
Abstract
Rapid antimicrobial susceptibility testing has the potential to improve patient outcomes and reduce healthcare-associated costs. In this study, a novel assay based on bacterial cell elongation after exposure to an antibiotic (ceftazidime) was evaluated for its ability to rapidly detect resistance in Gram-negative bacteria. The assay was used to detect resistance in a large collection of strains containing 320 clinical isolates of Acinetobacter baumannii, 171 clinical isolates of Klebsiella pneumoniae, and 212 clinical isolates of Pseudomonas aeruginosa, and the results were compared to those obtained using standard antimicrobial susceptibility testing methods. The assay identified ceftazidime-resistant strains with 100% sensitivity and 100% specificity for A. baumannii, 100% sensitivity and 97.2% specificity for K. pneumoniae, and with 82.3% sensitivity and 100% specificity for P. aeruginosa. Importantly, results were obtained in 1 hour 15 minutes from exponentially growing cultures. This study demonstrates that changes in cell length are highly correlated with phenotypic antibiotic susceptibility determined using standard susceptibility testing methods. This study therefore provides proof-of-concept that changes in cell morphology can be used as the basis for rapid detection of antibiotic resistance and provides the basis for the development of novel rapid diagnostics for the detection of antibiotic resistance.
Introduction
M
It has long been known that bacterial cells from multiple species adopt a filamentous shape in response to different types of environmental stress, including genome damage, 11 host immune effectors, 12 and nutrient limitation. 13 It has been hypothesized that this change in cellular morphology occurs due to cell-division arrest in the presence of continued volume expansion and that this phenotype is a component of a protective response that increases survival under stressful conditions. 14 Exposure to various antibiotic classes has also been shown to rapidly induce filamentation.13–15 In the present study, we use ceftazidime, a third-generation cephalosporin, to test the hypothesis that changes in bacterial morphology resulting from antibiotic exposure, which can be used to rapidly differentiate resistance from susceptible strains for various Gram-negative species. Our results indicate that monitoring cell morphology after antibiotic exposure can rapidly differentiate resistant and susceptible strains in agreement with standardized susceptibility testing methods and may therefore provide a basis for novel rapid susceptibility testing methods for bacterial infections.
Materials and Methods
Bacterial strains and minimum inhibitory concentration determination
Acinetobacter baumannii (n = 320), Pseudomonas aeruginosa (n = 212), and Klebsiella pneumoniae (n = 171) clinical isolates were obtained at University Hospital A Coruña between 2001 and 2013. The minimum inhibitory concentrations (MICs) were determined by automated microdilution (MicroScan WalkAway; Siemens) and confirmed by Etest (AB Biodisk; BioMerieux) according to the manufacturer's instructions, and susceptibility was interpreted using CLSI criteria. 15
Cell elongation assay
Bacteria cultured on solid media plates were used to inoculate a 2 ml Mueller-Hinton broth culture and incubated at 37°C for 90 minutes. Afterward, the bacteria were diluted to an OD600 of 0.1 in Mueller-Hinton broth and incubated at 37°C in 200 μl tubes with the indicated concentrations of ceftazidime for 60 minutes in a final volume of 30 μl. To assess cell length, a previously described protocol was used.16,17 Briefly, after incubation with the antibiotic, the bacteria were diluted to a concentration of 1 × 107 microorganisms/ml in Mueller-Hinton broth, included in a thin microgel on a slide, incubated in increasing concentrations of ethanol, dried, stained with the fluorochrome SYBR Gold, and examined under fluorescence microscopy. This process produces perfectly sharp images that accurately and confidently establish cell length. For all assays, digital image analysis (Apogee KX32 ME camera and Visilog 5.1 software; Noesis) was used to assess at least 200 cells/assay. Elongation was defined as an increase in length of at least 1.5-fold compared to untreated cells of the same strain. This methodology was used since it is not biased by intrinsic differences in cell length between strains.
Determining resistance using cell elongation
The relationship between cell elongation and CLSI breakpoint concentrations of ceftazidime was empirically established after processing multiple strains from each bacterial species comprising a wide range of MICs, including susceptible, intermediate, and resistant strains. Numerous strains with MICs close to the CLSI breakpoints for susceptibility and resistance were used. Antibiotic concentrations that increase cell length in the susceptible, but not in the intermediate and resistant strains and in the susceptible and intermediate strains only were determined. The concentrations were optimized such that a strain categorized as susceptible following CLSI criteria must demonstrate cellular elongation after incubation with the lowest concentration established as the breakpoint for susceptibility when using the parameter of elongation. In addition, the cells from an intermediate strain (according to CLSI criteria) must not exhibit elongation after incubation with the lowest concentration established as the breakpoint of susceptibility, but will appear elongated with the concentration established as the breakpoint for resistance using the parameter of cell elongation. Finally, a strain categorized as resistant following the CLSI criterion should not appear elongated with the concentrations of antibiotics used. Susceptibility testing using the cell elongation technique and by traditional methods was carried out by independent researchers, and results were compared at the end of the study.
Results
Cell elongation after antibiotic exposure was easily and reproducibly detected as can be seen in Fig. 1, in which changes in cell length for three A. baumannii strains incubated with increasing doses of ceftazidime (0, 2, 4, 6, 8 mg/L) are presented (200 cells were analyzed for each concentration tested). They correspond to a susceptible (MIC: 2.5 mg/L), an intermediate (MIC: 24 mg/L), and a resistant strain (MIC: 48 mg/L). In the susceptible strain, increase in cell length with respect to the control culture without antibiotic was evident from lowest concentration used, 2 mg/L. The intermediate strain did not show cell enlargement after incubation with 2 mg/L, but it was significant after 4 mg/L and progressively increased as the dose of cephalosporin increased. There was no significant cell elongation in the resistant strain at any concentration assayed. Similar results were obtained with K. pneumoniae and P. aeruginosa, representative of susceptible, intermediate, and resistant strains being presented in Fig. 1. In these cases, the ceftazidime doses used were much lower than in A. baumannii, as explained below. Figure 2 shows microscopic images of the three A. baumannii strains at the indicated concentrations of ceftazidime. Importantly, upon starting with isolated bacterial colonies on solid media, total time to the final result was ∼3.5–4 hours, including 30 minutes of hands-on time.

Cell length modification with increasing concentrations of ceftazidime in representative susceptible (S; above), intermediate (I; middle), and resistant (R; below) strains from Acinetobacter baumannii (S: MIC 2.5 mg/L; I: MIC 24 mg/L; R: MIC 48 mg/L), Klebsiella pneumoniae (S: MIC 0.047 mg/L; I: MIC 8 mg/L; R: MIC 256 mg/L), and Pseudomonas aeruginosa (S: MIC 8 mg/L; I: MIC 16 mg/L; R: MIC 64 mg/L). Ceftazidime concentrations are indicated in the horizontal axis (mg/L), whereas cell length is shown on the vertical axis (μm). The data are presented as box and whisker plots. The horizontal line in the box represents the median, the lower line of the box is the first quartile, the upper line of the box is the third quartile, and the whiskers (the end of the vertical lines) are maximum and minimum data values. The dots outside the box correspond to outlier values. Cells appear progressively larger in the susceptible strains, but without significant modification at any dose in the resistant strain. MIC, minimum inhibitory concentration.
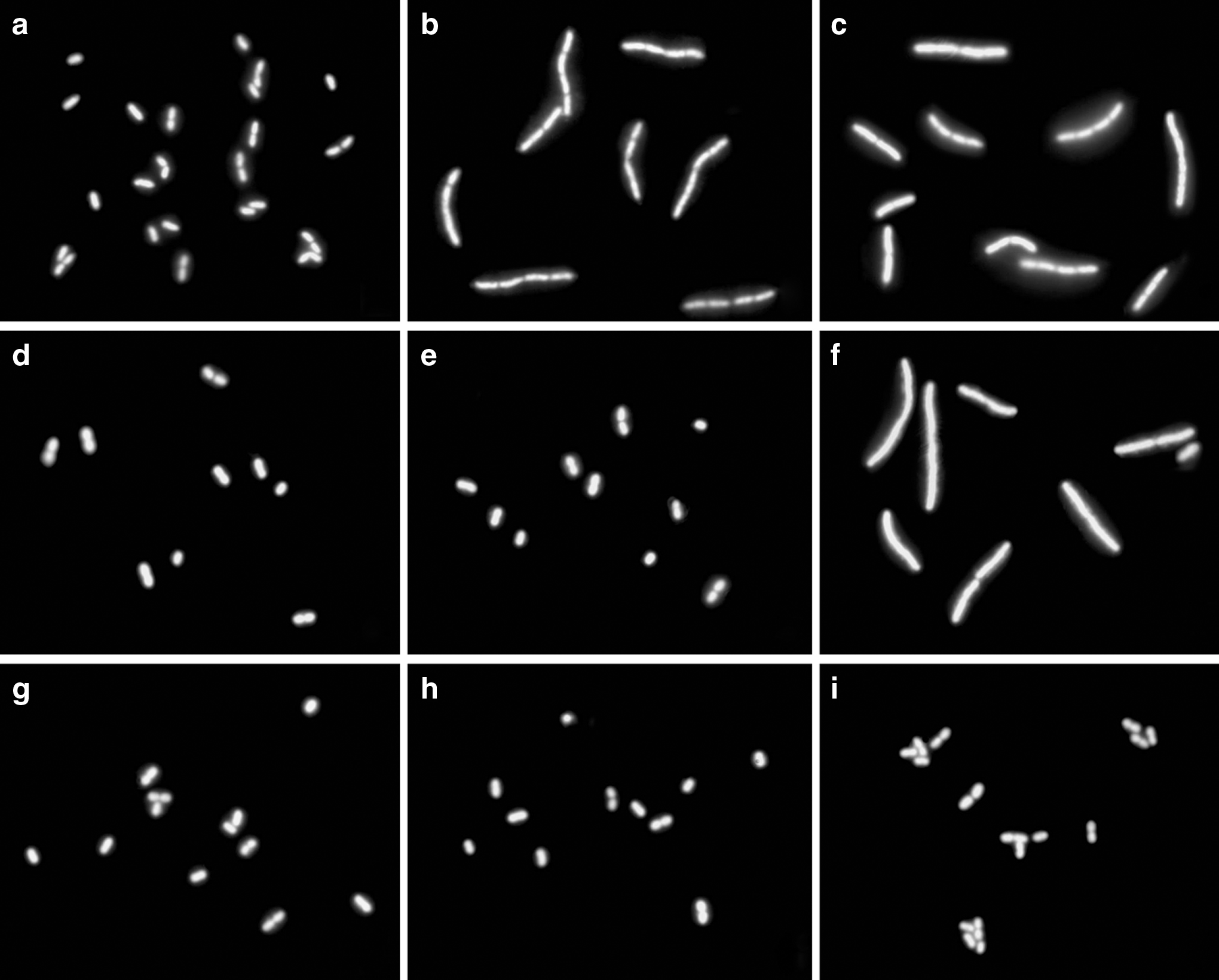
Strains of A. baumannii, incubated 1 hour with ceftazidime, enclosed in a microgel on slides, dehydrated, stained with SYBR Gold, and observed under fluorescence microscopy. First column left
For A. baumannii, the CLSI breakpoint MICs for ceftazidime are ≤8 and ≥32 mg/L for susceptibility and resistance, respectively. Using the cell elongation assay, we found that concentrations of ≤2 and >8 mg/L corresponded to susceptibility and resistance, respectively. That is, susceptible strains appear enlarged after incubating with 2 and 8 μg/ml and intermediate strains only after 8 mg/L, whereas resistant strains never appear affected in cell length with both concentrations (Fig. 2). To validate the assay, 320 A. baumannii clinical isolates were processed. Following standard MIC and CLSI guidelines, 51 strains were considered susceptible, 35 intermediate, and 234 resistant to ceftazidime. Using the criterion of elongation with their respective, empirically-determined breakpoint concentrations, all the strains were correctly identified with 100% sensitivity and specificity (Table 1).
For K. pneumoniae, CLSI breakpoints for susceptibility and resistance for ceftazidime are ≤4 and ≥16 mg/L, respectively, whereas the breakpoints in the cell elongation assay concentrations of ≤0.5 and >1.25 mg/L were shown to correlate with susceptibility and resistance. One hundred seventy-one clinical isolates of K. pneumoniae were studied, with 107 susceptible, 19 intermediate, and 45 resistant to ceftazidime according to CLSI standards. The results obtained using the cell elongation assay were highly concordant with these results. The only exceptions were three susceptible strains categorized as intermediate (2) and resistant (1) using the cell elongation-based assay and two intermediate strains that were classified as resistant. These results indicate that the cell elongation-based assay was more conservative, with a sensitivity of detection of nonsusceptible strains (intermediate and resistant) of 100% and a specificity of 97.2%. The false positive rate was 2.8% and no false negatives with resistance were detected. The positive predictive value was 95.5% and the negative predictive value was 100%.
For P. aeruginosa, the CLSI breakpoint concentrations for susceptibility and resistance to ceftazidime are ≤8 and ≥32 mg/L, respectively. When bacterial length was used, breakpoint concentrations for cell enlargement were determined to be much lower, ≤0.5 and >1 mg/L. When analyzing 212 isolates of P. aeruginosa, 178 were identified as susceptible, 18 intermediate, and 16 resistant to ceftazidime using CLSI guidelines. All susceptible strains were correctly identified with the elongation-based assay. However, six nonsusceptible strains demonstrated elongation after incubating with 0.5 μg/ml ceftazidime. Therefore, the cell elongation-based assay showed 82.3% sensitivity and 100% specificity. Whereas the false positive rate was 0% and the positive predictive value 100%, the false negative rate and the negative predictive value were 17.6% and 96.7%, respectively.
Discussion
This study demonstrates that changes in cell elongation can be used to rapidly detect resistance to ceftazidime in three Gram-negative species that often produce multidrug-resistant infections. To our knowledge, this is the first study that has used cell elongation to characterize antibiotic susceptibility and correlate the results with standardized testing methods using a large collection of clinical isolates. It is interesting to note that for all three bacterial species tested, the concentration of antibiotic required to induce cellular elongation was below the breakpoint for resistance defined by CLSI broth microdilution methods. These data would suggest that physiological changes occur in bacterial cells at concentrations that are subinhibitory according to standard microbiological testing. In agreement with this idea, previous work has reported that bacterial filamentation occurs at subinhibitory concentrations of antibiotics.16–18 Importantly, we demonstrate in this study that cell elongation at these subinhibitory concentrations correlates with susceptibility testing results using standard methods that define resistance breakpoints using higher antibiotic concentrations. Our results indicate that the elongation assay was somewhat less sensitive for detecting resistance in P. aeruginosa isolates; however, the mechanism underlying this difference compared to K. pneumoniae and A. baumannii is not clear.
Other methods for rapidly detecting antibiotic resistance in bacterial isolates such as nucleic acid-based tests (e.g., PCR and microarrays), mass spectroscopy-based approaches, and whole genome sequencing are currently being used clinically or are being developed. 2 All of these approaches have potential for rapid susceptibility testing; however, in most cases, they require more hands-on time than traditional approaches. Importantly, all of these methods require previous knowledge of the resistance mechanism for susceptibility testing; for example, the gene(s) that confers resistance must be known for PCR and microarray-based assay, and the specific metabolite of interest or the proteins producing resistance must be characterized in mass spectroscopy-based approaches. Thus, one major limitation of these methods is that they may not demonstrate high sensitivity in detecting uncharacterized mechanisms of resistance. Although it has not been empirically tested in this study, the cell elongation assay may not depend upon the mechanism of resistance to ceftazidime, and may, therefore, be able to detect uncharacterized resistance mechanisms, thus minimizing false negative results. An additional advantage is that the measurement of bacterial cell elongation can also be adapted to increase automation using automated microscopy and appropriate software for the detection of changes in cell length. It may therefore be possible to reduce the hands-on time required in the present study.
A critical issue regarding the method described in this study is its applicability to antibiotics other than ceftazidime. The bactericidal activity of ceftazidime is due to its specific binding to different penicillin-binding proteins (PBPs), inhibiting the final transpeptidation step required for peptidoglycan synthesis. The main targets are PBP1b, 1c, PBP2, and especially PBP3.19,20 It has low affinity for PBP4, resulting in very weak induction of the chromosomally encoded AmpC β-lactamase. 21 Numerous resistance mechanisms have been described for ceftazidime, including the production of drug-destroying enzymes such as β-lactamases, altered drug targets such as PBPs, decreased bacterial permeability due to porin alterations, and increased drug efflux mechanisms. 22 Although cellular elongation has been reported to occur after exposure to multiple antibiotics,16–18 it has not been shown that elongation correlates with phenotypic susceptibility determined by standard methods for these antibiotics. We point out, however, that during preliminary studies, we tested the ability of the cell elongation technique to detect resistance to ciprofloxacin. Our results indicated that cell elongation could not be used to reliably detect resistance to ciprofloxacin (data not shown). This may be due to the fact that ciprofloxacin does not target cell wall biosynthesis. With regard to additional antibiotics, further empirical testing is required to determine the applicability of this technique.
In summary, we demonstrate that changes in cell length are highly correlated with phenotypic susceptibility to ceftazidime determined using standard susceptibility testing methods. This study therefore provides proof-of-concept that changes in cell morphology can be used as the basis for rapid detection of antibiotic resistance.
Funding
This work has been supported by MagicBullet, the Fondo de Investigaciones Sanitarias PI14/01346, and by REIPI: Spanish Network for Research in Infectious Diseases (Instituto de Salud Carlos III, RD06/0008/0025). M.J.M. is supported by the Subprograma Miguel Servet from the Ministerio de Economía y Competitividad of Spain (CP11/00314). MagicBullet is a project funded by the European Union-Directorate General for Research and Innovation through the Seventh Framework Program for Research and Development (grant agreement number 278232).
Footnotes
Disclosure Statement
J.L.F. and J.G. are advisers of Halotech DNA SL. All other authors declare no conflicts.