
Abstract
The whole genome sequencing of extensively drug-resistant Shewanella xiamenensis T17 isolated from hospital effluents in Algeria revealed the presence of a novel 268.4 kb plasmid designated pSx1, which carries several antibiotic-resistance genes in the novel Tn1696 derivative (Tn6297), in addition to the chromosomal blaOXA-48-like gene (blaOXA-416). The presence of the plasmid was confirmed by nuclease S1-PFGE analysis and transformation by electroporation into Escherichia coli DH10B. Tn6297 contains an In27 class 1 integron harboring the dfrA12-orfF-aadA2 array, msr(E) and mph(E) associated with IS26; a new efflux pump multidrug resistance composite transposon delimited by two ISEc29s; Tn-tet harboring tetR and tetA(C); a class 1 integron with the qacG gene cassette; qnrVC6 and dfrA23 associated with ISCR1; and a complex class 1 integron In4-like containing aacC1, aadA1, blaVEB-16, catA2, sul1Δ, cmlA9, tetR, tetA(G), aac(6′)-II, and blaPSE-1. Its mer operon carries merB, but lacks merC, in contrast to Tn1696 and Tn21. This study represents the first characterization of a multidrug-resistant transposon and multidrug resistance plasmid in Shewanella and is the first report of blaOXA-416 in Algeria, providing evidence that Shewanella spp. could be an important reservoir and vehicle for drug resistance genes.
Introduction
M
Studies on the origin of ARGs have suggested that antibiotic resistance found in clinical settings was intimately associated with mechanisms found in the environment, 3 which may contribute to the emergence and spread of these genes.
Shewanella xiamenensis is a recently identified bacterial species from coastal sea sediment in China 4 and is rarely isolated from human infections. 5 Several naturally occurring β-lactamases (OXA-48-like carbapenemases) and quinolone resistance determinants (Qnr-type) have been detected in the Shewanella genus, indicating that it is an important reservoir of these genes.6–9
The blaOXA-48-like genes encode carbapenem-hydrolyzing enzymes. Five variants of these genes have been reported in S. xiamenensis strains: blaOXA-181, 10 blaOXA-48b, blaOXA-199, 11 blaOXA-204, 12 and blaOXA-416. 6 In this study, an extensively resistant strain of S. xiamenensis T17 was isolated in Algeria from hospital effluents and characterized using whole genome sequencing. This led to the isolation and characterization of a large plasmid, pSx1, which harbored a new MDR transposon derived from Tn1696 and designated Tn6297. The blaOXA-416 gene was also identified on the chromosome of this strain.
Materials and Methods
Bacterial strain identification, antimicrobial susceptibility testing, and extended-spectrum β-lactamase detection
Shewanella T17 was isolated on MacConkey agar supplemented with 2 μg/ml cefotaxime from a wastewater sample collected on March 2012 from the Nedir Mohamed Hospital's effluent collector in Tizi Ouzou, Algeria. Identification at the species level was based on the 16S rRNA and gyrB complete nucleotide sequences as described by Huang et al. 4
Antimicrobial susceptibility testing was performed using a disk diffusion assay on Mueller-Hinton agar plates, broth microdilution, and E-test according to the Clinical and Laboratory Standards Institutes (CLSI) guidelines. Extended-spectrum β-lactamase (ESBL) production was tested by the double-disk (DD) test using both cefotaxime and ceftazidime disks with or without clavulanate. 13
Cloning of blaOXA-48-like and transformation in Escherichia coli
The genes blaOXA-416 and blaOXA-48 (Ay236073 GenBank accession number) were synthesized and cloned in pUC57-Kan-plasmid at GenScript. The two recombinant plasmids and the nonrecombinant (control) were transformed into E. coli DH10B by electroporation using a MicroPulser according to the manufacturer's instructions (Bio-Rad). Transformants were selected on LB plates containing 45 μg/ml of kanamycin according to GenScript recommendations and subjected to antimicrobial susceptibility testing.
Shotgun sequencing, sequence assembly, and bioinformatics analysis
Whole genome sequencing was performed using PacBio (Pacific Biosciences) at the Yale Center for Genome Analysis, Yale School of Medicine, and Yale University (New Haven, CT) using 9 μg of purified genomic DNA. Genomic DNA was purified with the MagAttract DNA Mini M48 Kit using the BioRobot M48 (Qiagen) according to the manufacturer's instructions. PacBio genome sequencing was performed using the PacBio SMRTbell Template Prep Kit v 1.0 (Pacific Biosciences). PacBio long reads were assembled using the Hierarchical Genome-Assembly Process (HGAP) (v3). Consensus polishing was performed with the Quiver algorithm to generate 2 high-quality unitigs. 14 The unitigs were annotated using Prokka (v 1.11) (http://vicbioinformatics.com). The annotation presented in this study was confirmed manually using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and Artemis genome visualization tools (www.sanger.ac.uk/science/tools/artemis). Insertion sequences were identified using ISfinder (www-is.biotoul.fr).
The new transposon number was assigned using the Tn Number Registry webpage (www.ucl.ac.uk/eastman/research/departments/microbial-diseases/tn).
PlasmidFinder (http://cge.cbs.dtu.dk/services/PlasmidFinder) was used to search for known replicon types. traI was compared to plasmid sequences from other γ-Proteobacteria and used to design primers for MOB typing. 15
For strain identification, the S. xiamenensis T17 genome was compared with the genomes of 20 other Shewanella species using the Genome-to-Genome Distance Calculator (GGDC; http://ggdc.dsmz.de/). 16
Detection and sizing of the large plasmid by the S1 nuclease/PFGE method
To detect and confirm the size of the pSx1 plasmid, DNA-containing agarose plugs were incubated for 60 min at 37°C with 10 U Aspergillus oryzae S1 nuclease (Thermo Fisher Scientific). 17 The digested DNA plugs were electrophoresed at 200 V for 19 hr. Salmonella enterica serotype Braenderup strain H9812 was used as the size marker on the gel. 18
Plasmid isolation, electroporation, and conjugation
Plasmid DNA was extracted according to a modified QIAGEN Plasmid Mini Kit protocol. Solutions P1, P2, and P3 were used for lysis, neutralization, and precipitation, respectively. After centrifugation (15,000 × g at 4°C), the supernatant was poured into a new tube, and the nucleic acid was precipitated by adding isopropanol and centrifuged as described above. The pellet was washed with 70% ethanol (v/v), and the plasmid was resuspended in 30 μl of TE buffer. Two microliters of this extract was used for transformation into E. coli DH10B (Thermo Fisher Scientific) by electroporation according to the manufacturer's instructions using Bio-Rad MicroPulser. The transformants were selected on LB plates containing 32 μg/ml of ceftazidime and were subjected to antibiotic susceptibility testing.
For conjugation, the broth mating technique was performed using S. xiamenensis T17 as the donor with E. coli J53 AzR as the recipient. Donor and recipient cells were mixed at a ratio of 1:4, 1:1, and 4:1 donor-to-recipient. Luria-Bertani medium plates containing sodium azide (100 μg/ml), chloramphenicol (15 μg/ml), and ceftazidime (4 μg/ml) were used to select the transconjugants.
PCR detection of the dfrA12, dfrA23, and qnrVC6 genes in E. coli DH10B (pSx1)
PCR reactions were performed in a 25 μl volume according to the manufacturer's guidelines using AmpliTaq DNA polymerase (Thermo Fisher Scientific). The primers and PCR conditions for the dfrA12 and dfrA23 PCRs were described previously.19,20 Primers to amplify qnrVC6 were designed in this study: VC-f (5′-AGTGAACTTCTCACATCAGG-3′) and VC-r (5′-TCAGGAACAATGATTACCCC-3′), and the same PCR conditions as used for dfrA23 with an annealing temperature of 50°C were used.
Accession numbers
The whole genome of S. xiamenensis, blaOXA-416 with its genetic environment and pSx1 sequences were deposited in GenBank under accession numbers LDOA01000000, KU198597, and CP013115.
Results
Bacterial strain identification and susceptibility testing
Shewanella isolate T17 exhibited 99.5% 16S rRNA nucleotide sequence identity with the S. xiamenensis type strain S4. 4 A comparison of the gyrB sequence with those available in GenBank showed similarities between 95.6% and 98.3% (accession nos. FJ589040, KC765141, KR780672, and KP264118) with 95.6% similarity with S. xiamenensis type strain S4. 4 In addition, a comparison between the T17 genome with the genomes of 20 other Shewanella species by GGDC showed a high similarity (80.90% ± 3.65) with S. xiamenensis BC01, confirming the identification of isolate T17 as S. xiamenensis.
Susceptibility testing showed S. xiamenensis T17 to be resistant to at least 16 antibiotics from 11 different classes, including penicillin, ampicillin, piperacillin, ticarcillin, cefotaxime, ceftazidime, cefepime, ertapenem, aztreonam, gentamicin, streptomycin, ciprofloxacin, trimethoprim/sulfamethoxazole, tetracycline, azithromycin, and chloramphenicol. This isolate was susceptible to imipenem, meropenem, amoxicillin–clavulanate, ticarcillin–clavulanate, amikacin, and colistin. The MICs (minimum inhibitory concentration) of the antibiotics are shown in Table 1. The DD test was positive for ESBL production.
E. coli DH10B (pUC57-OXA-416), (pUC57-OXA-48), (pUC57) and (pSx1) transformants.
AMK, amikacin; AMP, ampicillin; AMX, amoxicillin; ATM, aztreonam; AZM, azithromycin; CAZ, ceftazidime; CIP, ciprofloxacin; CST, colistin; CTX, cefotaxime; ETP, Ertapenem; FEP, cefepime; GEN, gentamicin; IPM, imipenem; MEM, meropenem; PEN, penicillin; PIP, piperacillin; SXT, trimethoprim–sulfamethoxazole; TET, tetracycline; TZP, piperacillin–tazobactam.
ND, MICs not determined.
Genomic assembly and cloning and expression of blaOXA-416
PacBio sequencing was used to produce reads larger than 10 kb, and Genome assembly yielded two unitigs, 0 (5,085,271 bp) and 1 (282,919 bp) covering the chromosome and a large plasmid, with G + C contents of 46.27% and 47.7%, respectively.
blaOXA-416 was located on unitig 0, S. xiamenensis T17 chromosome. The genetic context of this gene was similar to that described for blaOXA-48-like genes in other S. xiamenensis strains11,12,21 and shared 91.5–100% nucleotide identity (Fig. 1).
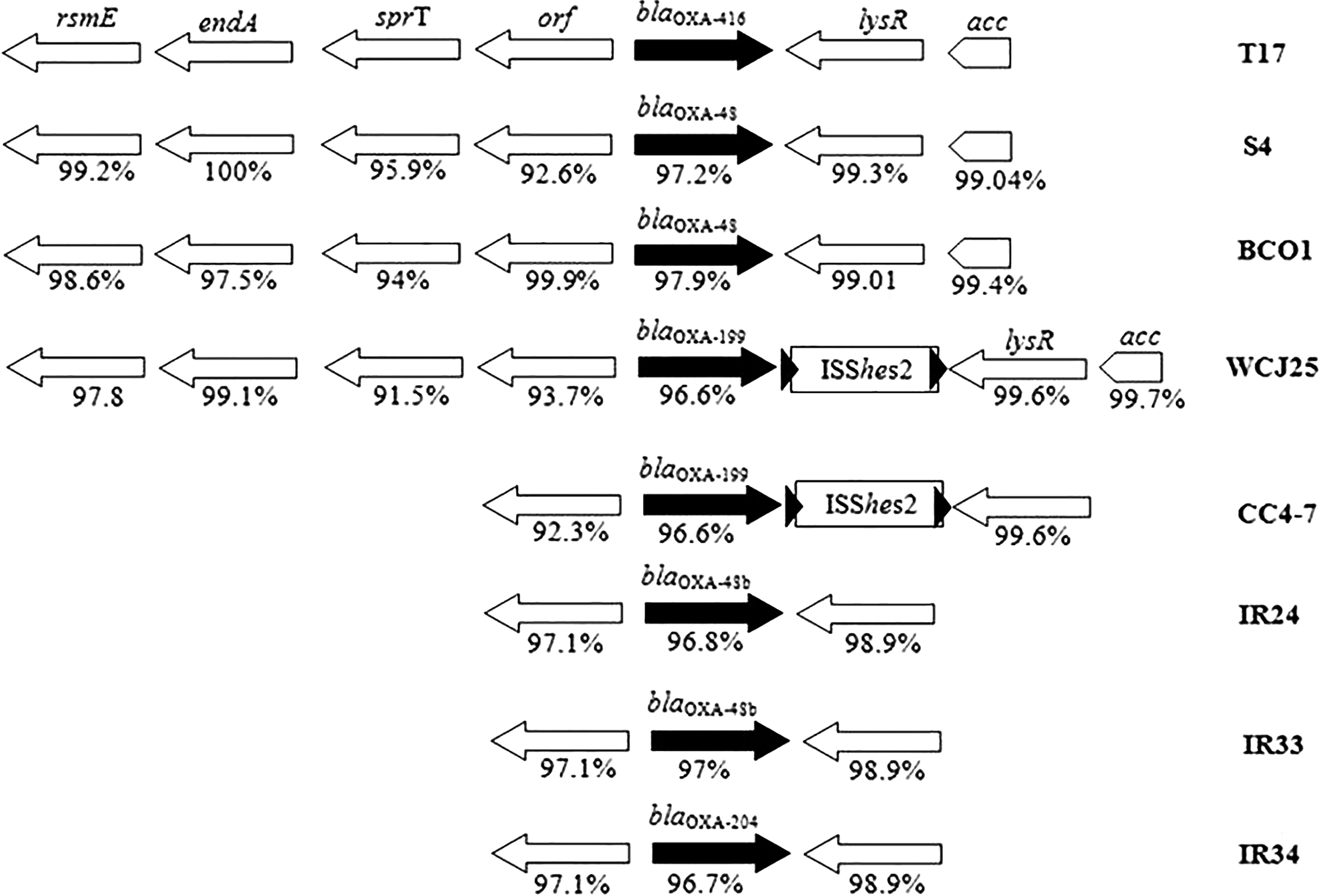
Comparison of the genes surrounding blaOXA-416 identified in Shewanella xiamenensis T17 with those of S. xiamenensis strains S4, 11 BC01 (GenBank accession number JGVI01000025), WCJ25, 11 CC4-7, 21 IR24, IR33, and IR34. 12 The blaOXA-48-like genes are shown as black arrows, and the genes upstream and downstream are shown as white arrows. The acc sequences of S4 and WCJ25 that were used for the comparison were partials. The black triangles indicate the direct repeats (CCT) flanking ISShes2.
The E. coli DH10B (pUC57-Kan-OXA-416) and (pUC57-Kan-OXA-48) transformants showed MICs of 2 μg/ml for imipenem and ertapenem, 1 μg/ml for meropenem, and >128 and 32/4 μg/ml for piperacillin and piperacillin/tazobactam, respectively (Table 1). The MIC of piperacillin was decreased by tazobactam in S. xiamenensis T17 and E. coli DH10B (pUC57-OXA-416) and (pUC57-Kan-OXA-48), suggesting that OXA-48 and OXA-416 may be partially inhibited by tazobactam.
Experimental detection of a novel large plasmid
The 14,538 bp at both ends of unitig 1 showed 99% identity, indicating that it was likely a circular plasmid. In addition, the sequence coverage of unitig 1 was significantly higher than that of unitig 0 (226.36X vs. 96.92X). The read-length coverage of the plasmid was 2.35 times higher than that of the chromosome, suggesting the presence of approximately two copies of the plasmid per chromosome. The presence of this large plasmid was further confirmed by S1 nuclease digestion followed by PFGE, which showed a single band with an estimated size of 270 kb. Plasmid electroporation into E. coli showed the growth of ceftazidime-resistant strains, confirming the existence of this plasmid. This new plasmid was designated pSx1. The attempted conjugation of pSx1 into E. coli was unsuccessful.
The sequence analysis of pSx1 revealed the presence of repA encoding an initiator protein and parA and parB encoding partition proteins and transfer genes, including traC_F, traV, trbI, traK, traE, traL, traD, traF-like, traH, traG-like, traR, traG, virD2-like, traI, traF, traN, traU, and traW. No known replicon type was identified using PlasmidFinder. This plasmid was not classified into any MOB family based on the sequence analysis of relaxase.
Based on the AT content and direct/inverted repeats, the predicted replication and transfer origins oriV and oriT were located at 193,670–194,613 bp and 248,457–249,184 bp, respectively.
Identification and description of a new transposon, Tn6297
XDR-associated genes are mainly clustered in a 78,178 bp segment on pSx1 located between nucleotide positions 81,645 and 159,821 bp. The G + C content of this segment is 53.96% compared with 45.1% for the rest of the plasmid. The plasmid harbors 84 ORFs and is delimited by two 38 bp inverted repeats (IR tnp and IR mer ) from Tn1696 and flanked by a pair of short 5 bp direct repeats (ATGAT), suggesting a duplication event during transposition (Fig. 2). Designated Tn6297, this transposon includes three class 1 integrons and transposons associated with drug resistance genes.

Schematic representation of the 78-kb multidrug resistance Tn6297 in S. xiamenensis T17. Black arrows with white dots correspond to the transposases, resolvases, and tni module genes. Gray arrows represent the integrase genes, and dashed arrows indicate the resistance genes. Black arrows represent the 3′-CS conserved segment of class 1 integrons, and white arrows indicate the other genes, including ORFs with unknown function. The IS are shown as gray boxes. The ovals represent the attC gene cassettes recombination sites. IS, insertion sequences.
The In27 class 1 integron carries 5′-CS, which contains the intI1, dfrA12-orfF-aadA2 gene cassette array conferring resistance to trimethoprim and aminoglycosides and the conserved segment 3′-CS (qacEΔ1, sul1, and orf5). It includes tniBΔ1 and tniA (part of the tni module), a hybrid of Tn402 and related transposons, and is flanked by inverted repeats from Tn402 (IRi and IRt-like).
The second class 1 integron harbors a qacG gene cassette conferring resistance to quaternary ammonium compounds. This integron lacks IRi and IRt, and its integrase was disrupted by the insertion of Tn-tet.
The third class 1 integron is 26,640 bp with an In4-like backbone. It is delimited by a pair of inverted repeats, IRi and IRt, contains three 3′-CS (3′-CS1, 3′-CS2, and 3′-CS3), ISCR1, and ISCR3. It also includes the aacC1-orfP-orfQ-aadA1 array, which has been reported in MDR Acinetobacter baumannii from several countries. 22 3′-CS1 is followed by a tetracycline resistance gene derived from transposon Tn1721 (tetA(A)Δ5′) and a truncated blaVEB-16 gene cassette, which confers high-level resistance to broad-spectrum cephalosporins and aztreonam. blaVEB-16 is flanked by 135 bp repeat elements (Re2 and Re1), which could play a role in its movement by homologous recombination.23,24 Downstream of 3′-CS2, this integron contains catA2 (encoding chloramphenicol acetyltransferase), sul1Δ (which confers resistance to sulfamethoxazole), cmlA9 (encoding chloramphenicol efflux protein), tetR and tetA(G) (which confer resistance to tetracycline), lysR (transcriptional regulator), groEL/intI1 (chaperonin/integrase), aac(6′)-II gene cassette (which confers resistance to aminoglycosides), and blaPSE-1 gene cassette (conferring resistance to β-lactams (ampicillin)). The blaPSE-1 gene cassette is followed by 3′-CS3 (qacEΔ1, sul1, orf5, and orf6). At the right hand, a region similar to that found in the standard In4-type integron is present. This includes the last 123 bp from the IRt end of Tn402 in inverse orientation, followed by IS600, and the last 152 bp from the IRt end of the Tn402 tni module. Mercury resistance genes (merE, merD, merB, merA, merP, merT, and merR) were also detected. The segment covering sul1Δ, cmlA9, tetR, tetA(G), lysR, ISCR3, and groEL/intI1 has 99% identity to SGI2 of S. enterica subsp. enterica serovar Emek. 25
A composite transposon harboring a set of genes encoding multidrug efflux pumps (acrA, acrB, and oep) and a transcriptional regulator (acrR) was identified. This transposon is delimited by two ISEc29 and contains tnpMut encoding a mutator transposase. The second ISEc29 of the efflux system composite transposon lacks the IRR; this could be due to the insertion of Tn-tet delimited by two IS26. These insertion sequences have been shown to cause adjacent deletions excluding the evidence of insertion provided by DR.26,27 No similar organization was found in other described microorganisms. This region exhibits a G + C content of 45.16%, close to the G + C content of S. xiamenensis (46.27%), and different from the G + C content of Tn6297 (53.96%). This suggests that this region is native and unique to S. xiamenensis. The second composite transposon Tn-tet is delimited by two IS26 and contains tetA(C) and its regulator tetR. Additional genes, including res (resolvase) and xre (antitoxin), were detected.
The msr(E) encoding a macrolide efflux pump protein and mph(E) encoding macrolide 2′-phosphotransferase protein were located downstream of In27 and associated with IS26.
A truncated qnrVC6 gene cassette and dfrA23 conferring resistance to quinolone and trimethoprim, respectively, were associated with ISCR1. The qnrVC6 gene cassette was followed by orfc372-2 (unknown function) and gcn5 (encoding related N-acetyltransferase). The qnrVC6 was previously described in a Vibrio parahaemolyticus plasmid. 28 This is the first report of the qnrVC6 gene in the Shewanella genus. The fragment covering the last 8 bp at the 3′ end of gcn5, dfrA23, the second copy of Tn1696, and up to the 114 bp upstream of intI1 from the In4-like complex class 1 integron shares 99% identity with a homologous region from the virulence plasmid t-ST4 of an S. enterica serotype Typhimurium isolate. 29
Because the antimicrobial susceptibilities of ciprofloxacin and trimethoprim/sulfamethoxazole for the E. coli DH10B (pSx1) transformants were not consistent with the antimicrobial susceptibilities obtained from S. xiamenensis T17 (Table 1), dfrA12, dfrA23, and qnrVC6 were amplified. The results showed that these genes were absent from E. coli DH10B (pSx1) transformants and present in the positive control S. xiamenensis T17. This could be explained by possible genetic rearrangements in E. coli, resulting in the deletion of these genes.
Discussion
The genus Shewanella has been regarded as a reservoir and vehicle of ARGs due to the several antibiotic resistance determinants identified in this genus, including OXA-54 oxacillinase identified in Shewanella oneidensis, which is considered to be the progenitor of OXA-48. 8 The blaOXA-48-like genes were the most reported in S. xiamenensis.10–12 In addition, aacA3, qnrA1, dfrA1, aadA2az, msr(E), and mph(E) were also detected in MDR S. xiamenensis CC4-7. These genes were associated with the mobile genetic elements (class 1 integron, IS26 and ISCR1). 21
This extensively drug-resistant S. xiamenensis T17 isolated from hospital waste water in Algeria is the first complete genome of S. xiamenensis. A draft genome of S. xiamenensis, strain BC01, is available in the GenBank database (GenBank accession number JGVI00000000). The genome size was 4,677,169 bp with a GC content of 46.21%, whereas the complete chromosome size of S. xiamenensis T17 was 5,085,271 bp. A genomic analysis detected a chromosomally encoded OXA-416 and a new MDR Tn1696 derivative named Tn6297 located on a novel large plasmid (pSx1).
Recently, a new variant of blaOXA-48, blaOXA-416, was identified in S. xiamenensis DDP1 from a fecal sample from a 1-year-old child in Italy. 6 The resistance profile of T17 was significantly different from that of DDP1. The genetic organization of blaOXA-416 in isolate T17 was similar to the organization of other blaOXA-48-like genes found previously in S. xiamenensis harboring blaOXA-48, blaOXA-48b, and blaOXA-204. However, insertion sequences have only been found in the immediate environment of blaOXA-199 of S. xiamenensis WCJ25 and CC4-7.11,21 The detection of S. xiamenensis producing blaOXA-416 in the intestine of a 1-year-old patient in Italy (after infection with Klebsiella pneumoniae) and in the hospital effluents in Algeria (this study) emphasizes the potential role of Shewanella species as reservoirs of carbapenemase genes, especially of blaOXA48-like.
The most interesting finding of this study is the identification of the large plasmid pSx1 harboring the majority of the resistance genes responsible for the XDR phenotype. These genes were clustered on the new transposon Tn6297, a large Tn1696 derivative, and associated with several mobile genetic elements. To date, only a few plasmids have been described in other Shewanella strains and do not encode antimicrobial resistance genes.
The expression of resistance genes associated with pSx1 in E. coli was demonstrated after electroporation in E. coli. However, the mobilization of the plasmid by conjugation should be confirmed under optimized conditions because transfer genes and predicted transfer origin have been identified.
The complex transposon Tn6297 has likely undergone several genetic rearrangements through transposition and recombination. Its mercury resistance module includes the merB gene, but lacks merC, making it different from that encoded by Tn1696 and Tn21. It does share 99% identity with mercury resistance modules in the pRMH760 and pDU1358 plasmids of K. pneumoniae and Serratia marcescens, respectively.30,31
The In27 and complex class 1 integrons were preceded by tnp1696. These types of integrons are known to target the res sites of the Tn21 family. 32 The dfrA12-orfF-aadA2 gene cassette array associated with In27 is widely distributed in Gram-negative bacteria, such as S. enterica and E. coli.33,34 The absence of DRs flanking the IRi and IRt of these class 1 integrons suggests recombination events. 35
Tn-tet is 99% identical to that previously identified on the plasmids of uncultured bacterium pB3 36 and the MDRGI of Stenotrophomonas maltophilia GZP-Sm1. 37
The combination ISCR1-3′-CS inserted into class 1 integrons could move different structures located upstream. 2 In Tn6297, ISCR1 associated with qnrVC6 could play a role in the dissemination of this gene to other bacteria.
The phenotype ACSSuT (resistance to ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline) is a public health concern, as it was described and is currently monitored in the important foodborne pathogen Salmonella spp.25,38 In the present study, this phenotype is mainly encoded on a complex class 1 integron in Tn6297 of S. xiamenensis T17; therefore, we cannot exclude the role of Shewanella as a reservoir of this resistance phenotype and/or in its dissemination. In addition, sequence analysis revealed that the first 5-kb fragment covering intI1, the aacC1-orfP-orfQ-aadA1 array and the 3′-CS1 of In4-like complex class 1 integron, shares extensive nucleotide sequence identity with the corresponding sequences in several A. baumannii strains.22,39,40 This suggests a common ancestor or an exchange between S. xiamenensis and one or more A. baumannii strains.
A similar blaVEB-16 region is also present in several Proteus mirabilis strains, for example, the SGI1-V genomic island of P. mirabilis strain VB1248. 41 The presence of homologous blaVEB-16 regions in P. mirabilis and S. xiamenensis T17 indicates that they may have been acquired from a common ancestor.
This study provides new knowledge on the genetic content of S. xiamenensis strain T17 with an XDR phenotype, including the identification of the new transposon Tn6297. To our knowledge, this is the first report of MDR plasmid in Shewanella and the first identification of dfrA12, dfrA23, aacC1, aadA1, aac(6′)-II, tetA, qacG, qnrVC6, blaVEB-16, blaPSE-1, catA2, and cmlA9 in this species.
The coexistence of this plasmid with a large MDR transposon in hospital sewage isolate denotes its evolution and its potent ability to acquire and disseminate different resistance genes. The detection of such extensively resistant Shewanella supports the idea that the environment could play a critical role in the dissemination of multidrug resistance.
Footnotes
Acknowledgments
The authors thank the bacteriology team and Sylvie Clermont at Laboratoire de santé publique du Québec for technical assistance. Thanks are extended to Florence Doualla-Bell and Ashley Kearney for the revision of the article.
Disclosure Statement
No competing financial interests exist.