
Abstract
Berberine is a plant-derived alkaloid possessing antimicrobial activity; unfortunately, its efflux through multidrug resistance pumps reduces its efficacy. Cellular life span of Escherichia coli is generally shorter with prolonged berberine exposure; nevertheless, about 30% of the cells still remain robust following this treatment. To elucidate its mechanism of action and to identify proteins that could be involved in development of antimicrobial resistance, protein profiles of E. coli cells treated with berberine for 4.5 and 8 hours were compared with control cells. A total of 42 proteins were differentially expressed in cells treated with berberine for 8 hours when compared to control cells. In both 4.5 and 8 hours of berberine-treated cells, carbohydrate and peptide uptake regimens remained unchanged, although amino acid maintenance regimen switched from transport to synthesis. Defect in cell division persisted and this condition was confirmed by images obtained from scanning electron microscopy. Universal stress proteins were not involved in stress response. The significant increase in the abundance of elongation factors could suggest the involvement of these proteins in protection by exhibiting chaperone activities. Furthermore, the involvement of the outer membrane protein OmpW could receive special attention as a protein involved in response to antimicrobial agents, since the expression of only this porin protein was upregulated after 8 hours of exposure.
Introduction
S
Berberine is a clinically important plant alkaloid derived from the amino acid tyrosine. 4 It is currently utilized in Ayurvedic and Chinese medicine. 5 It possesses no genotoxic activity and is not a potent mutagenic agent in dividing cells. Furthermore, it does not induce any significant mutagenic, cytotoxic, or recombinogenic effects. 6
Berberine has anticancer, 7 antiviral, 8 antifungal, 9 and antibacterial 10 activities and targets a wide range of proteins. Among its reported target proteins are tyrosine phosphatase 1B (PTP1B), 11 arylamine N-acetyltransferase (NAT), 12 HIV-I reverse transcriptase, 13 DNA topoisomerase II, 14 alpha amylase, 15 and lipase. 16 Unfortunately, this natural alkaloid is also a substrate for the multidrug resistance (MDR) efflux pump p-glycoprotein (P-gp), which complicates its clinical applications. 17 On the contrary, the modulation of P-gp transport activity by HIV-protease inhibitors serves as a promising strategy in the administration of berberine. 18
Therapeutic efficacy of berberine against bacteria fosters hope in battling bacterial infections. 19 Studies that focus on the antibacterial action of berberine demonstrated that this alkaloid binds and inhibits FtsZ, a protein that is required for cell division and viability in Escherichia coli. 20 Berberine was also reported to be a potent inhibitor for bacterial NAT 21 and sortase. 22 Additive and/or synergistic effects of berberine with different antibiotics further offer enhanced potency.23,24 However, efflux of berberine by MDR pumps impairs its antibacterial activity against gram-negative bacteria. This could, in part, be overcome by combinatorial use of berberine and MDR pump inhibitors. 25
In our previous study, we tried to elucidate berberine-mediated repression of cell growth in E. coli, integrating proteomic and transcriptomic approaches. The presence of 0.6 × minimum inhibitory concentration (MIC) berberine was found to alter genes involved in cell wall/membrane biosynthesis and transport/motility-related functions following 4.5 hours of exposure. 26 Furthermore, the number of viable E. coli cells was significantly lower, although there were still actively growing cells. In the current study, our motivation was to investigate the changes in E. coli cells following prolonged exposure to berberine, 8 hours, to elucidate the mechanistic details of its mode of action and identify proteins that could be involved in stress development.
Materials and Methods
Chemicals
All chemicals, reagents, and kits used in the current study were obtained from Bio-Rad (Hercules, CA), Merck, Sigma, Qiagen (Valencia, CA), or Fermentas. Berberine was purchased from Sigma-Aldrich. Sterile dH2O was used as the solvent for berberine.
Bacterial strain and growth conditions
The model microorganism E. coli K12 (ATCC 10789) was obtained from the Turkish Scientific and Technological Research Institute—Institute for Genetic Engineering and Biotechnology (Tubitak-Gebi). Cells were cultured in Luria Bertani (LB) medium (per liter: 10 g tryptone, 5 g yeast extract, 10 g NaCl) at 37°C and 180 rpm. Two parallel cultures (control and berberine treated) were inoculated from an overnight grown preculture. 0.6 × MIC (750 μg/ml) berberine was added to the growth medium at the time of inoculation. The MIC value of berberine for E. coli cells was taken from a study by Ozbalci et al. 27 Following 8 hours of growth, cells were harvested by centrifugation at 6,000 rpm and 4°C for 20 minutes and then washed with 50 mM Tris buffer (pH 7.8). Cells were kept at −80°C for later use.
Protein sample preparation and proteomic analysis
Protein extraction was achieved using the ProteoPrep® Total Extraction Sample Kit (Sigma PROT-TOT) protocol. Protein extraction was performed from at least three independently grown cultures. Impurities were removed with the ReadyPrep™ 2-D Cleanup Kit (Bio-Rad). Protein concentration was determined by the Bradford method. 28
Extracted proteins were separated by two-dimensional (2D) gel electrophoresis using immobilized pH gradient (IPG) strips of 17 cm (pH of 4–7; Bio-Rad) in the first dimension and sodium dodecyl sulfate/polyacrylamide gel electrophoresis in the second dimension. 29 Reagents used in 2D gel electrophoresis were from the ReadyPrep 2-D Starter Kit (Bio-Rad). The CBB-G250 dye method 30 was used to visualize protein spots on 2D gels, and the GE HEALTHCARE® Image Scanner II system was used for scanning the stained gels. After alignment of gel images, the same protein spots were detected by background subtraction, normalization, and matching. Spot lists were statistically ordered by p-value from the one-way ANOVA. Top ranking protein spots with a cutoff value of greater than twofold expression and a p-value of <0.05 were accepted by going through the spot rank table.
Differentially expressed protein spots were excised from triplicate 2D gels. Protein identification was performed at the Université de Liége (Liége, Belgium; GIGA Proteomics Facility) based on tandem mass spectrometry fragmentation data, which filtered matches to accept only peptides with a score of 0.05 or lower. The obtained spectral data were then searched against a subset of the EcoProt database (4,284 sequences; 1,351,035 residues).
Total RNA extraction, complementary DNA synthesis, and qualitative polymerase chain reaction analysis
RNA was stabilized using RNA Protect Bacteria Reagent (Qiagen). Extraction of total RNA was achieved based on the RNeasy mini kit (Qiagen) protocol. Using RNase-free DNase set (Qiagen), on-column DNase digestion was performed to remove genomic DNA. RNA concentration was measured with the Qubit® 2.0 fluorometer (Invitrogen). Complementary DNA (cDNA) synthesis was achieved with the QuantiTect® Reverse Transcription Kit (Qiagen).
Qualitative polymerase chain reaction (qPCR) analyses were carried out in LightCycler 1.5 Instrument (Roche Applied Science) with SYBR Green I (LightCycler® FastStart DNA MasterPLUS SYBR Green I; Roche Applied Science). Primers used to amplify each target gene are listed in Table 1. Reaction conditions for gene amplification were 95°C for 10 minutes for denaturation followed by 40 cycles of 95°C for 10 seconds, 57°C for 10 seconds, and 72°C for 45 seconds. Primer dimer formation and other artifacts were cross checked by melting curve analysis. 16S ribosomal RNA (rRNA) was selected as the housekeeping gene for data normalization. Fold change in each target gene was calculated by the 2−ΔΔCT method of Livak and Schmittgen. 31
Housekeeping gene.
Data analysis of proteins
Functional information of differentially expressed proteins was retrieved from the Universal Protein Resource Knowledgebase (UniProtKB; www.uniprot.org/). 32
Metabolic pathway analysis for the differentially expressed enzymes was performed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) PATHWAY database. 33
Analysis with scanning electron microscopy
Twenty-five to 30 μl of cell suspension (control or berberine treated) was carefully placed onto a 0.22-μm-pore-sized membrane filter and fixed for 1.5 hours by floating the filter on 2% glutaraldehyde. After fixation, the filter was washed thoroughly with water. The filter was then dehydrated in increasing concentrations of ethanol, 30%, 50%, 70%, and 90% (10 minutes each), and finally kept in absolute ethanol for 1 hour. The filter was air-dried overnight and then was coated with platinum sputter coater before imaging. Imaging was performed with Philips XL30 ESEM-FEG/EDAX system under high vacuum mode with 5 kV energy and 3.0 spot size.
Results
Regulation of proteins under prolonged berberine stress
A total of 61 different protein spots were analyzed. Images of 2D gels from control and berberine-treated cells are given in Fig. 1. Results of protein identification have shown that 42 proteins were differentially expressed after prolonged berberine exposure (Table 2). Of these proteins, 28 were upregulated and 14 were downregulated. Information regarding the biological processes of these proteins was retrieved from UniProtKB database.
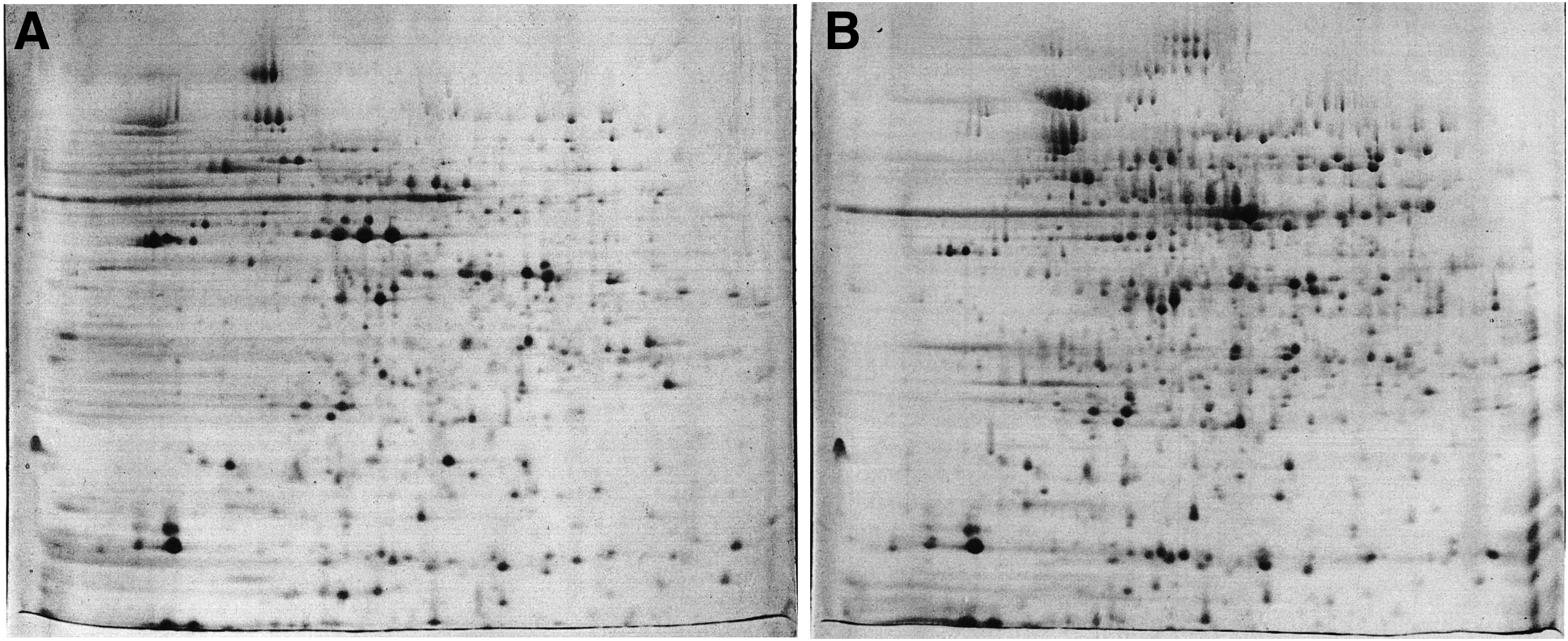
Proteome maps of untreated
UniProtKB, Universal Protein Resource Knowledgebase.
A number of proteins were identified as multiple spots on 2D gels, probably due to post-translational modifications. 34 Among these proteins were MalE (spots D2, D3, D7), OmpC (spots D4, D4f, D6), OmpF (spots D9, D11), AcnB (spots P11, P12), AspA (spots P24, P25), GlyA (spots I21, I22), LpdA (spots I10, I18, I20), Tsf (spots I3, I16), FusA (spots P13, P14), ProS (spots P21, P22), AckA (spots I24, I25), AtpA (spots I15, I19), and AtpD (spots I13, I14). This is not unexpected as various other research groups reported similar findings for some of these proteins, such as MalE, OmpF, LpdA, and Tsf, under different conditions.35,36
Expression analysis of selected proteins by qPCR
Five proteins with different regulation patterns (TufA, LamB, MalE, Glk, and Crr) were selected for qPCR analysis to verify the accuracy of the proteomic findings. Melting curve analysis confirmed the specificity of the primers and the absence of dimer formation. qPCR analysis supported our proteomic results with a 6.3-fold increase in tufA and ∼20-fold decrease in lamB. The change for malE could not be numerically expressed since its expression in the presence of berberine was too low to be calculated by the method of Livak and Schmittgen. 31 As expected, the expression of two genes of two different paths of glucose utilization, crr and glk, decreased in the presence of berberine (Table 3).
Pathway analysis of proteins
KEGG pathway database was used to visualize the altered metabolic pathways on prolonged berberine stress (Fig. 2). Nine pathways were affected and enzymes of the affected pathways were mainly clustered in the central carbon metabolism (glycolysis/gluconeogenesis, citric acid cycle) and amino acid metabolisms (glycine, serine, threonine, cysteine, methionine, alanine, aspartate, and glutamate). Proteins that are not enzymes were not depicted in this figure.

Pathway maps of differentially expressed enzymes in 8-hour berberine-treated E. coli. Snapshot of each particular pathway is retrieved from KEGG Pathway database. 33 Differentially expressed gene names are written in bold and indicated with thick arrows. KEGG, Kyoto Encyclopedia of Genes and Genomes.
Visualization of E. coli by scanning electron microscopy
Scanning electron microscopy was used to visualize the structural changes in the cell walls. Images obtained from control and berberine-treated cells are given in Fig. 3. After 8 hours of growth, cell walls in the control culture started to deteriorate and acquired rough surfaces (Fig. 3A). The sizes of these cells were typically between 1.5 and 2 μm (Fig. 3C). On the contrary, cell membranes after treatment with berberine were smooth and seemed to be intact (Fig. 3B). Furthermore, these cells were longer than normal E. coli cells, about 2.5–3.5 μm in size (Fig. 3D).
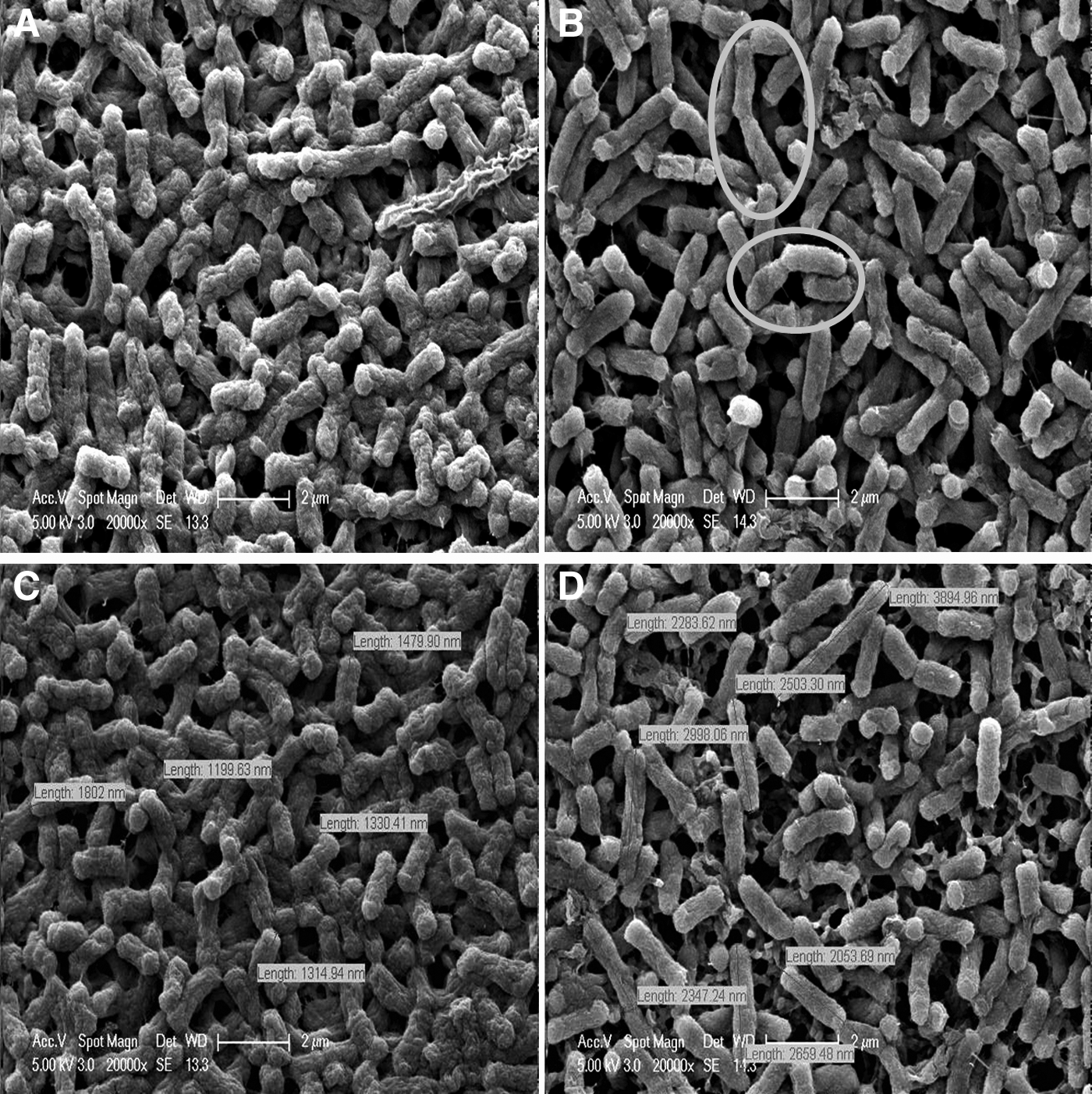
Images obtained from scanning electron microscopy,
Discussion
Berberine is primarily known as an alkaloid that binds DNA, with preference for AT-rich sequences. Recent studies showed that berberine targets not only DNA but also several proteins. 37 In our previous work, we have tried to elucidate the antimicrobial effect of berberine in E. coli cells following 4.5 hours of growth in its presence. Proteomic analysis showed that carbohydrate and amino acid metabolisms were repressed and major outer membrane proteins were downregulated, suggesting that membrane integrity may be compromised. Furthermore, after 4.5 hours of exposure to berberine, viable cell number was 50% lower compared to the control culture.
Apparently, the presence of berberine reduced the number of viable cells after 4.5 hours of exposure; however, it was only after 6 hours that growth ceased in the presence of berberine (Fig. 4). After 8 hours of exposure to berberine, viable cell number was 70% lower compared to the control culture. Taking this as the starting point, the motivation for this work was to identify target proteins that could trigger stress development against berberine by their continued presence during prolonged berberine exposure. We interpreted berberine stress-related changes comparatively with the results of the 4.5-hour berberine-exposed cells. 26 Changes in 4.5-hour-treated cells relevant to the discussion of the current study are taken from a study by Karaosmanoglu et al. 26 and summarized in Table 4.
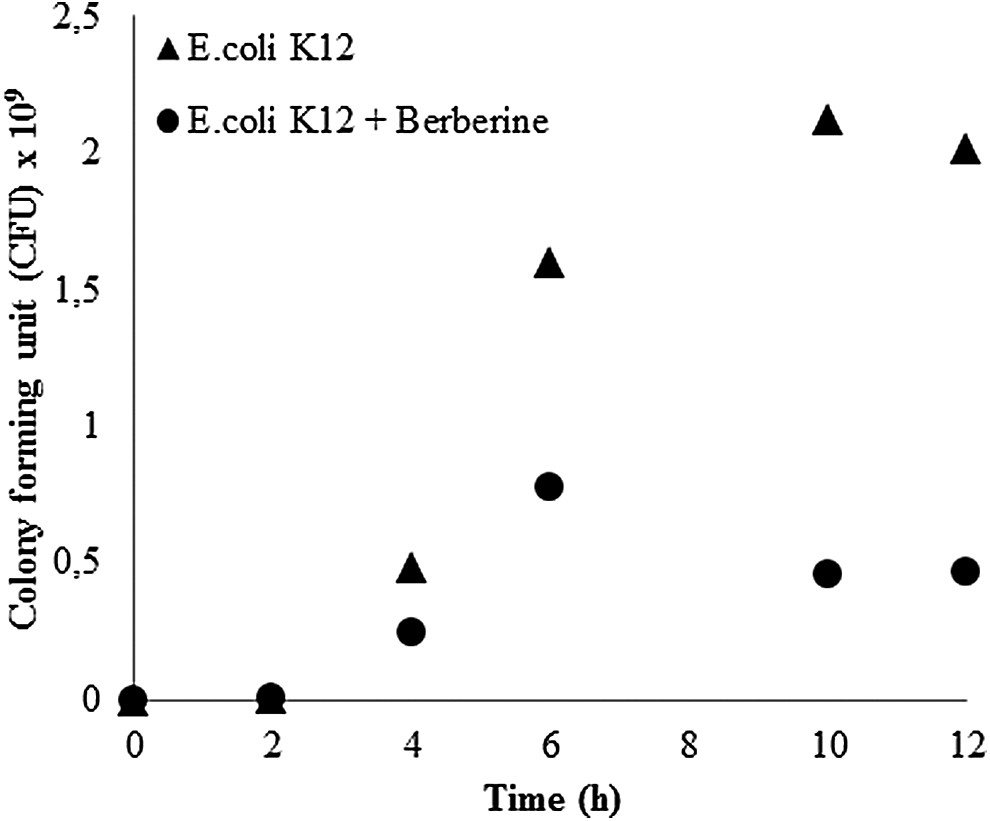
Viable cell numbers of E. coli K12 cells in the presence and absence of berberine. Plots were reproduced from the previous data. 26
Common at 4.5 and 8 hours of berberine treatment.
UspA, universal stress protein A.
Defect in division persists with prolonged exposure to berberine
Berberine is a well-known DNA ligand. Consequently, the general expectation is the induction of the SOS response, which is activated by DNA damage. However, Boberek et al. 20 reported that berberine does not induce SOS response in E. coli and suggested cell division protein FtsZ as the major target of berberine. 20 Our results also showed that, the abundance of FtsZ was constantly low in berberine-treated samples. Also, the cell division inhibitor gene, slmA, was upregulated after 4.5 hours of exposure to berberine. Both these results suggested that berberine action involves inhibition of cell division.
Further clues for growth arrest were obtained by electron microscopy. In Fig. 3D, berberine-treated E. coli cells were longer. Moreover, cells that encounter difficulty in division after berberine treatment can clearly be visualized. These cells are marked with circles in Fig. 3B.
To the contrary, the gene ftsK, which colocalizes with FtsZ to the septal ring structure, was upregulated in 4.5-hour berberine-exposed cells. This is an essential protein in cell division and chromosome partitioning. 38 The expression of ftsK has previously been associated with the SOS response through RecA. However, with cells in growth arrest condition and in the absence of the SOS response, the induction of ftsK should be independent of cell division. 39 Indeed, Diez et al. 38 reported that FtsK levels increase not only as a response to the demands of the cell division apparatus but also in response to DNA damage.38,40
Universal stress protein A could not contribute to resistance development
Multiple members of the universal stress proteins are reported to cooperate in defense against DNA damage.41,42 Furthermore, there is evidence that a change in the level of FtsK transduces DNA damage signal to the universal stress protein A (UspA) promoter, which modulates uspA transcription. In fact, overexpression of the C-terminal half of FtsK has been shown to increase the activity of UspA. 40 The usp system responds to DNA aberrances, although not as a part of classical SOS response. 39 Could ftsK be involved in inducing uspA in response to DNA anomalies? As a DNA ligand, the presence of berberine could trigger ftsK expression. However, this was not the case since UspA remained low in abundance in berberine-exposed cells and this condition persisted.
The low abundance of UspA could be explained with repression in the expression of cold shock proteins, CspC and CspE. Phadtare and Inouye reported that messenger RNA (mRNA) stability decreased drastically in the ΔcspC ΔcspE mutant and increased in a strain that overproduced CspC. 43 CspC and CspE seem to be critical elements in regulating the expression of uspA of the complex stress response network of E. coli. Indeed, both cspC and cspE were downregulated in 4.5-hour berberine-exposed cells.
Elongation factors are involved in protection from stress
As 8-hour berberine-exposed cells entered death phase, parts of the translational machinery were induced. Not only subunits of 30S ribosomal protein, RpsA and RpsF, but also four elongation factors (EFs) Tu, Ts, G, and P (EF-Tu, EF-Ts, EF-G, and EF-P) encoded by tufA, tsf, fusA, and efp, respectively, increased in abundance. However, the only noteworthy change in the translational machinery after the 4.5-hour berberine exposure was an increase in the 50S ribosomal subunits. Efficient and coordinated synthesis of proteins is required for growth, motility, and stress survival, and thus, activation of the translational machinery is not unexpected. However, we thought that some other function could also be attributed to EFs.
In bacteria, the elongation phase of translation is normally governed by the EFs EF-Tu, EF-Ts, EF-G, and EF-P. EF-Tu accelerates aa-tRNA binding to the ribosome, whereas EF-Ts is the guanine nucleotide exchange factor of EF-Tu. EF-Tu is a member of the GTPase family, which provides energy from GTP hydrolysis during protein folding. 44 Thus, its upregulation, as predicted, was accompanied by an increase in the abundance of proteins involved in guanine nucleotide synthesis, PurC and GuaB. On peptide bond formation, EF-G translocates mRNA and EF-P alleviates ribosome stalling during synthesis of proteins containing consecutive Pro residues, possibly by augmenting the peptidyl transferase activity of the ribosome. 45 All of these EFs are indispensible for the elongation phase of protein synthesis. However, recent studies by independent groups have suggested the involvement of EFs in stress response with chaperone activities.46,47 Caldas et al.48,49 reported the interaction of E. coli EF-Tu and EF-G with unfolded and denatured proteins to be similar to molecular chaperones that function in protein folding and protein renaturation. Han et al. 50 showed EF-Ts as a stress-induced protein with a critical role in prevention of inclusion body formation. Furthermore, EF-Ts increased the solubility of an aggregation-prone protein when used as an N-terminus fusion partner. 50 EF-P has been reported to be a nonessential gene 51 and its biological function remains enigmatic. Interestingly, some proteins necessary for stress tolerance seem to require EF-P for synthesis. Change in the abundances of these proteins renders cells susceptible to stress, eventually leading to the disruption of the bacterial outer membrane integrity. 52 The clues that support chaperone-like properties of EFs suggest that these proteins are involved in stress response.
Membrane acts as an important barrier in protection from berberine
Biological membranes have a major influence on the susceptibility of microorganisms to different compounds. Regulation of solute trafficking through pore-forming porins is one mechanism used by cells to keep toxic molecules outside. However, some drug molecules can simply diffuse across the lipid bilayer. There is strong proof that membrane lipid composition is modified in response to different drugs. 53
Consistent with previous reports, our previous experimental findings showed that a number of genes involved in membrane phospholipid biosynthesis (pssA, pgpA, and opgB), together with fadK, were upregulated after 4.5 hours of exposure to berberine. 26 FadK catalyzes the esterification of exogenous fatty acids either for subsequent degradation or for incorporation into phospholipids; hence, its upregulation should be a direct indication of modulation of the membrane phospholipids. The general tendency toward fatty acid degradation with the downregulation of fadR, as reported previously, 26 should also be correlated with membrane modifications, although utilization of fatty acids as carbon sources cannot be disregarded at this stage because of carbon source storage. Another modification in the membrane lipid structure was induced by the upregulation of the pagP gene, which encodes an enzyme that alters the lipid A of the outer membrane. By altering the lipid A composition of the outer membrane, the PagP enzyme renders microorganisms less susceptible to the membrane permeabilizing effects of antimicrobial peptides. 54 The changes induced by the upregulation of this interesting outer membrane protein may contribute to protection from berberine.
Extending the berberine exposure period to 8 hours leads to a change in the integrity and robustness of the membranes. One of the changes is the increase in the abundance of the β barrel-like outer membrane protein, LptD. This protein is involved in assembling the lipopolysaccharides at the surface of the outer membranes. 55 On its depletion, E. coli cells exhibit abnormal membrane structures. 56 Based on the functional interaction between lipopolysaccharide and protein assemblies, 55 the increase in the abundance of the protein essential in the assembly and insertion of outer membrane β-barrel proteins (BamA),57,58 together with LptD, could be an evidence for the increase in the fitness of the cells exposed to berberine.
The change in the permeability of the membrane is accompanied by a change in membrane phospholipid composition. There is continuous phospholipid breakdown, as suggested by the increase in the abundance of glycerophosphoryl diester phosphodiesterase (GlpQ), and at the same time, there is continuous fatty acid synthesis as suggested by the increase in the abundance of the alpha subunit of acetyl-coenzyme A carboxylase carboxyl transferase (AccA). The enzyme GlpQ is involved in the utilization of the glycerol moiety of phospholipids and triglycerides during its degradation, whereas AccA is the enzyme that catalyzes the first step of malonyl-CoA from acetyl-CoA in fatty acid synthesis.
Cyclopropane fatty acid formation is a postsynthetic modification of bacterial membrane lipid bilayers seen in many bacteria as they enter the stationary phase. 59 The presence of cyclopropane fatty acids has been proposed to decrease permeability to H+, and to provide protection for membranes against phospholipid degradation.60,61 The changes in GlpQ and AccA could contribute to protective modifications in the membrane phospholipid composition that lead to the accumulation of cyclopropane fatty acids. However, since these cells are already in their stationary phase, these modifications could also be a part of their expected life span. The smooth and intact surfaces of the berberine-exposed cells (Fig. 3) are a further indication of the cell membrane/cell wall modifications for survival.
Carbohydrate and peptide uptake remains unchanged but amino acid maintenance regimen switched from transport to synthesis with prolonged berberine exposure
One of the striking effects of berberine was the significant decline in carbohydrate uptake, as evidenced by downregulation of the major outer membrane porin proteins, OmpC, OmpF, and LamB, after 4.5 hours. 26 OmpC and OmpF are nonspecific porins, whereas LamB is specific for maltose and maltodextrins. 62 Restriction of the outer membrane permeability to nutrient influx continued as the exposure period to berberine was extended. There is strong evidence on the penetration of various antibiotics through these porins. 63 Moreover, their regulation in the presence of a variety of antibiotics has been demonstrated in resistant E. coli strains.64–69 Our results suggest that cells avoided berberine influx by regulating the expression of porin proteins.
Limited permeability for carbohydrates has automatically led to a decrease in the abundances of proteins involved in glucose transport (Crr), maltose transport (MalE), galactose transport (MglB), and D-ribose transport (RbsB) after 8 hours of exposure. The simultaneous decrease in the expression of glucokinase and Enzyme IIAGLC of two different glucose import pathways clearly shows intracellular glucose shortage. This condition is similar to that found for 4.5 hours of berberine exposure.
In contrast to carbohydrate uptake, cells showed variability in their preference for amino acid transport and metabolism as the period of berberine exposure increased. After 4.5 hours of growth, amino acid synthesis decreased while transport increased. While some amino acids, such as arginine and lysine, require OmpC and OmpF for transport, others such as glutamate move across the outer membrane via other routes. 70 Therefore, amino acid influx that was independent of OmpC and OmpF was probably sufficient to compensate for the amino acids that required OmpC and OmpF. Amino acid interconversion reactions could have counterbalanced amino acids with limited influx rates. With prolonged exposure of the eight proteins with changing abundances, three belonging to amino acid transport decreased, whereas five belonging to synthesis increased. This may be due to the depletion of free amino acids, leading to a switch from transport to synthesis.
The outer membrane protein OmpW deserves special attention as a protein involved in antimicrobial resistance
Similar to OmpC and OmpF, OmpW also belongs to the family of small outer membrane proteins in gram-negative bacteria. There are reports on its involvement in the transport of small hydrophobic molecules; nevertheless, its function is still largely uncharacterized. 71
Recent proteomic evidence suggested a correlation between the presence of OmpW in cells and bacterial resistance to antibiotics. Interestingly, there is no consent on the regulation pattern of this protein. Its abundance has been reported to vary depending on the antimicrobial agent used. Antibiotics, ampicillin, tetracycline, nalidixic acid, and chloramphenicol, all increased the abundance of OmpW, whereas in the presence of chlortetracycline and streptomycin, OmpW was downregulated.64–68,72 In the present work, the abundance of OmpW increased when the period of berberine exposure increased. Interestingly, its expression was downregulated following 4.5 hours of berberine exposure. Therefore, we believe that this protein could have a critical role in stress development.
Spermidine uptake is tightly regulated
Polyamines (putrescine, spermidine, and spermine) are important molecules with a variety of biological functions, including binding to nucleic acids, stabilization of membranes, stimulation of enzymes, and involvement in stress response. Their presence positively correlates with growth rate but their intracellular concentrations should be fine-tuned since they are toxic when found in large quantities. They are also reported to be used as carbon and nitrogen sources by E. coli. 73
The ABC transport system PotABCD is involved in polyamine transport, displaying preference for spermidine. Polyamine biosynthesis and transport mechanisms are intricately linked with cellular functions; hence, polyamines are absolutely required for growth. Intracellular spermidine could further act as a precursor for the synthesis of other polyamines. Transcriptional evidence from 4.5 hours of exposure to berberine showed that two genes of this operon (potB/C) were upregulated. In a nutrient-limited state due to reduced permeability, it is plausible to think that polyamines may serve as a carbon or nitrogen source; however, the downregulation of two genes (puuA, puuD), absolutely essential to metabolize putrescine, weakened this hypothesis. The increase in polyamine transport could be related to the necessity for regulating cellular functions. On the contrary, their continued accumulation should not be disregarded, as their toxicity could be one of the reasons of cell death.
Interestingly, as berberine exposure was extended to 8 hours, the abundance of PotD of the potABCD gene cluster was downregulated. PotD is the periplasmic binding protein for spermidine uptake as well as being an inhibitor of spermidine uptake 74 ; therefore, it is difficult to interpret its exact role under the current condition. Its downregulation is more likely to be a sign for the depletion of spermidine supply in the growth medium but requires further investigation.
Rates in energy production and metabolism increase
Under prolonged berberine stress, energy requirement increased, as indicated by increases in the abundances of proteins found in glycolysis/gluconeogenesis and citric acid cycle. Among these proteins were those encoded by the genes fbaA, tpiA, lpd, and ackA. Furthermore, two subunits of the ATP synthase, AtpA and AtpD, increased in abundance. This finding may suggest that cells have a higher energy demand during prolonged berberine stress. However, we believe that this condition could also be independent of drug stress and could be related to the growth phase of the cells, as cells are approaching death phase.
Conclusions
Proteomics is an attractive technique applied in antibacterial drug discovery and elaboration of antimicrobial mechanisms and bacterial resistance to antibiotics. 75 In our previous work, we tried to elucidate the mechanistic details of the antimicrobial action of berberine in E. coli cells following 4.5 hours of exposure. With this treatment of berberine, viable cells were reduced by 50%. Further exposure to berberine dropped the number of viable cells by 70% after 8 hours. Motivated by the presence of viable cells after this period, possible mechanisms of stress development that lead to an increased cellular fitness in berberine-exposed cells were pursued. Carbohydrate and peptide uptake regimens remained unchanged with prolonged berberine exposure, but amino acid maintenance regimen switched from transport to synthesis. Interestingly, universal stress proteins did not contribute at all to protection. The significant increase in the abundances of EFs could suggest a participation of these proteins as chaperones in stress response. Therefore, they could constitute possible targets against resistance development. Furthermore, the involvement of the outer membrane protein, OmpW, could receive special attention as a protein involved in antimicrobial resistance, since it was only this porin protein that was upregulated following 8 hours of exposure to berberine.
Footnotes
Acknowledgments
We thank Ayse Ogan and Bulent Mertoglu for letting us use their facilities. This work was supported by the TUBITAK Research Grant 108M597 and Marmara University Research Foundation FEN-E-130313-0077. N.B.G. was supported by the TUBITAK Graduate Fellowship (BIDEB).
Disclosure Statement
No competing financial interests exist.