
Abstract
This first nationwide study was conducted to analyze the prevalence of plasmid-mediated quinolone resistance (PMQR) genes in phenotypically unselected (consecutive) clinical enterobacteria. We studied 1,058 isolates that had been consecutively collected in 66 hospitals of the WHONET-Argentina Resistance Surveillance Network. Overall, 26% of isolates were nonsusceptible to at least one of the three quinolones tested (nalidixic acid, ciprofloxacin, and levofloxacin). The overall prevalence of PMQR genes was 8.1% (4.6% for aac(6′)-Ib-cr; 3.9% for qnr genes; and 0.4% for oqxA and oqxB, which were not previously reported in enterobacteria other than Klebsiella spp. from Argentina). The PMQR prevalence was highly variable among the enterobacterial species or when the different genes were considered. The prevalent PMQR genes were located in class 1 integrons [qnrB2, qnrB10, and aac(6′)-Ib-cr]; in the ColE1-type plasmid pPAB19-1 or Tn2012-like transposons (qnrB19); and in Tn6238 or bracketed by IS26 and blaOXA-1 [aac(6′)-Ib-cr]. The mutations associated with quinolone resistance that were located in the quinolone resistance-determining region (QRDR mutations) of gyrA, parC, and gyrB were also investigated. The occurrence of QRDR mutations was significantly associated with the presence of PMQR genes: At least one QRDR mutation was present in 82% of the PMQR-harboring isolates but in only 23% of those without PMQR genes (p < 0.0001, Fisher's Test). To the best of our knowledge, this is the first report on the prevalence of PMQR genes in consecutive clinical enterobacteria where all the genes currently known have been screened.
Introduction
T
Besides the chromosomal-driven mechanism described above, three kinds of plasmid-mediated quinolone resistance (PMQR) mechanisms have been described. These constitute the second most relevant group of quinolone resistance mechanisms and are based on: (i) Qnr proteins, which protect the quinolone targets; (ii) the enzyme Aac(6′)-Ib-cr, which acetylates aminoglycosides and also ciprofloxacin and norfloxacin; and (iii) efflux pumps, which either specifically (QepA) or unspecifically (OqxAB) decrease the intracellular concentration of quinolones. 7 Five classes of qnr genes are currently known, namely qnrA, qnrB, qnrS, qnrC, and qnrD, with the first three including several alleles (www.lahey.org/qnrStudies). PMQR genes and QRDR mutations are often found together in enterobacteria with high-level quinolone resistance, highlighting the interplay and evolution of both mechanisms in resistant clinical isolates. 7
In the previous decade, many studies on PMQR prevalence were conducted in clinical enterobacteria but the vast majority of them focused on isolates with specific resistance phenotypes, such as resistance/decreased susceptibility to quinolones; resistance to aminoglycosides or expanded-spectrum cephalosporins (ESC); and production of extended-spectrum β-lactamases, which are commonly associated with PMQR genes. These selection criteria greatly overestimate the prevalence of these genes in unselected populations of clinical enterobacteria. 7 Only few PMQR surveys were based on phenotypically unselected isolates, that is, those that were consecutively isolated from clinical specimens in the microbiology laboratory of the hospital.8–22 Most of these studies analyzed a single8–15 or a few16–20 of the enterobacterial species found during the study period, whereas only two of them included all the species found.21,22 In regard to Argentina, only two surveys were reported and they addressed PMQR prevalence in clinical enterobacteria with resistance/decreased susceptibility to quinolones 23 or ESC resistance. 24
Here, we report a large nationwide study based on a point prevalence collection of 1058 unselected clinical enterobacteria that we conducted to estimate the prevalence of all the PMQR genes currently known.
Materials and Methods
Bacterial isolates and antimicrobial susceptibility testing
We used a collection of 1,058 unselected (consecutive), nonduplicated enterobacterial isolates. These strains were specifically collected for this prevalence study during the same 5 consecutive days in 2007, in the 66 hospitals of the WHONET-Argentina Resistance Surveillance Network, which were located in Buenos Aires City, and the 23 provinces of Argentina (Supplementary Fig. S1). The bacterial collection comprised 673 Escherichia coli; 133 Klebsiella pneumoniae; 10 Klebsiella oxytoca; 66 Shigella spp.; 58 Proteus mirabilis; 7 Proteus vulgaris; 38 Enterobacter cloacae; 14 Enterobacter aerogenes; 18 Serratia marcescens; 1 Serratia liquefaciens; 8 Citrobacter freundii; 7 Citrobacter koseri; 12 Morganella morganii; 9 Salmonella spp., and 4 Providencia stuartii (see epidemiological data in Supplementary Table S1). The 81 isolates of the tribe Proteeae (Proteus, Morganella, and Providencia) were analyzed in a preliminary paper, 6 but we included in this work some results reported therein to facilitate the analysis of the complete bacterial collection.
Antimicrobial susceptibility tests were performed by disk diffusion (DD) and agar dilution according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI). 25 DD was used to determine the susceptibility profile to three quinolones [nalidixic acid (NAL), ciprofloxacin, and levofloxacin] and 19 antibiotics of other families (ampicillin, amoxicillin-clavulanic acid, piperacillin-tazobactam, cephalothin, cefoxitin, cefotaxime, ceftazidime, ertapenem, imipenem, meropenem, amikacin, gentamicin, kanamycin, tobramycin, chloramphenicol, colistin, tigecycline, trimethoprim, and trimethoprim-sulfamethoxazole). Agar dilution was used to determine the MICs of NAL, ciprofloxacin, and levofloxacin.
PCR and DNA sequencing
All the primers used for PCR and DNA sequencing are listed in Supplementary Table S2. DNA sequencing was performed by using BigDye terminators with an ABI 3500 Genetic Analyzer (Applied Biosystems/Perkin Elmer, Foster City, CA). Sequences were edited and analyzed with BioEdit, v7.2.5 (www.mbio.ncsu.edu/bioedit/bioedit.html), 26 ClustalX2, v2.1 (www.clustal.org/clustal2), 27 and the BLAST algorithm (www.ncbi.nlm.nih.gov/BLAST).
Detection of PMQR genes
The presence of qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxA, and oqxB (only in oqxA-positive isolates) was tested by PCR as described. 23 The different qnrB or qnrS alleles were identified by PCR and DNA sequencing of complete genes. The presence of aac(6′)-Ib-cr and/or aac(6′)-Ib was only determined in the subset of isolates that were nonsusceptible to kanamycin (DD inhibition zones ≤17 mm) by using separate allele-specific PCRs (ASPCRs) with two primer sets directed against each variant. 23
Analysis of the QRDRs of gyrA, parC, and gyrB
Only the mutations located in QRDRs that were previously associated with quinolone resistance3,28 were considered (named onward as QRDR mutations). The QRDR of gyrA was analyzed by PCR and DNA sequencing as described. 23 The QRDR of parC was additionally analyzed 23 in those isolates where at least one QRDR mutation was detected in gyrA. The QRDR of gyrB from some isolates was also analyzed as described. 6 To determine the presence of QRDR mutations, the obtained gyrA, parC, or gyrB sequences were compared with reference sequences (Supplementary Table S2).
Genetic environments of PMQR genes
The genetic environments of qnrB2 and qnrB10 were analyzed by PCR cartography as described. 23 The qnrB19-harboring plasmid pPAB19-1 was identified by PCR with intragenic divergent primers and restriction fragment length polymorphisms with ApoI and RsaI as described. 23 The ISEcp1C-based genetic support of qnrB19 29 was identified by PCR and DNA sequencing (Supplementary Table S2).
The genetic environments of aac(6′)-Ib-cr were analyzed by PCR reactions that were designed to amplify the 5′- and 3′-genetic surroundings that corresponded to the most common locations of this gene, namely class 1 integrons, Tn6238, and those bracketed by IS26 and blaOXA-1.30,31 Briefly, the 5′-reactions were done by using alternative forward primers that were directed against intI1, the tnpA of Tn6238, or the tnpA of IS26, and the reverse primer used in the ASPCR for aac(6′)-Ib-cr; whereas the 3′-reactions were performed with the forward primer used in the ASPCR for aac(6′)-Ib-cr and alternative reverse primers directed against the 3′-conserved segment of class 1 integrons, blaOXA-9 or blaOXA-1. All the obtained amplicons were sequenced.
Results and Discussion
Analysis of quinolone susceptibility
Overall, 26% (279/1058) of the isolates were nonsusceptible (resistant plus intermediate CLSI categories 25 ) to at least one of the three quinolones tested. The distributions of quinolone susceptibility according to species are summarized in Figure 1. Excluding the particular cases of Shigella spp. and P. stuartii, for which almost all of the isolates were fully susceptible or fully resistant to quinolones, respectively, the levels of nonsusceptibility ranged from 11% (Salmonella spp.) to 42% (M. morganii), for NAL; from 7.1% (E. aerogenes) to 37% (E. cloacae), for ciprofloxacin; and from 0% (K. oxytoca) to 36% (P. mirabilis), for levofloxacin (Fig. 1).

Quinolone susceptibility in the enterobacterial collection. For each indicated species (Pst, Providencia stuartii; Mmo, Morganella morganii; Ecl, Enterobacter cloacae; Pmi, Proteus mirabilis; Kpn, Klebsiella pneumoniae; Sma, Serratia marcescens; Eco, Escherichia coli; Cfr, Citrobacter freundii; Eae, Enterobacter aerogenes; Kox, Klebsiella oxytoca; Pvu, Proteus vulgaris; Sal, Salmonella spp.; and SHI, Shigella spp.), the percentages of nonsusceptibility (resistant plus intermediate categories) and decreased ciprofloxacin susceptibility (MIC of 0.12–1 μg/ml, Salmonella is excluded) are shown (highest and lowest values are indicated above the bars). Data for the Proteeae species are from our preliminary work. 6 All the Citrobacter koseri and the unique isolate of Serratia liquefaciens were susceptible to all quinolones.
We also examined the level of decreased ciprofloxacin susceptibility (DCS, defined as a ciprofloxacin MIC of 0.12–1 μg/ml) in all the species except Salmonella spp., for which DCS is currently included in the CLSI categories of nonsusceptibility. 25 DCS was observed in 64 of 1,049 (6.1%) isolates (Salmonella was excluded), of which 48 (75%) were nonsusceptible to NAL. Interestingly, S. marcescens showed the highest DCS level (44%) observed in our collection, being two-times higher than the corresponding percentage of ciprofloxacin nonsusceptibility (Fig. 1). This might be due to the presence of the chromosomal qnr homolog (smaqnr) described in S. marcescens. 32
In the E. coli subset of the collection, 499 isolates were fully susceptible to NAL (MIC ≤4 μg/ml) and also to ciprofloxacin and levofloxacin; 2 isolates displayed susceptibility to NAL (MIC of 8–16 μg/ml) with DCS (NALS-DCS); 7 isolates showed intermediate or low resistance to NAL [(NALI-LR), MIC of 32–64 μg/ml, DD inhibition zones of 9–17 mm], and 165 isolates were highly resistant to NAL (MIC ≥128 μg/ml, DD inhibition zones ≤8 mm). To reduce the number of isolates to be analyzed, a sample was taken from the E. coli subset as follows: 10% of the isolates fully susceptible to quinolones (50 out of 499) and 25% of those highly resistant to NAL (41 out of 165) were randomly selected, whereas all the 2 NALS-DCS and the 7 NALI-LR isolates were included (Supplementary Fig. S2). The obtained sample of 100 isolates was used in the subsequent analyses.
Occurrence and prevalence of PMQR genes
The PMQR genes were found in 29 out of 66 hospitals, located in Buenos Aires City and 15 of the 23 provinces of Argentina. The qnr genes were detected in 30 isolates: qnrB2, 9 isolates; qnrB10 and qnrB19, 7 isolates each; qnrB9, 2 isolates; qnrB28 (first report in Argentina) and qnrB6, 1 isolate each; and qnrS1, 3 isolates. In addition, we had previously found qnrD1 in 1 P. mirabilis and 1 P. vulgaris. 6
Thirty isolates harbored aac(6′)-Ib-cr. In 22 of them, it was the unique PMQR gene (14 Klebsiella spp., 6 E. coli, and 2 E. cloacae). The remaining eight isolates were the only ones that harbored more than one PMQR gene and harbored aac(6′)-Ib-cr combined with qnrB10 (2 K. pneumoniae, 1 E. cloacae, and 1 C. freundii), qnrB2 (3 K. pneumoniae), or qnrS1 (1 K. pneumoniae). qnrB2 and qnrB10 were significantly associated with aac(6′)-Ib-cr (p = 0.014 and p < 0.001, Fisher's Test, respectively), whereas qnrS1 was not (p = 0.175, Fisher's Test).
The qepA gene was not found, and oqxA and oqxB were detected in 1 E. coli (isolate Q4160 from a urine specimen) and in 92% of the Klebsiella sp. isolates (93% of K. pneumoniae, 90% of K. oxytoca). However, it was reported that in Klebsiella spp. these genes were mostly located on the chromosome and their presence did not correlate with the ciprofloxacin MIC, probably due to different expression levels of the oqxAB operon.23,33–35 Consistently, considering the Klebsiella sp. isolates of this work that did not harbor other PMQR genes nor QRDR mutations in gyrA or parC, the ciprofloxacin MICs of the isolates with and without oqxA and oqxB were almost identical (MIC50 of 0.03 μg/ml, range 0.008–0.06 μg/ml and MIC50 of 0.03 μg/ml, range 0.015–0.06 μg/ml, respectively). Therefore, the subsequent analyses of PMQR genes were carried out regardless of the presence of oqxA and oqxB in Klebsiella spp.
E. coli Q4160 showed intermediate resistance to chloramphenicol and trimethoprim, as expected from the expression of OqxAB. 36 The sequences of the amplified fragments of oqxA and oqxB from Q4160 were identical to those of pOLA52, the representative oqxA-/oqxB-harboring plasmid of E. coli. 7 This is the first report of oqxA and oqxB in enterobacteria other than Klebsiella spp. from Argentina. The mobilization of oqxA and oqxB was evaluated by biparental conjugation of Q4160 and E. coli J53 azide-resistant, as described 23 (150 μg/ml of sodium azide and 8 μg/ml of chloramphenicol were used for transconjugant selection). Although several attempts were made, no transconjugants were obtained.
Interestingly, 45 of the 55 PMQR-harboring isolates showed high resistance to NAL with variable ciprofloxacin susceptibility, from DCS to high resistance (Table 1). In particular, all the 10 PMQR-harboring E. coli were included in the sample subgroup that was highly resistant to NAL (41 isolates), which was the 25% of the corresponding one (165 isolates) in the complete E. coli subset (see section Analysis of Quinolone Susceptibility). Therefore, to calculate prevalence, the number of E. coli isolates in the enterobacterial collection that contained a given PMQR gene was estimated by multiplying by four the number of isolates with this gene in the sample set. This resulted in 85 out of 1,058 isolates with PMQR genes giving an overall prevalence of 8.1%, with disaggregated values of 4.6% for aac(6′)-Ib-cr, 3.9% for the qnr genes (mostly qnrB alleles, 3.1%), and 0.4% for oqxA/oqxB. However, the PMQR prevalence was highly variable among the species of the collection and ranged from 3.1% in Proteus spp. 6 to 22% in Klebsiella spp., with an outlier value of 63% for C. freundii (Fig. 2A). The overall PMQR prevalence found here was similar to that observed in phenotypically unselected clinical enterobacteria from Spain (6.4%) 22 and was well below those found in the two earlier surveys from Argentina, namely 57% 23 and 60%, 24 because both studies were based on selected isolates with resistance/decreased susceptibility to quinolones and ESC resistance, respectively.
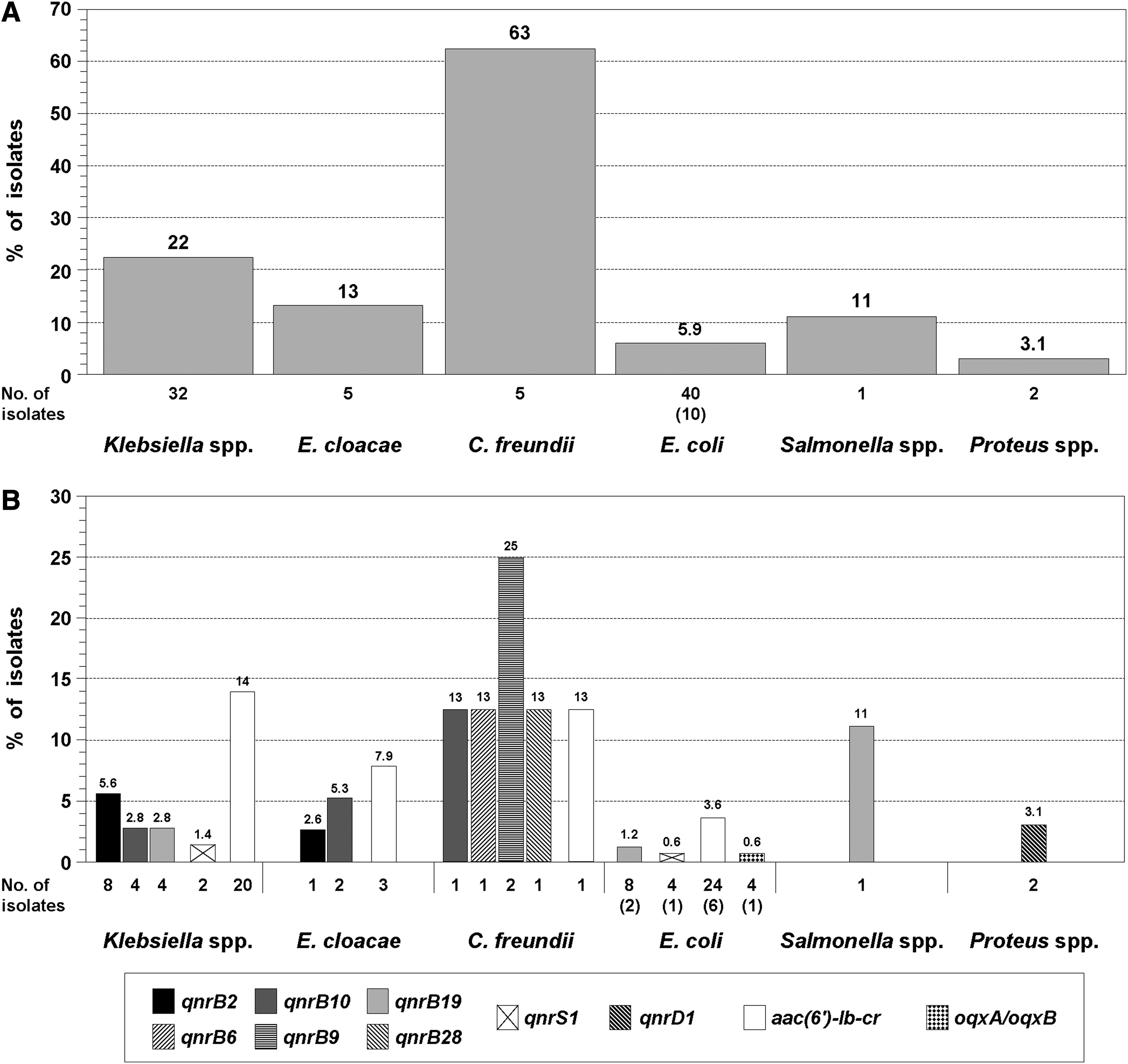
Prevalence of PMQR genes. In the collection of 1,058 isolates, the number of E. coli that harbored a given PMQR gene was estimated by multiplying by four the number of isolates with this gene found in the sample set of 100 E. coli, which is indicated between brackets. For simplicity, K. pneumoniae and K. oxytoca are grouped together, as well as P. mirabilis and P. vulgaris (data are from our preliminary work
6
).
The ranges of MIC and DD for each group are shown. PMQR genes were not detected in Enterobacter aerogenes, Citrobacter koseri, Serratia spp., and Shigella spp., nor in Morganella morganii and Providencia stuartii. 6
The different PMQR genes and combinations of them found in each group are indicated.
Number of isolates with or without at least one QRDR mutation in gyrA.
Proteus mirabilis and Proteus vulgaris, data from our preliminary work. 6
DD, disk diffusion inhibition zone; MIC, minimum inhibitory concentration; PMQR, plasmid-mediated quinolone resistance; QRDR, quinolone resistance-determining region.
Besides the interspecies variability in the overall PMQR prevalence, there were differences among the species when the different PMQR genes were considered. First, Salmonella spp. and Proteus spp. only harbored a unique gene (qnrB19 and qnrD1, respectively), whereas three to five different genes were found in the other species. Moreover, qnrD1 was only found in Proteus spp. (Fig. 2B). These findings agree well with a previous work with another enterobacterial collection from Argentina, where it was observed that Salmonella spp. contained almost exclusively qnrB19. 23 Similarly, although qnrD1 has been occasionally reported in other enterobacteria, it was especially found in some species of the tribe Proteeae, such as P. mirabilis, P. vulgaris, and Providencia rettgeri, and could have originated there. 7 Second, qnrB6, qnrB9, and qnrB28 were only found in C. freundii (Fig. 2B), which is consistent with the notion that this species is likely the source of qnrB alleles. 7 This concept could also account for the high PMQR prevalence found here for C. freundii, since all the PMQR-harboring isolates of this species contained qnrB alleles (Fig. 2B). In other surveys based on phenotypically unselected enterobacteria, it was also found that the qnr genes in C. freundii were almost exclusively qnrB alleles and that this species had the highest qnr prevalence (25%–71%).16,19,21,22
Coexistence of QRDR mutations and PMQR genes
In this study, 45 of the 55 PMQR-containing isolates showed high resistance to NAL, suggesting the presence of QRDR mutations in the topoisomerase II-encoding genes.3,23 The presence of QRDR mutations in gyrA and parC was investigated in all the PMQR-containing isolates and in the isolates that were nonsusceptible to NAL (DD inhibition zones ≤18 mm) of those without PMQR genes. QRDR mutations were only found in two groups (Table 2). First, all the isolates that were highly resistant to NAL (DD inhibition zones ≤8 mm, MIC ≥128 μg/ml) harbored QRDR mutations in the codon 83 of gyrA. The 30% of this group had a second QRDR mutation in gyrA (codon 87), and 83% showed at least one QRDR mutation in parC. Second, only the isolates of E. coli and Shigella sp. with NAL inhibition zones of 9–14 mm (MIC of 32–64 μg/ml) harbored the single QRDR mutations in gyrA, S83A, or D87G/Y, which were reported to confer low-level NAL resistance.37–39 These specific QRDR genotypes were observed in 6 of the 7 NALI-LR E. coli (S83A, D87Y, and D87G, 1, 2, and 3 isolates, respectively) and in the unique NAL-resistant Shigella sp. (D87G) (Table 2), giving a prevalence of 0.9% in E. coli and of 1.5% in Shigella spp.. The limited distribution of these QRDR genotypes can be related, at least in part, with the low-level quinolone resistance that they confer 38 and is in agreement with previous reports. Indeed, single substitutions in codon 87 were the unique QRDR mutations found in NAL-resistant S. flexneri from Argentina, 23 whereas S83A was also found occasionally among clinical enterobacteria from other regions.28,40 It is important to highlight that the seven isolates with these uncommon QRDR genotypes were susceptible to ciprofloxacin and levofloxacin according to the MIC and DD breakpoints 25 (Table 1). Moreover, one of the D87Y-harboring E. coli and the D87G-containing Shigella sp. resulted in being susceptible when they were tested with the pefloxacin 5-μg disk 41 that was proposed for the screening of ciprofloxacin nonsusceptibility in Salmonella. 25
Only the isolates with at least one QRDR mutation were considered (n = 144).
Kpn, Klebsiella pneumoniae; Kox, Klebsiella oxytoca; Ecl, E. cloacae; Eae, E. aerogenes; Cfr, C. freundii; Sma, Serratia marcescens; Eco, E. coli; SHI, Shigella spp.; Pmi, P. mirabilis; Mmo, M. morganii; Pst, P. stuartii. The unique isolate of P. vulgaris that contained QRDR mutations (gyrA, S83I and parC, S80I) 6 is not included in the table for simplicity. The percentages of the individual amino acid variants found in each indicated codon of gyrA or parC were calculated over the total number of isolates of each enterobacterial species in Group 1 or Group 2 (values for the prevalent amino acid variants are indicated in bold; “—”: variant not found). The gray background indicates that there were no isolates with a particular phenotype (SHI in Group 1 and all the species, except Kpn, in Group 2) or the isolates with a particular phenotype did not harbor QRDR mutations (Kpn in Group 2).
Data from our preliminary work. 6
T83I in Kox, Eae, and Cfr.
This group includes six of the seven E. coli isolates that showed intermediate or low resistance to NAL (NALI-LR). The remaining NALI-LR isolate showed a DD of 17 mm for NAL and did not harbor QRDR mutations. In this group, the ciprofloxacin MICs were 0.015–0.12 μg/ml and the levofloxacin MICs were 0.015–0.5 μg/ml (DD of 28–36 mm and 25–35 mm, respectively).
NAL, nalidixic acid; WT, wild type.
In Salmonella Typhi, the single gyrB substitution S464F has been associated with an unusual quinolone resistance phenotype.4,5 This phenotype was very similar to that of the unique quinolone-resistant Salmonella of our collection (Table 1), which showed wild-type gyrA and parC QRDRs, but the analysis of the gyrB QRDR of this isolate did not reveal any QRDR mutation. In addition, in the preliminary analysis of the Proteeae isolates of our collection, we observed that single QRDR mutations in both gyrA and parC were not sufficient to reach high-level ciprofloxacin resistance and that an additional substitution in codon 464 of gyrB (S464F/Y) was necessary. 6 To know whether these mutations could also be involved in the high-level ciprofloxacin resistance observed in both K. pneumoniae and E. cloacae, the gyrB QRDRs of four isolates of each species were analyzed but no substitution was found in codon 464.
The occurrence of QRDR mutations was significantly associated with the presence of PMQR genes: At least one QRDR mutation was present in 82% (45/55) of the PMQR-harboring isolates but in only 23% (99/430) of those without PMQR genes (p < 0.0001, Fisher's Test) (Tables 1 and 3). Such a high proportion of QRDR mutations in PMQR-harboring isolates was also observed in other surveys based on phenotypically unselected enterobacteria,10,11,13,14 but the occurrence of QRDR mutations in isolates without PMQR genes was not analyzed in any of them, hampering the statistical association provided here.
For simplicity, species of the same genus are grouped together.
Percentages were calculated over the total number of isolates with or without PMQR genes into each species.
E. coli sample of 100 isolates described in the section Analysis of Quinolone Susceptibility.
This subgroup includes the six isolates with intermediate or low resistance to nalidixic acid (NALI-LR) that harbored S83A or D87G/Y in gyrA as the unique QRDR mutation found.
Data from our preliminary work. 6
The high level of coexistence of QRDR mutations and PMQR genes could be due, at least in part, to the possible presence of several clonally related PMQR-harboring isolates. However, in the analysis of susceptibility to 19 antibiotics other than quinolones, we observed that the isolates of the same species that harbored the same PMQR gene, or combination of PMQR genes, had different susceptibility profiles (Table 4). Moreover, these isolates were mostly collected in different hospitals (Table 4). These facts suggest extensive genetic heterogeneity among the PMQR-harboring isolates.
The isolates are grouped according to: (i) whether they had or had not at least one QRDR mutation in gyrA; (ii) the enterobacterial species; and (iii) the PMQR genes found in each isolate.
The species that only contained 1 PMQR-harboring isolate were excluded (P. mirabilis, P. vulgaris, and Salmonella spp.).
For simplicity, the different qnr genes found (the qnrB alleles and qnrS1) were not shown. The combinations of qnr genes with aac(6′)-Ib-cr are indicated by a plus sign.
Number of different profiles of susceptibility to 19 antibiotics other than quinolones observed in each group.
The two E. coli isolates collected in the same hospital contained different qnr genes (qnrB19 and qnrS1).
It is well known that the PMQR genes facilitate the selection of higher levels of quinolone resistance. 7 In vitro studies on E. coli showed that these genes promoted the selection of mutations that conferred higher levels of quinolone resistance, which were located preferentially in several targets that were different from the type II topoisomerase-encoding genes.42–45 However, a different outcome was observed in vivo. First, several observational studies showed that QRDR mutations in gyrA were commonly found in clinical PMQR-harboring isolates of E. coli and other enterobacteria.10,11,13,14,30,46 Second, the intrapatient selection of QRDR mutations in gyrA and parC over a qnr-positive background was observed during fluoroquinolone treatment of infections caused by E. coli 47 or Salmonella Typhimurium. 48 Moreover, two longitudinal surveys provided additional support for a role of the PMQR genes in promoting the emergence of QRDR mutations in gyrA and parC in clinical settings.17,20 Although our results do not allow us to figure out which of these mechanisms occurs first in the development of high-level resistance, the finding of PMQR-harboring isolates without QRDR mutations indicates that the PMQR genes, including several qnrB alleles and aac(6′)-Ib-cr (Table 1), might be acquired before the selection of these mutations.
Genetic environments of the prevalent PMQR genes
The genetic environments of qnrB2, qnrB10, qnrB19, and aac(6′)-Ib-cr were analyzed. In all cases, qnrB2 and qnrB10 were located in the same genetic structures of complex class 1 integrons (variable region 2) that were previously found in Argentina, which have proved to be contained in conjugative plasmids. 23
In 5 out of 7 qnrB19-containing isolates, this gene was found in the ColE1-type plasmid pPAB19-1 that was proposed as a natural reservoir. 23 This plasmid and its close genetic variants were almost exclusively found in E. coli and Salmonella spp. and could be transferred by conjugation to E. coli, even when they are not self-transferable.23,49 Interestingly, besides E. coli and Salmonella spp., here, we found pPAB19-1 in 2 K. pneumoniae isolates, which suggests that the mobilization of this plasmid also occurred in vivo. In the remaining 2 qnrB19-containing isolates (K. pneumoniae), this gene was located in the genetic context of the ISEcp1C-based Tn2012. 29 This is the first report of such a qnrB19 genetic environment in Argentina, which, along with the three previously described, 23 highlights the abundance of genetic elements involved in its mobilization.
In 23 out of 30 (77%) aac(6′)-Ib-cr-harboring isolates, this gene was found as a cassette of class 1 integrons. These included six out of seven isolates in which aac(6′)-Ib-cr was combined with qnrB2 or qnrB10. Therefore, the significant association observed between these genes and aac(6′)-Ib-cr (see section Occurrence and Prevalence of PMQR Genes) might be explained by genetic linkage in complex class 1 integrons. In the other 4 (13%) isolates, aac(6′)-Ib-cr was located in Tn6238, which was only described in K. pneumoniae from Argentina. 31 Interestingly, besides 1 K. pneumoniae, we found Tn6238 in 2 E. cloacae and 1 E. coli, suggesting dissemination wider than expected among clinical enterobacteria. In the remaining three isolates, aac(6′)-Ib-cr was found to be bracketed by IS26 (upstream) and blaOXA-1 (downstream), as described. 30
Concluding Remarks
To the best of our knowledge, this is the first report on the prevalence of PMQR genes in phenotypically unselected clinical enterobacteria where all the genes currently known have been screened. We observed an overall prevalence of 8.1% and a wide distribution of these genes throughout Argentina. One main strength of this work is that it constitutes a large national point prevalence study on PMQR genes. This was based on the enterobacterial collection method used that allowed us to obtain a large number of unselected isolates collected at the same time point from many hospitals all along the country. Although this approach also provided a diverse spectrum of isolates, it had the limitation that some species, such as Salmonella spp., C. freundii, C. koseri, P. vulgaris, and P. stuartii (9, 8, 7, 7, and 4 isolates, respectively), could be underrepresented. Since the isolates were consecutively collected during the hospital routine, the low numbers of isolates of these species in the enterobacterial collection are probably a consequence of their abundance in our clinical settings. Given the high number of isolates tested, we could not perform a molecular subtyping analysis (MLST, repPCR, or PFGE) to address their genetic relatedness, which constitutes another limitation of our study.
In E. coli and Shigella spp., we found that the QRDR genotypes based on single gyrA mutations (S83A or D87G/Y) confer low NAL resistance. Although these genotypes have low prevalence, the fact that they cannot be detected by the current susceptibility breakpoints of ciprofloxacin and levofloxacin for enterobacteria, nor by the pefloxacin-disk test recommended for Salmonella, represents a threat for fluoroquinolone treatment. In this context, it should be stressed that the NAL susceptibility (DD or MIC determination) was very suitable to detect these QRDR genotypes.
Finally, our data provide additional evidence for the coexistence of PMQR genes and QRDR mutations in clinical enterobacteria. Indeed, the fact that 82% of the PMQR-harboring isolates also contained QRDR mutations involves an advantage for the clinical laboratories, since most of the PMQR-harboring enterobacteria can be easily detected and only a few of them (those without QRDR mutations) could represent a challenge to categorize quinolone susceptibility.
Footnotes
Acknowledgments
This work was supported by a grant from ANPCYT (PICT 2007-01804), Buenos Aires, Argentina, to A.P. E.A was supported by fellowships from ANPCYT and CONICET.
WHONET-Argentina participants include the following: Daniela Ballester, Hosp. Piñero; Ana Di Martino, Sanatorio de la Trinidad Mitre; Laura Errecalde, Hosp. Fernández; Angela Famiglietti, Hosp. de Clínicas; Analía Fernández, Hosp. Universitario Fundación Favaloro; Nora Gómez, Hosp. Argerich; Horacio Lopardo, Hosp. de Pediatría Garrahan; Nora Orellana, FLENI; Miryam Vazquez, Hosp. de Niños Gutiérrez; Miriam Mortarini, Hosp. Muñiz (all in Buenos Aires City); Adriana Di Bella, Hosp. Posadas, El Palomar; Marcelo Zotta, Instituto Nacional de Epidemiología Juan Jara, Mar del Plata; Mónica Machain, Hosp. Piñeyro, Junín; Andrea Pacha, Hosp. San Juan de Dios, La Plata; Marisa Almuzara, Hosp. Eva Perón, San Martín; Ana Togneri, Hosp. Evita, Lanús; María Luz Benvenutti, Hosp. Penna, Bahía Blanca, and Cecilia Vescina, Hosp. de Niños Sor Maria Ludovica, La Plata (all in the province of Buenos Aires); Viviana David, Hosp. Interzonal San Juan Bautista, and Patricia Valdez, Hosp. de Niños Eva Perón, Catamarca (province of Catamarca); Norma Cech, Hosp. 4 de Junio-Ramón Carrillo, Pte. Roque Saenz Peña, and Isabel Marques, Hosp. Perrando, Resistencia (province of Chaco); Omar Daher, Hosp. Zonal de Esquel (province of Chubut); Claudia Aimaretto, Hosp. Regional de Villa María; Marina Bottiglieri, Clínica Reina Fabiola; Catalina Culasso, Hosp. de Niños de la Santísima Trinidad; Liliana González, Hosp. Infantil Municipal; Ana Littvik, Hosp. Rawson, and Lidia Wolff, Clínica Privada Vélez Sarsfield, city of Córdoba (all in the province of Córdoba); Ana María Pato, Hosp. Llano, and Viviana García Saitó, Hosp. Pediátrico Juan Pablo II, Corrientes (province of Corrientes); María Moulins, Hosp. Masvernat, Concordia, and Francisco Salamone, Hosp. San Martín, Paraná (province of Entre Ríos); Nancy Pereira, Hosp. Central, and María Vivaldo, Hosp. de la Madre y el Niño, Formosa (province of Formosa); Marcelo Toffoli, Hosp. de Niños Quintana, and Maria Weibel, Hosp. Pablo Soria, Jujuy (province of Jujuy); Gladys Almada, Hosp. Molas, Santa Rosa, and Adriana Pereyra, Hosp. Centeno, Gral. Pico (province of La Pampa); Sonia Flores, Hosp. Vera Barros, La Rioja (province of La Rioja); Lorena Contreras, Hosp. Central, and Beatriz García, Hosp. Pediátrico Notti, Mendoza (province of Mendoza); Ana María Miranda, Hosp. SAMIC El Dorado (province of Misiones); María Rosa Núñez, Hosp. Castro Rendón, and Herman Sauer, Hosp. Heller, Neuquén (province of Neuquén); Néstor Blázquez, Hosp. Carrillo, Bariloche, and Mariela Roncallo, Hosp. Cipolletti (province of Río Negro); María Luisa Cacace, Hosp. San Vicente de Paul, Orán, and Jorgelina Mulki, Hosp. Materno Infantil, Salta (province of Salta); Roberto Navarro, Hosp. Rawson, and Nancy Vega, Hosp. Marcial Quiroga, San Juan (province of San Juan); Ema Fernández, Policlínico Regional J.D. Perón, Villa Mercedes, and Hugo Rigo, Policlínico Central, San Luis (province of San Luis); Alejandra Vargas, Hosp. Regional, Río Gallegos, and Josefina Villegas, Hosp. Zonal de Caleta Olivia (province of Santa Cruz); María Baroni, Hosp. de Niños Alassia, and Emilce de los Ángeles Méndez, Hosp. Cullen, Santa Fe; Isabel Bogado, Facultad de Bioquímica, Noemí Borda, Hosp. Español, and Adriana Ernst, Hosp. de Niños Vilela, Rosario (all in the province of Santa Fe); Ana María Nanni de Fuster, Hosp. Regional Ramón Carrillo, Santiago del Estero (province of Santiago del Estero); Iván Gramundi, Hosp. Regional de Ushuaia, and Marcela Vargas, Hosp. Regional de Río Grande (province of Tierra del Fuego); José Assa, Hosp. del Niño Jesús; and Amalia del Valle Amilaga, Hosp. Padilla, and Humberto Musa, Centro de Microbiología Médica, Tucumán (province of Tucumán).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.