antibiotic-resistant strains, the high effectiveness of tea tree, thyme, and clove oils at their low MIC values (0.06–0.25% v/v) was noted. The susceptibility of yeasts to EOs could be related to their CSH. 39 High antifungal activity of EOs indicates their potential usefulness in the treatment or prevention of the development of Candida strains, both clinical and food borne, antibiotic sensitive, as well as antibiotic resistant.
Footnotes
Disclosure Statement
No competing financial interests exist.