
Abstract
Carbapenem-resistant Gram-negative pathogens have progressively disseminated to different countries worldwide, presenting a serious public health concern. The aims of this study were to determine the prevalence of carbapenem resistance in Gram-negative bacteria in Lebanon, to elucidate molecular mechanisms, and to identify genetic relatedness of incriminated strains. Carbapenem nonsusceptible Enterobacteriaceae, Acinetobacter baumannii, and Pseudomonas were collected from 11 Lebanese hospitals in 2012. Antimicrobial susceptibility was assessed with phenotypic tests, genes encoding carbapenemases were screened via PCR-sequencing, and genetic relatedness was examined by PGFE and ERIC-PCR. A total of 398 nonrepetitive carbapenem nonsusceptible isolates were studied, of which 44 were Enterobacteriaceae, 142 were A. baumannii, and 212 were Pseudomonas. Among Enterobacteriaceae, 70.4% carried blaOXA-48-like gene on IncL/M-type plasmids, while acquired AmpC cephalosporinases, extended-spectrum-β-lactamases, and efflux-pump were additional contributors to carbapenem resistance. Among A. baumannii, 90% produced OXA-23 and GES-11 and carried insertion sequence ISAba1 upstream and adjacent to blaOXA-23 and blaAcinetobacter-derived cephalosporinases. Among Pseudomonas, 16% harbored VIM-2, 4.2% IMP-2, and 1.4% IMP-1 metallo-β-lactamases. Fingerprint analysis indicated that the spread of OXA-48-like carbapenemases was mostly mediated by horizontal transfer, while OXA-23 and GES-11 diffusion in A. baumannii and VIM-2 diffusion in P. aeruginosa were primarily due to clonal dissemination. This study is the first nationwide investigation of carbapenem resistance in Lebanon, showing low level of resistance in Enterobacteriaceae, and higher levels in A. baumannii and Pseudomonas. With current changes in the region, continuous surveillance of carbapenem resistance is crucial.
Introduction
T
Clinically important Gram-negative bacteria are endowed with continuously progressing enzymatic and nonenzymatic passageways for development of carbapenem resistance. Enterobacteriaceae become carbapenem resistant via (i) secretion of a carbapenemase belonging to Ambler class A (like KPC), class B (like IMP, VIM, or NDM), or class D (OXA-type carbapenemases); and/or (ii) production of a β-lactamase with nonsignificant carbapenem-hydrolyzing activity (ESBL or derepressed AmpC cephalosporinase of Ambler class C) in addition to porin loss or efflux pump overactivity. 5
In Acinetobacter baumannii, carbapenem resistance is mainly due to acquired oxacillinases of subfamilies OXA-23, OXA-24/33/40, OXA-58, and OXA-143. Other carbapenemases have been also identified, 6 namely of Ambler classes A and B.7–9 Hyperproduction of the intrinsic OXA-51-like enzymes, drug extrusion by AdeABC efflux pump, 10 and reduced expression of outer membrane porins11–13 may additionally contribute to a low level of carbapenem resistance in this organism. The effectiveness of carbapenems against infections caused by Pseudomonas has been well compromised mostly by loss or alteration of the outer membrane protein OprD, followed by the production of metallo-β-lactamases, KPC carbapenemase, and overexpression of efflux pumps. 14 Furthermore, carbapenem resistance determinants in Gram-negative bacteria may be subject to upregulation by mobile elements. For instance, the insertion of the mobile element IS1999 of the transposon Tn1999 was found to be associated with mobilization and expression of the carbapenem-hydrolyzing oxacillinase OXA-48 in Enterobacteriaceae. 15 Likewise, in A. baumannii, presence of insertion sequence ISAba1 upstream from blaOXA-23, blaOXA-58, blaOXA-51, and the genes coding for Acinetobacter-derived cephalosporinases (ADCs) provides an efficient mean to promote their overexpression. 16
Reports on the presence of carbapenem-resistant Gram-negative pathogens in the Middle East have been published from Turkey, Israel, Egypt, Kuwait, Saudi Arabia, UAE, and Lebanon. 17 Historically, Lebanon has been a country with very limited restriction on drug use, including antibiotics, with unchecked access to these medications and absence of tight regulations on their exploitation. 18 Given this, and in light of recent mobility and demographic alterations in the region due to civil war, profound consequences on evolution and movement of antibiotic resistance genes are expected. In that context, the magnitude and mechanisms of carbapenem resistance have not been evaluated previously on a nationwide scale.
The objectives of the current study were (i) to estimate the prevalence of carbapenem-resistance in isolates of Enterobacteriaceae, A. baumannii, and Pseudomonas in Lebanese hospitals; (ii) to identify the types of carbapenemases present in these isolates, and, possibly, characterize other mechanisms of carbapenem resistance, through both phenotypic and genotypic techniques; and (iii) to determine the genetic relatedness of carbapenemase-producing isolates from various Lebanese hospitals. This work was presented at the 55th ICAAC meeting (Interscience Conference on Antimicrobial Agents and Chemotherapy), September 17–21, 2015, San Diego, California, abstract number C-1114.
Materials and Methods
Bacterial isolates
Carbapenem-resistant Enterobacteriaceae, Pseudomonas, and A. baumannii were collected from January till December of the year 2012, from 11 tertiary care centers distributed over 5 Lebanese districts as follows: Beirut (Hotel Dieu de France, Saint-George hospital, Clinique du Levant); Mount Lebanon (Arz Hospital, Bellevue Medical Center); North Lebanon (Mounla Hospital); South Lebanon (Secours Populaire Libanaise, Labib Medical Center); and Bekaa (Farhat Hospital, Chtaura Hospital, Bekaa Hospital).
Isolates were collected irrespective of patient history, specimens, or ward, and they were eligible for inclusion if they met criteria of the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines on carbapenem intermediate/resistant properties. 19 Accordingly, Enterobacteriaceae isolates with inhibition zone diameter of imipenem equal to or smaller than 22 mm, or of ertapenem equal to or smaller than 25 mm, were included. A. baumannii and Pseudomonas isolates with imipenem inhibition zone diameters equal to or smaller than 23 mm and 20 mm respectively were included.
The identity of Enterobacteriaceae strains was confirmed by API20E and that of A. baumannii and Pseudomonas by API20NE. A. baumannii species was confirmed using PCR to detect intrinsic blaOXA-51-like gene. 20 Bacterial strains were received at the microbiology laboratory of the School of Pharmacy of Saint-Joseph University where the purification, archiving, and phenotypic tests were performed. Then, PCR-sequencing and clonality studies were performed at the Rodolphe-Mérieux Laboratory of Beirut and at the School of Pharmacy of Paris Descartes University.
Antibiotic susceptibility testing and cloxacillin test
Antimicrobial susceptibility testing was performed on Mueller–Hinton agar by standard disk diffusion method according to EUCAST 201219 guidelines. Inhibition zone diameters of amoxicillin/clavulanic acid (AUG), ceftazidime (CAZ), cefotaxime (CTX), cefepime (CPM), cefoxitin (FOX), aztreonam (ATM), imipenem (IMI), gentamicin (GM), tobramycin (TN), amikacin (AK), ofloxacin (OFX), ciprofloxacin (CIP), and levofloxacin (LEV) were measured. For Pseudomonas, discs of piperacillin (PRL), ticarcillin (TC), meropenem (MEM), and colistin (CO) were also tested and CLSI 2012 21 guidelines were used to assess susceptibility to colistin. All antibiotic discs were from Mast Diagnostics. The double-disc synergy test (DDST) was applied for phenotypic detection of ESBLs. 22 After overnight incubation, an expansion of the inhibition zone diameters around one of the third (CTX and/or CAZ) and/or fourth (FEP) generation cephalosporin and/or imipenem disks toward the amoxicillin/clavulanic acid disk indicated the presence of an ESBL.23,24 Also, minimal inhibitory concentration (MIC) of ertapenem was measured for all Enterobacteriaceae isolates using Etest strip (Liofilchem) on Mueller–Hinton agar plates.
Antibiotic susceptibility testing was additionally performed on all isolates using Mueller–Hinton agar with cloxacillin (25–500 μg/L) to inhibit AmpC cephalosporinases. An increase in inhibition zone diameters of cephalosporins in presence of cloxacillin was considered positive for AmpC production. 23 The DDST was again applied in presence of cloxacillin to eliminate phenotypic effect of AmpCs in certain strains and favor visualization of ESBLs.
Phenotypic detection of carbapenemases and efflux pump activity
A modification of the Hodge test was performed to screen for carbapenemase production in Enterobacteriaceae as previously described by Lee et al. 25
To test the production of Ambler class B carbapenemases (metallo-β-lactamases), which are inhibited by metal ion chelators like EDTA, a combined Etest of imipenem/imipenem+EDTA (Liofilchem) was used on Mueller–Hinton agar according to the manufacturer's instructions. A ratio of MICimipenem/MICimipenem+ EDTA ≥ 8 was considered to be a phenotypic indication of metallo-β-lactamases. 23 This Etest for metallo-beta-lactamases was performed in all Enterobacteriaceae, A. baumannii, and Pseudomonas isolates.
Efflux pump inhibitor tests were performed for isolates of Enterobacteriaceae using the efflux pump inhibitor phenylalanin arginine-β-naphthylamide hydrochloride as previously described. 26 Escherichia coli ATCC 25922 was used as an internal control.
Molecular detection of carbapenemases
Genomic DNA was extracted from samples by boiling method, and stored at −20°C. Polymerase chain reactions (PCRs) and sequencing for β-lactamase-encoding genes were routinely performed using a set of in-house primers as designed by Institut Pasteur, France. 27 Routine testing was performed for carbapenemases of Ambler class A (blaKPC and blaGES) and class B (blaIMP-1, blaIMP-2, blaVIM, and blaNDM). For Ambler class D carbapenemases, blaOXA-48-like gene was tested in Enterobacteriaceae while blaOXA-23, blaOXA-24, and blaOXA-58 were tested in A. baumannii. Enterobacteriaceae isolates were screened for ESBL-encoding genes, blaCTX-M group 1 and blaSHV, and for acquired AmpC cephalosporinases of classes blaACC, blaFOX, blaMOX, blaCMY, blaDHA, and blaLAT.
PCRs detecting insertion sequence IS1999 in blaOXA-48-like harboring Enterobacteriaceae and ISAba1 in A. baumannii were also performed as previously described by Aubert et al. 28 and Turton et al. 16 The blaOXA-48-like harboring Enterobacteriaceae were tested for the presence of IncL/M-type plasmids using primers designed by Poirel et al. 29 The position of ISAba1 in A. baumannii with respect to genes encoding ADCs, OXA-51, and OXA-23 was located using PCR mapping, where ISAba1 forward primer was used with reverse primers for ADCs and OXA-23. All PCR primers and cycling conditions are listed in Supplementary Tables S1–S3; (Supplementary materials are available online at www.liebertpub.com/mdr).
Analysis of genetic relatedness
To obtain genetic relationships among blaOXA-48-like harboring Enterobacteriaceae, Pulsed Field Gel Electrophoresis (PFGE) was performed at the School of Pharmacy of Paris Descartes University. Genomic DNA digested with XbaI (Roche) was subjected to PFGE using a clamped homogeneous electric-field apparatus (CHEF-DRII, Bio-Rad). Bionumerics (Applied Math) was used to establish a similarity matrix for DNA based on calculation of Dice coefficient (pairwise comparison of strains). A dendrogram was generated with unweighted pair group using arithmetic means (UPGMA) hierarchical algorithm. Gels were compared using Staphylococcus aureus NCTC 8325 as reference strain. PFGE patterns were interpreted according to international recommendations used to determine genetic relatedness of a group of organisms, as described by Tenover et al., 30 and applying cutoff value of 80% for genetic relatedness.
Genetic similarity of all A. baumannii and the VIM-2 producing P. aeruginosa isolates was determined using Enterobacterial Repetitive Intergenic Consensus-based PCR (ERIC-PCR). 31 Cycling conditions were in concordance with those described by Ferreira et al., 32 with annealing performed at 51°C for A. baumannii and at 58°C for P. aeruginosa. The amplified products were visualized on 1.5% agarose gel stained with 0.5 μg/ml of ethidium bromide. Banding patterns were converted by GelQuest/Sequentix® software into a binary matrix, calculated using Dice coefficient. A dendrogram was constructed via UPGMAs. Only visible bands in the ERIC-PCR fingerprinting were used to construct the similarity matrix and the dendrogram. Isolates with more than 85% similarity were considered to be clonally related.
Results
A total of 12,045 isolates of Enterobacteriaceae, A. baumannii, P. aeruginosa, and Pseudomonas species were screened by the participating hospitals during 2012, of which 103 out of 9485 (1.2%) Enterobacteriaceae, 638 out of 724 (88%) A. baumannii, and 760 out of 1848 (41%) Pseudomonas were nonsusceptible to carbapenems (Tables 1 and 2). Of these, a total of 398 nonrepetitive carbapenem-resistant isolates of Enterobacteriaceae, A. baumannii, and Pseudomonas that fit the inclusion criteria were collected during the study period by participating hospitals and sent to our central laboratory for analysis. Among the studied 398 isolates, 44 were Enterobacteriaceae, 142 were A. baumannii, 186 were P. aeruginosa, and 26 were Pseudomonas species. The isolates were obtained from various clinical specimens including pus, urine, stool, sputum, bronchial aspirates, blood, wound swabs, pleural fluid, and hepatic drainage.
MICERT, minimum inhibitory concentration of ertapenem; AUG, amoxicillin/clavulanic acid; FOX, cefoxitin; CTX, cefotaxime; FEP, cefepime; CAZ, ceftazidime; ATM, aztreonam; IMP, imipenem; S, sensitive; I, intermediate; R, resistant.
GES-11 is a carbapenemase of class A; IMP and VIM are carbapenemases of class B; OXA-23 and OXA-24 are carbapenemases of class D; ADC stands for Acinetobacter-derived cephalosporinases.
The geographical distribution of the collected isolates was as follows: 74.8% from Beirut, 11% from Bekaa, 10.3% from South Lebanon, 3.3% from Mount Lebanon, and 0.5% from North Lebanon. Accordingly, the majority of the samples were recovered from patients hospitalized in Beirut and its suburbs including Mount Lebanon, which represent the heaviest population density in the country.
The beta-lactamases and/or genes of insertion sequences identified in Enterobacteriaceae are shown in Table 1, while those in A. baumannii and Pseudomonas spp. are shown in Table 2. Carbapenemase production was detected in 55% (219/398) of total isolates; 70.4% of Enterobacteriaceae had blaOXA-48-like gene; all A. baumannii were positive for blaOXA-51 and had either blaOXA-23 (97.8%) or blaOXA-24 (1.4%), and/or blaGES (91.5%) genes. Twenty-one percent of P. aeruginosa and 27% of Pseudomonas spp. had either blaIMP or blaVIM metallo-beta-lactamase genes.
Twenty-percent of Enterobacteriaceae displayed an ertapenem intermediate phenotype with MICs of ertapenem being equal either to 0.5 or 1 mg/L, while 80% revealed higher MICs (MICERT >1 mg/L). Results of susceptibility to tested β-lactam antibiotics are shown in Table 1. Synergy between amoxicillin/clavulanic acid and third or fourth generation cephalosporins in the DDST was better visualized on cloxacillin-containing media and was observed for 70% of Enterobacteriaceae strains among which 36% were Klebsiella pneumoniae, 29% were Enterobacter cloacae, 19% were E. coli, 13% were Serratia marcescens, and 3% were Morganella morganii. These isolates were candidates for ESBL genes detection.
Synergy between amoxicillin/clavulanic acid and imipenem did not exist for any of our Enterobacteriaceae isolates, indicating the absence of ESBLs with carbapenem-hydrolyzing activity. The cloxacillin test was positive in two isolates of K. pneumoniae and two isolates of E. coli, indicating the possibility of production of an acquired AmpC cephalosporinase. The cloverleaf-like growth pattern in the modified Hodge test was positive for 31 out of the 44 Enterobacteriaceae isolates. Concerning efflux mechanism, six Enterobacteriaceae isolates, namely one K. pneumoniae, four E. cloacae, and one Serratia odorifera, all with high level ertapenem resistance, produced positive efflux pump inhibition test. A total of 70.4% of Enterobacteriaceae harbored blaOXA-48-like gene, with 90% of them also carrying insertion sequence IS1999. Sequencing of the OXA-48-like amplicons revealed that 67% (eight K. pneumoniae, six E. coli, two E. cloacae, four S. marcescens, and 1 M. morganii) had blaOXA-48, 19% (6 K. pneumoniae) had blaOXA-162, and 13% (one K. pneumoniae, two E. coli, and one E. cloacae) had blaOXA-232. All OXA-48-like producing isolates showed presence of IncL/M-type plasmids described by Poirel et al. 29 Acquired AmpC genes were detected in one K. pneumoniae harboring blaACC concurrent with blaOXA-48, another K. pneumoniae harboring blaMOX only, and two E. coli harboring blaCIT. More than 70% of the isolates had in addition to blaOXA-48-like gene, an ESBL of CTX-M group 1 or SHV type, making them nonsusceptible to third generation cephalosporins.
The XbaI-digestion PFGE patterns of the OXA-48-like producing isolates is shown in Figure 1. Clonally related strains existed among K. pneumoniae (4 out of 15), E. coli (2 out of 8), E. cloacae (2 out of 3), and S. marcescens (2 out of 4 isolates). The clonally related K. pneumoniae were isolated from different hospitals in Beirut, while the clonally related E. coli were isolated from a hospital in Beirut and another in Bekaa district. E. cloacae strains of pulsotype 1 and S. marcescens strains of pulsotype 1 were isolated respectively from the same hospital. Interestingly, K. pneumoniae of pulsotypes 5 and 7 produced OXA-162 while E. coli of pulsotype 7, E. cloacae of pulsotype 1, and S. marcescens of pulsotype 1 produced OXA-48.
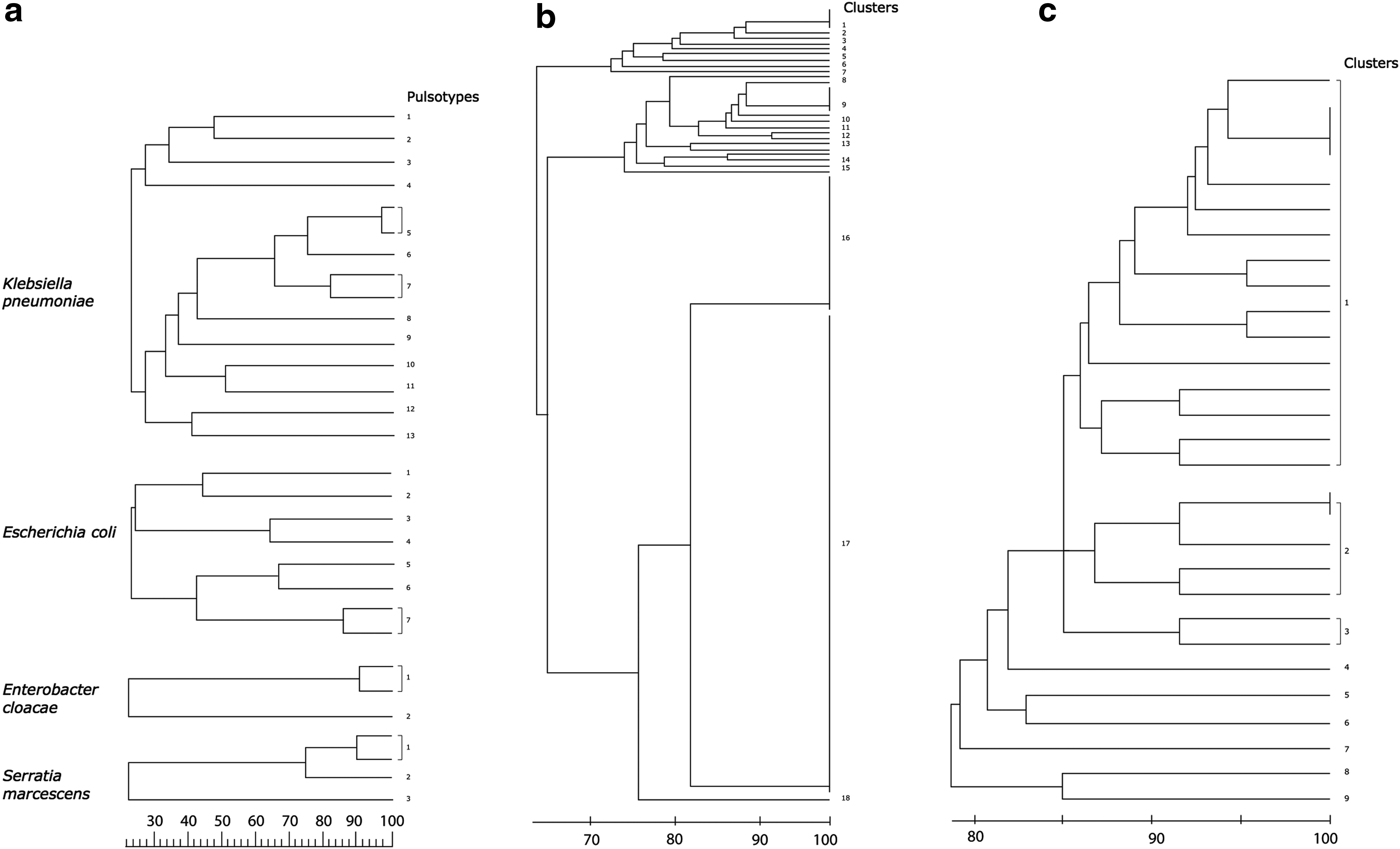
DNA fingerprinting analysis.
All 142 A. baumannii isolates were resistant to all tested antibiotics. Sixty-eight isolates yielded positive cloxacillin test indicating phenotypic evidence of hyperproduction of their ADC. Sixty-one isolates showed synergy between clavulanic acid and imipenem, indicating production of a class A ESBL with carbapenemase activity.
Genotyping of A. baumannii allowed the detection of blaOXA-51 gene in all strains, confirming species identification. GES-type carbapenemase concomitant with OXA-23 enzyme was observed in the majority of the strains (128 strains or 90%), indicating that these two carbapenemases were the most prevalent mechanisms of resistance among our isolates. Sequencing experiments of the blaGES variant revealed GES-11. Eleven strains harbored either OXA-23 alone, or OXA-24 alone, or OXA-24 with GES-11. In one strain, GES-11 was the only detected enzyme with carbapenemase-hydrolyzing capacity.
The insertion sequence ISAba1 was identified in all isolates, and mapping of this element in OXA-23-poducers revealed a band of 1371 bp, indicating that ISAba1 lies upstream and adjacent to blaOXA-23 in all strains. Moreover, mapping of ISAba1 to blaADC revealed a band of 693 bp, showing that IsAba1 existed upstream and adjacent to the ADC in 97% of the isolates. Mapping of ISAba1 to blaOXA-51 was negative.
Fingerprinting by ERIC-PCR (Fig. 1) showed the existence of 18 clusters of imipenem-resistant A. baumannii. Cluster 17 included the majority of strains (61%, 87/142) and represented 7 hospitals in Beirut, Mount Lebanon, South Lebanon, and Bekaa. Interestingly, different carbapenemase profiles were noticeable among the related isolates. For instance, strains harboring blaOXA-23 alone or blaGES-11 alone existed in cluster 17 together with those co-harboring blaOXA-23 and blaGES-11. The remaining strains, not within cluster 17, showed various fingerprinting patterns, with fewer than 10 strains in each cluster, indicating different clones.
Like A. baumannii, Pseudomonas isolates were resistant to all tested antibacterial agents with the exception of colistin. Evidence of chromosomal AmpC overexpression was detected in the cloxacillin test. Detection of metallo-β-lactamases by the Etest method gave negative results for all Enterobacteriaceae and A. baumannii isolates, but was positive for 22% (39 out of 186) P. aeruginosa and 27% (7 out of 26) Pseudomonas spp. PCR and DNA sequencing identified the presence of the metallo-β-lactamases VIM-2 gene in 16% of P. aeruginosa and in 19% of Pseudomonas spp. Also, 2% of P. aeruginosa harbored blaIMP-1, while 4% of P. aeruginosa and 8% of Pseudomonas sp harbored blaIMP-2.
Because the majority of carbapenemase producers among P. aeruginosa harbored VIM-2, clonal relatedness for these isolates was investigated. Fingerprinting by ERIC-PCR for the 29 VIM-2 positive P. aeruginosa strains showed the existence of nine clusters (Fig. 1). Sixteen strains belonged to cluster 1, and these strains originated from two hospitals in Beirut and one in South Lebanon. Clusters 2 and 3 included 5 and 2 strains respectively. The six remaining strains belonged to separate clusters (4–9).
Discussion
Despite the substantial rise of carbapenem resistance among Gram-negative pathogens in the last two decades, data concerning Lebanon remained scarce and limited to case reports or studies involving particular medical centers,33–37 stressing the need for a consolidated study. We report here the first nationwide, cross-sectional surveillance on carbapenem unsusceptibility among Gram-negative pathogens in tertiary care centers in Lebanon, involving both epidemiology of these strains and molecular basis of carbapenem resistance. The three arms of this study, directed toward Enterobacteriaceae, A. baumannii, and Pseudomonas, highlight the current status of carbapenem resistance in these groups of pathogens, and represent baseline data for further investigation on this issue.
According to our findings, a higher prevalence of carbapenem resistance exists among Pseudomonas and Acinetobacter compared to Enterobacteriaceae clinical isolates. Indeed, 1.2% of Enterobacteriaceae were nonsusceptible to carbapenems. The prominent mediator of carbapenem resistance was blaOXA-48-like gene in this study and it was found on IncL/M-type plasmids, common vehicles for blaOXA-48 described worldwide. 38 The detection of blaOXA-48-like gene among K. pneumoniae and E. coli adjoins other data reported in Lebanon.33,39 Since the first description of OXA-48 producing K. pneumoniae in Turkey, 40 numerous reports indicated its dissemination to Mediterranean countries like France and Egypt, 41 and fewer reports exist from countries in the Americas. 42 The blaIMP-1 and blaNDM-1 genes had been identified too in previous case studies in Lebanon.34,33 However, although KPC is endemic now in neighboring countries like Israel, 43 it was not detected in our samples. This indicated that Lebanese hospitals encounter a spread of Ambler class D OXA-48-like carbapenemase among Enterobacteriaceae, while spread of classes A and B carbapenemases appears to be still restricted.
In addition to the production of OXA-48-like enzyme, known to confer a low level resistance to ertapenem, 38 the studied Enterobacteriaceae associated with high MICERT levels expressed other resistance mechanisms including efflux pumps expression, ESBLs production, and plasmid AmpC cephalosporinase acquisition. We report for the first time in Lebanon the presence in Enterobacteriaceae of plasmid-borne AmpC enzymes of the classes CIT, ACC, and MOX, either alone or in combination with or OXA-48-like enzyme or an ESBL, specifically of CTX-M group 1. When combined with the loss of a major outer membrane protein or efflux pumps, AmpC cephalosporinases can contribute to increased level of carbapenem resistance. 5
From an epidemiologic perspective, analysis of the clonality of OXA-48-like-producing isolates by PFGE showed both, clonal and horizontal spread. Clonal dissemination played a role in spread of 4 OXA-162 producing K. pneumoniae and 2 OXA-48 producing E. coli strains that revealed high genetic relatedness, whereas horizontal spread accounted for the presence of unrelated strains. OXA-48 producing E. cloacae and S. marcescens strains demonstrating genetic relatedness originated from the same hospitals, suggesting that small hospital outbreaks contributed to the dissemination of the strains.
Concerning A. baumannii, 88% of isolated strains were nonsusceptible to carbapenems and represented a challenge for treatment due to significant compromise of effective antibiotics. 11 In this study, blaOXA-23 and blaGES-11 coexisted in the majority of the analyzed A. baumannii strains, confirming the predominance of these carbapenemase genes among Lebanese isolates. The dissemination of OXA-23 in Lebanon agrees with the described worldwide epidemiology of OXA-23. 44 This carbapenemase has been previously reported in countries in regions like Egypt, Saudi Arabia, United Arab Emirates, Kuwait, and Iran45–47 ; these countries may have patient or population exchange with Lebanon. Recently, this carbapenemase was described in livestock from Lebanon, 48 indicating also transfer of A. baumannii resistance genes to the community through the food chain. Our study is the first Lebanese report of OXA-24 gene, but did not detect OXA-58 described previously in Lebanon, 35 nor NDM-1 described in Syrian patients of civil war presenting to a hospital in North Lebanon. 49 The insertion sequence ISAba1 of A. baumannii was located adjacent to blaOXA-23 and to blaADC, and promoted the overexpression of these genes, consequently decreasing the susceptibility to β-lactams. 11
Genetic fingerprinting of A. baumannii strains revealed a major cluster to which more than 60% of the isolates belonged, signifying clonal dissemination as a major contributor to spread of carbapenem resistance for this organism. Remaining strains belonged to diversified clusters, suggesting that horizontal transmission plays a role, although slighter, in the dissemination of carbapenemases among A. baumannii in Lebanon. Some isolates with blaOXA-23 alone or blaGES-11 alone exhibited clonal relatedness to those co-harboring blaOXA-23 and blaGES-11, revealing that the genetic support for these two genes may not be the same. This has been previously suggested in a study from Tunisia, where blaOXA-23 harbored both chromosomal and plasmid locations, while blaGES-11 was only plasmid-borne. 50
The prevalence of imipenem resistance among P. aeruginosa in 2012 was equal to 43%. A previous Lebanese study has identified P. aeruginosa resistance to imipenem in a university medical center to range from 14% to 25% over a decade (2000 till 2011). 51 Reports describing prevalence of imipenem-resistant P. aeruginosa in our region indicate variable rates, from 16% in Israel 52 to 30% in Turkey. 53 While P. aeruginosa continues to be a predominant cause of infections with intrinsic resistance to antibiotics, such rates of carbapenem resistance in Lebanon and other countries are of imperative clinical concern. Carbapenem resistance in this organism can involve a combination of mechanisms, such as energy-dependent multidrug efflux pumps present on the cytoplasmic membrane like MexAB-OprM, and unusually restricted outer membrane permeability through mutation or changes in the porin protein OprD structure, sometimes with derepressed chromosomal AmpC β-lactamases. 14 Nevertheless, increase in carbapenem-resistant strains mediated by acquired metallo-β-lactamases including IMP and VIM in Pseudomonas have been observed worldwide. The metallo-β-lactamase IMP-1, detected in 2 of our P. aeruginosa isolates, was previously described in another Lebanese survey in K. pneumoniae. 34 This study reports, in addition, production of IMP-2 and VIM-2 carbapenemases by Pseudomonas. The finding of blaIMP-1, blaIMP-2, and blaVIM-2 metallo-β-lactamase genes in Pseudomonas strains in Lebanon is concurrent with other studies in our region, namely from Turkey and Saudi Arabia. 54 Like for A. baumannii, clonal linkage examined via ERIC-PCR revealed that mostly clonal dissemination and to a lesser extent horizontal spread of blaVIM-2 were responsible for the existence of this carbapenemase among species of Pseudomonas isolated from various hospitals.
The originality of this study lies primarily in the inclusion of isolates from variable geographic zones, revealing a nationwide view of carbapenem resistance in our country. In addition, phenotypic and genotypic analyses were performed, and ertapenem was used to increase the sensitivity in the detection of carbapenem-resistant Enterobacteriaceae. For genotyping purposes, in Acincetobacter and Pseudomonas, we used ERIC-PCR that is a rapid and inexpensive method, practical in limited resource settings. A larger sample contribution from participating hospitals would have been more efficient to analyze clonal relationships.
Looking back into clinical data available from patient profiles at Beirut and Mount Lebanon hospitals showed that some patients were from other countries like Syria, Iraq, or France. Nevertheless, the majority of the patients were Lebanese with underlying malignancy, immune-suppression, severe trauma, or chronic complications of diseases like diabetes, heart disease, or respiratory insufficiency. Some also suffered from nosocomial acquisition of resistant pathogens due to long hospital stays or procedures like catheterization. Few patients were transferred from remote hospitals to others in Beirut to manage critical cases. Such data suggest that carbapenemases might have been present in the country from a long time, and warrant the need for prolonged surveillance studies that may better represent the status of carbapenemase dissemination. In addition, patient transfers from neighboring countries eventually increases the spread of carbapenemase-producers in Lebanon, especially with the demographic changes and public health fluctuations caused by civil war and political instability in the region. This, in turn, raises the need for infection control practices to contain the dissemination of these strains before they become alarming.
In conclusion, carbapenem-nonsusceptible Enterobacteriaceae are slightly diffusing in Lebanese hospitals and resistance depends predominantly on OXA-48 production. However, carbapenemase-producing A. baumannii and Pseudomonas are highly disseminating with the predominance of OXA-23 and GES-11 in A. baumannii and the occurrence of metallo-β-lactamases IMP and VIM, in Pseudomonas clinical isolates. Responding to the challenges of Gram-negative resistance will require a multifaceted approach including advanced and rapid methodology of detection, improved surveillance, adherence to basic measures of infection control, education, and ongoing research on resistance mechanisms and epidemiology.
Footnotes
Acknowledgment
The authors would like to acknowledge Charbel Akkawi for technical assistance in the experimental work.
Disclosure Statement
Authors have no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.