
Abstract
Extended-Spectrum Beta-Lactamases (ESBL)-producing Escherichia coli have disseminated in both humans and animals worldwide. However, the ESBL epidemiology in these two reservoirs differs markedly, with CTX-M-15 frequently found in humans and CTX-M-1 preferentially found in animals. Our goal was to estimate the prevalence of fecal carriage of ESBL producers in cattle from 31 farms in Lebanon and to characterize the responsible enzymes. This prevalence was high (26/31, 84% of ESBL-positive farms), with a majority of isolates producing CTX-M-15 (27/40, 67.5%). Strikingly, this distribution is reminiscent of the human ESBL epidemiology, even though none of the bovine isolates belonged to the ST131 human clone. This is the first report of ESBL-producing E. coli in animals in Lebanon. Our data rather suggest the spread of CTX-M-15 plasmids in different E. coli backgrounds. Nonetheless, some CTX-M-15-producing E. coli clones found here have already been reported from animal, human, or environmental sources.
Introduction
I
Food-producing animals are generally considered a reservoir of ESBL-producing Enterobacteriaceae. In the bovine sector, ESBL genes have been reported in adult cattle in several countries from either fecal carriage or clinical mastitis.4,11,17–19 Nonetheless, these enzymes are even more prevalent in veal calves.5,20–23 This latter issue, at least partly, results from various practices at farms contributing to the selection of antimicrobial resistance, such as calves fed waste milk containing antibiotic residues (including broad-spectrum cephalosporins) or ESBL producers. 24
After Asia, the Middle East displays the second highest rate of ESBL-producing Enterobacteriaceae in human invasive infections and, in this world area, Lebanon presents the highest prevalence (46%). 25 In parallel, a national survey performed in 2011–2013 in hospitalized patients showed a prevalence of 32.29% in ESBL-producing Escherichia coli across the whole country. 26 Among human E. coli, CTX-M-15 is, by far, the most common enzyme responsible for the ESBL phenotype.25,27 Moreover, this enzyme can be found either at hospitals or in the community among healthy subjects. 28 Conversely, the situation in food-producing animals in Lebanon is still unknown. Thus, our goal was to determine the prevalence of fecal carriage of ESBL-producing E. coli in adult cattle sampled in the north and south parts of Lebanon, and to molecularly characterize the collected isolates.
Materials and Methods
Bacteria isolation and identification
Fecal samples from 174 healthy adult cattle were collected in 31 different farms between October 2012 and February 2013. A total of 18 farms (representing 114 samples) were from North Lebanon, and 13 farms (representing 60 samples) were from South Lebanon. Approximately 10 g of feces were precisely diluted 1/10 in phosphate buffer. Bacteria that were resistant to broad-spectrum cephalosporins were selected on Drigalski agar plates supplemented with ceftazidime (4 mg/L) and incubated at 37°C for 24 hr. A presumptive E. coli colony was arbitrarily selected from the selective plates. Isolates were identified using an MALDI-TOF MS (Microflex, Bruker).
Antimicrobial susceptibility testing and ESBL production
Antimicrobial susceptibility was tested by the disk diffusion method on Mueller–Hinton agar and interpreted according to the clinical breakpoints recommended by the Antibiogram Committee of the French Society of Microbiology (www.sfm-microbiologie.org). E. coli ATCC 25922 strain was used as the quality control. A total of 16 beta-lactam (amoxicillin, piperacillin, ticarcillin, amoxicillin-clavulanic acid, piperacillin-tazobactam, ticarcillin-clavulanic acid, cefalotine, cefuroxime, cefotaxime, ceftiofur, ceftazidime, cefoxitin, cefepime, cefquinome, aztreonam, and ertapenem) and 16 non-beta-lactam (tetracycline, kanamycin, tobramycin, gentamicin, amikacin, apramicin, netilmicin, streptomycin, florfenicol, chloramphenicol, colistin, sulfonamides, trimethoprim, nalidixic acid, ofloxacin, and enrofloxacin) antibiotics of veterinary and human interest was tested. ESBL production was determined by the double-disk synergy test. 29
Characterization of the resistance and virulence genes
The blaCTX-M genes were detected using a CTX-M group-specific multiplex PCR. 30 For the CTX-M-1 group, an additional PCR was performed using external primers (ISEcp1L1, 5′-CAGCTTTTATGACTCG and P2D, 5′-CAGCGCTTTTGCCGTCTAAG) and all amplicons were sequenced (Beckman Coulter, London, United Kingdom). On CTX-M-negative ESBL isolates, the blaTEM and blaSHV genes were screened by PCR and sequenced.31,32 The presence of the newly reported colistin resistance mcr-1 gene was systematically screened by PCR. 33 The presence of the stx1 and stx2 genes was detected by PCR to identify Shiga toxin-producing E. coli (STEC). 6
Phylogeny and genetic relatedness
The detection of the major E. coli phylogenetic groups (A, B1, B2, or D) was performed as recently described by Doumith et al. 34 PFGE was performed on all isolates using the restriction enzyme XbaI. 35 Electrophoresis was conducted in a CHEF DRIII system using 6 V cm−1 at 14°C for 17 hr, with pulse times ramping from 1 to 25 sec using an angle of 120°. PFGE results were interpreted according to international recommendations. 36 Multi-Locus Squence Typing (MLST) was performed on all isolates according to the protocol described on the E. coli MLST website (http://mlst.ucc.ie/mlst/dbs/Ecoli).
Results
A total of 40 ESBL-producing E. coli were identified (40/174, 22.9%). These resistant bacteria originated from 26 out of the 31 farms studied. Consequently, 84% of the farms were positive for the presence of at least one ESBL-producing isolate. Most isolates presented associated resistances to tetracyclines (82.5%), streptomycin (77.5%), sulfonamides (72.5%), and trimethoprim (75%). Resistances to chloramphenicol (30%), nalidixic acid (30%), enrofloxacin (17.5%), gentamicin (22.5%), and kanamycin (15%) were also detected. Intermediate resistance (inhibition zone of 16–17 mm) to colistin was observed in 15 isolates, but the newly described plasmidic mcr-1 gene was not detected and Minimum Inhibitory Concentrations were confirmed as susceptible by broth microdilution.
The 40 isolates belonged to phylogroup A (n = 19), B1 (n = 13), or D (n = 8) and none of them presented the stx1 or stx2 virulence genes. They were mainly nonclonal, with a large majority of isolates sharing PFGE profiles with less than 80% of homology (Fig. 1). Only a few strains were clonal (PFGE profiles with >90% of homology and an identical phylogroup): 36245/36251, 36266/36270, and 36246/36261 were from two different farms that were not located in the same village, whereas 36275/36279 originated from two different farms from the same village. Also, the two isolates 36244/36248 were from the same village, whereas the third isolate 36242 originated from a different one. The same situation was found for isolates 36273/36253, which originated from the same village in South Lebanon whereas 36271 was from a different one. Finally, 36256 was from North Lebanon and 36277 was from South Lebanon. MLST performed on all isolates revealed a high diversity of sequence types, as suggested by the diverging PFGE profiles (Fig. 1).
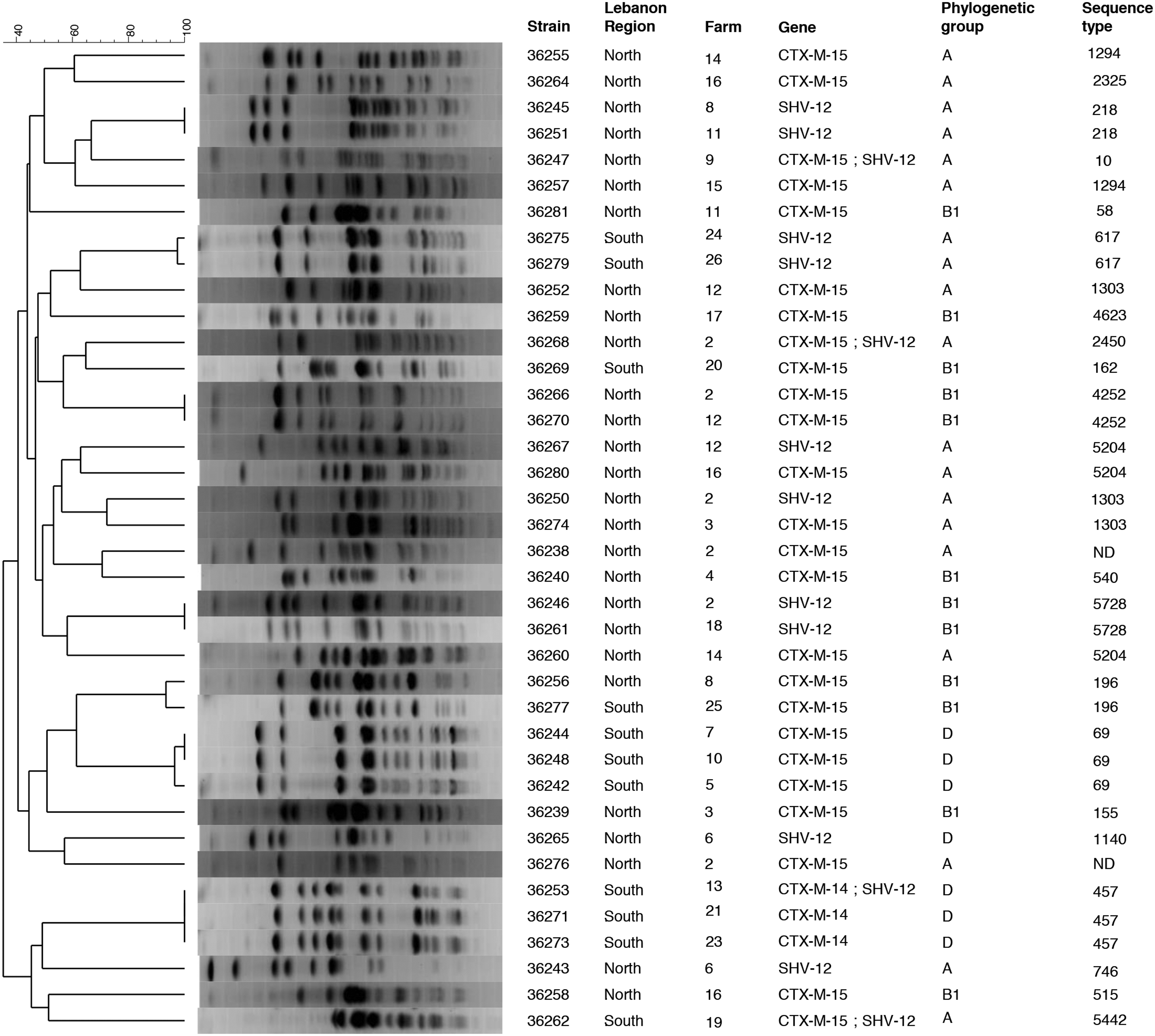
PFGE patterns and characteristics of 38 ESBL isolates. The two isolates 36237 (North Lebanon, CTX-M-15, phylogroup A) and 36722 (North Lebanon, CTX-M-15, phylogroup D), presenting smearing profiles despite several attempts, were not included in this figure.
The CTX-M-15 enzyme was identified in 27 isolates, of which three additionally produced an SHV-12 enzyme. The CTX-M-14 was detected in 3 isolates (1 also presenting an SHV-12), whereas the SHV-12 enzyme was identified alone in 10 isolates.
Discussion
In this study, we found that 84% of randomly chosen cattle farms in Lebanon were ESBL positive, with a total prevalence of 23% among ESBL healthy carriers. No STEC were identified. Direct comparisons with other studies are not always fully relevant because of methodological bias. Nevertheless, it is usually admitted that the prevalence of ESBL carriers is much lower in adult cattle compared with veal calves. 18 Since the prevalence found here in adults is higher than what has been reported in Europe or Japan in comparable populations,4,17–19 but similar to what has been reported in veal calves in Europe,5,22 our data suggest a massive colonization of the Lebanese bovine sector with ESBL producers. This would be also in line with the high rates of ESBL carriage in the human population in this country. Further studies are, thus, needed to estimate whether these high ESBL rates in humans and animals in Lebanon are inter-connected.
Most isolates presented multiple-associated resistances, highlighting that ESBL-producing E. coli can be selected by the use of other veterinary-licensed antibiotics than broad-spectrum cephalosporins. This is also coherent with the routine veterinary practices in Lebanon, since adult cattle are usually not treated by cephalosporins. Indeed, penicillin G in combination with streptomycin is routinely used for the local treatment of mastitis. Also, respiratory tract and foot infections are the most frequently treated with oxytetracyclines whereas gentamicin is one of the first-line treatments for gastro-intestinal enteritis. Of note, the highest resistance rates observed in this study were precisely to these latter antibiotics. On the contrary, the newly reported colistin resistance mcr-1 gene was not retrieved in this collection and this is in contrast with the high prevalence rates recently highlighted in cattle in France, although in young calves. 37
The enzymes responsible for the ESBL phenotype usually reported in cattle or calves are mostly CTX-M-1, followed by CTX-M-14.5,22,38 Here, most enzymes were CTX-M-15 (27/40, 67.5%), which are widely disseminated in the human community and in hospitals both in Lebanon and worldwide.25,27,28,38 However, the hypothesis of a direct transmission of CTX-M-15-producing E. coli clones from humans to cattle is surely too simplistic. Indeed, MLST revealed a high genetic diversity and none of the strains belonged to the ST131 clone that is mostly responsible for the spread of CTX-M-15 in humans. 39 Other widespread CTX-M-15-producing clones, such as ST405 or ST648, 12 were not identified either. On the contrary, some STs have been reported in both humans and animals (such as ST10, ST617, ST58, and ST69 as well as ST155 or ST1303), whereas others have no common history.40–42 This argues more in favor of a large reservoir of E. coli carrying and disseminating blaCTX-M-15 plasmids in diverse genetic backgrounds, as also recently proposed. 43 Such a plasmid-mediated spread may explain recent findings highlighting the presence of the blaCTX-M-15 gene in nonclinical contexts. 9 In humans also, a recent study showed fecal carriage with non-ST131 CTX-M-15 E. coli in certain human subgroups. 44 All these data may suggest possibly new epidemiological pictures of the blaCTX-M-15 gene in humans and animals.
The second most common enzyme identified in this study was SHV-12, which accounted for 35% (14/40) of all ESBL-producing E. coli and was detected either alone (n = 10) or in combination with a CTX-M-type enzyme (n = 4). SHV enzymes were largely distributed in humans until the 1990s,3,38 before being supplanted by CTX-M-type enzymes. They are now quite unfrequently isolated in E. coli around the world, but they have been reported even recently in Lebanon.27,45 Here also, the possible transmission from humans to animals remains to be documented.
In conclusion, this study is the first one reporting ESBL-producing Enterobacteriaceae in animals in Lebanon. In particular, we report a high prevalence of ESBL-producing E. coli in healthy cattle and ESBL-positive cattle farms in this country. Considering that adult cattle are usually poorly colonized by ESBL-producing Enterobacteriaceae compared with calves or other food-producing animals such as broilers, there is an urgent need to monitor the prevalence of these enzymes in other Lebanese animal species. Also, such a high prevalence in live animals makes us believe that food products derived from Lebanese livestock may be markedly contaminated with ESBL producers. In addition, we observed a surprising ESBL epidemiology in those animals, with CTX-M-15 being the most frequently identified enzyme. This partition is unusual in food-producing animals (even though CTX-M-15 were also sporadically reported in animals), and it is reminiscent of what is recurrently reported in humans. However, none of these CTX-M-15-producing E. coli were of ST131 and further work is now needed to clarify the sources and transmission pathways of the blaCTX-M-15 gene between humans, animals, and the environment.
Footnotes
Acknowledgments
The authors especially thank Taha Abdou, Maryam Yahya, Estelle Saras, Véronique Métayer, and Pierre Châtre for their helpful assistance.
Disclosure Statement
No competing financial interests exist.