
Abstract
Non-O157 Shiga toxin-producing Escherichia coli (STECs) are not as well characterized as O157 STEC cases, despite their similar prevalence in many countries. Hence, the objective of this study was to investigate the phenotypic and genotypic basis of multidrug resistance (MDR) in non-O157 STEC farm- and abattoir-sourced isolates and assess the potential dissemination of these MDR profiles in vitro. Susceptibility testing to 20 antimicrobials was performed on 146 non-O157 STECs isolated from farm and abattoir environments. Eighty-seven percent of non-O157 STEC isolates were multidrug resistant to antimicrobials used during veterinary and agricultural practice. Antimicrobial resistance was significantly higher in abattoir isolates compared with the farm isolates (p < 0.05). Corresponding resistance determinants and integrons were investigated by polymerase chain reaction, with the predominant resistance determinants detected being floR, ampC, tet(A), blaTEM, and sul1. This is the first report of tet(G) in a non-O157 STEC isolate. Class 1 integrons were detected in 17 isolates. Resistance to ampicillin, cephalothin, chloramphenicol, kanamycin, neomycin, sulfonamides, trimethoprim, and tetracycline was associated with transferable plasmids belonging to incompatibility groups IncP, IncB/O, and IncFIB. Most MDR non-O157 STECs (90%) isolated in this study belong to phylogenetic groups A and B1. These findings suggest that MDR non-O157 STECs are emerging as a result of nonpathogenic E. coli acquiring virulence and resistance genes. This may convey a certain competitive advantage in the colonization of cattle when antimicrobial selective pressures are present, thereby leading to an increase in contamination of food with MDR non-O157 STECs.
Introduction
S
Half of all confirmed human STEC cases in Europe are caused by non-O157 STECs, 5 and in the United States, non-O157 STEC infections outnumber O157 cases.1,6 The lack of convenient, high-throughput, and economical methods capable of distinguishing STECs from other E. coli has resulted in an information gap with regard to the emerging non-O157 serogroup and consequently their potential public health significance.1,7
Although antimicrobial therapy is not the means of treating STEC infections, multidrug-resistant (MDR) STECs are a public health concern as these strains contribute to the reservoir of resistance genes that can then be easily exchanged between Enterobacteriaceae in the host and in the environment. Both human and animal-derived non-O157 STEC strains have demonstrated multiple resistances to commonly applied antimicrobial compounds used in the animal production industry.8,9 MDR in non-O157 STECs could convey a competitive advantage to these strains, resulting in their increased prevalence in production animals and ultimately in the food products derived from them. 10 It is widely accepted that the extensive use of antimicrobial agents in animal production acts as an important driver for the development of MDR bacteria. 11 It is established that pre-exposure of bacteria to subtherapeutic doses of antibiotics promotes transient adaptive resistance to lethal levels of these antibiotics and potentially to other unrelated antibiotics.11–14 Long-term subtherapeutic exposure to antibiotics can result in the enrichment of mutations and/or the acquisition of mobile genetic elements that can then confer a reduced susceptibility phenotype to these compounds.11,13–15
Several mechanisms of horizontal gene transfer (HGT) exist, which enable bacteria to acquire antibiotic resistance genes, including but not limited to plasmids, transposons, and integrons.16–18 Since subinhibitory concentrations of antibiotics can also enhance the HGT of antibiotic resistance-encoding genes between closely related and unrelated bacteria, this raises concerns regarding the dissemination of antimicrobial resistance through bacterial populations in ecological niches connected to animal and human health.19–22 HGT has been experimentally detected at several stages in the food production process, including the farm, the abattoir, and during processing on various food matrices.8,16,23–25 The presence of antimicrobial-resistant bacteria in food, regardless of pathogenicity, constitutes a risk to public health as the pool of genes from which pathogenic bacteria can acquire antibiotic resistance is increased. 13
Integrons are considered the main mechanism driving the dissemination of antibiotic-resistant genes among gram-negative Enterobacteriaceae such as STECs.8,26,27 Subtherapeutic drug doses enhance the rate of conjugative plasmid transfer. 19 This development in turn induces the bacterial SOS response, upregulating the expression of int1, a gene located in the 5′conserved segment (5′CS) of their structure and encoding a site-specific integrase, resulting in the capture or exchange of one or more resistance-encoding genes in gram-negative pathogens such as STEC. 22
Given that Ireland has the highest incidence of STEC infections per capita (8.99 per 100,000) in Europe, 5 the widespread presence of MDR STEC bacteria within the food chain is a significant public health concern.13,28 As part of a larger study, farm and abattoir environments were screened for non-O157 STEC isolates, as published previously by Monaghan et al.,7,29 whereby culturable non-O157 STEC isolates (defined as the presence of stx1 and/or stx2 and their variants) were recovered at a frequency of 8% and 3% in fecal and soil samples, respectively, and at 6% and 1% in hide and carcass samples, respectively. To further assess the potential public health impact of these isolates, the aim of this study was to investigate the phenotypic and genotypic bases of MDR in these farm- and abattoir-sourced isolates and assess the potential dissemination of these MDR profiles in vitro.
Materials and Methods
Selection of bacterial isolates for this study and resistance profiling
Previously, isolates were selected from farms (sampled bimonthly from July 2007 to July 2008; Monaghan et al. 7 ) and abattoirs (sampled bimonthly from July 2009 to July 2010; Monaghan et al. 29 ) based on their non-O157 STEC serogroup and virulence gene profiles (farm n = 106 and abattoir n = 40). Susceptibility testing by disc diffusion was performed, and results were interpreted as recommended by Clinical and Laboratory Standards Institute guidelines. 34 The following antimicrobial compounds were used: amikacin (AK), 30 μg; amoxicillin–clavulanic acid (AMC), 20/10 μg; ampicillin (AMP), 10 μg; cefpirome (CPO), 30 μg; cefpodoxime (CPD), 10 μg; ceftiofur (EFT), 30 μg; cephalothin (KF), 30 μg; chloramphenicol (C), 30 μg; ciprofloxacin (CIP), 5 μg; colistin (CT), 25 μg; florfenicol (FFC), 30 μg; furazolidone (SAM), 15 μg; gentamicin (CN), 10 μg; kanamycin (K), 30 μg; nalidixic acid (NA), 30 μg; neomycin (N), 30 μg; sulfonamide (S3), 300 μg; trimethoprim–sulfamethoxazole (SXT), 1.25/23.75 μg; tetracycline (TE), 30 μg; and trimethoprim (W), 5 μg. Antibiotic-containing discs were supplied by Oxoid (Basingstoke, United Kingdom), and E. coli ATCC 25922 was included as a quality control strain.
MDR non-O157 STEC isolates representative of all resistance profiles in farm and abattoir environments (n = 90) were selected for further genotypic analysis. MDR is defined as resistance to three or more different classes of antibiotics. 31
Phylogenetic grouping
Phylogenetic groups were determined for each non-O157 STEC isolate (n = 90) using an established multiplex polymerase chain reaction (PCR) targeting chuA, yjaA, and TSPE4.7 according to the protocol of Clermont et al. 32 This method classifies E. coli into four phylogenetic groups designated A, B1, B2, and D. Target amplification was performed using the original primer concentrations and cycling conditions. Amplicons generated were separated by conventional 1.7% (wt/vol) agarose gel electrophoresis, stained with 0.1 g/ml ethidium bromide (Sigma-Aldrich, Steinheim, Germany) in 0.5× Tris-EDTA-boric acid buffer, and subsequently assigned to one of the lineages (A, B1, B2, or D) using the criteria outlined previously by Clermont et al. 32
Detection of antibiotic resistance genes and class 1 and class 2 integrons
Total genomic DNA was extracted using a Promega Wizard Genomic DNA purification kit (Promega, Madison, WI) following the manufacturer's protocol. The DNA concentration was measured using a Nanodrop ND-1000 spectrophotometer (Thermoscientific, Wilmington, DE). Detection of antibiotic resistance markers and integron-associated genes was performed using the primers listed in Table 1. The following resistance determinants were investigated by PCR: ampC, blaOXA, blaSHV, blaTEM, and blaCTX-M, which encode β-lactamases; chloramphenicol (cmlA1) and chloramphenicol/florfenicol (floR) efflux pumps; sulfonamide resistance gene (sul1); and the tet(A), tet(E), and tet(G) tetracycline efflux pumps. Previously published PCR methods were employed to determine the presence of conserved integron-associated genes, including int1 and int2 (coding for integrases of classes 1 and 2, respectively), qacEΔ1, sul1, and the variable regions of class 1 and 2 integrons (Table 1). All PCRs were performed in a final volume of 25 μl consisting of 2.5 μl 10× PCR buffer (New England BioLabs, Ipswich, MA), 25 pmol of each primer (MWG-Biotech AG, Ebersberg, Germany), deoxynucleoside triphosphates at a concentration of 200 μM (Promega), 1 U Taq DNA polymerase (New England BioLabs), and 50 ng of genomic DNA.
F, forward; R, reverse.
Conjugation experiments
Conjugation mating experiments were performed with the plasmid-free recipient strain E. coli 26R793 (Rifr, lac negative) to assess the transfer of the associated resistance markers. Seventeen integrin-positive non-O157 MDR STEC isolates from the collection were selected as donor strains. Briefly, single colonies of the donor and recipient were inoculated in Luria-Bertani (LB) broth (Oxoid) and grown overnight at 37°C. Donor and recipient cultures were mixed at a ratio of 1:4, gently centrifuged, and resuspended in 30 μl cold saline buffer. In each case, the mating mixture was spotted onto a dry, prewarmed LB agar plate (Sigma-Aldrich) and incubated for 6 hours at both 37°C and room temperature. After incubation, the mating mixtures were resuspended in cold saline buffer and spread onto MacConkey agar plates (Sigma-Aldrich) containing 100 g/ml rifampicin (Sigma-Aldrich) and 50 μg/ml ampicillin, 20 μg/ml chloramphenicol, 30 μg/ml tetracycline, or 50 μg/ml trimethoprim (Sigma-Aldrich). Three colonies from each selection plate were examined and phenotypically confirmed to have newly acquired antimicrobial resistance by disc diffusion tests as described previously.
Plasmid incompatibility typing
Plasmid DNA was extracted from confirmed transconjugants using QuickGene plasmid kit S (Fuji, Tokyo, Japan) according to the manufacturer's instructions. Plasmids were assigned to incompatibility groups (Inc) on the basis of the presence of specific replicon sequences identified by PCR according to the method developed by Carattoli et al. 33 PCR was performed using 18 pairs of primers recognizing the FIA, FIB, FIC, HI1, HI2, I1-Iy, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and FIIA replicons representative of the major incompatibility groups circulating among the Enterobacteriaceae.
Statistical analysis
Data were analyzed with SSPS version 16.0 software (SPSS, INC., Chicago, IL). For univariate analysis, the χ2 or Fisher's exact test was used as appropriate. A p-value of <0.05 was considered significant.
Results
MDR profiles
The distribution of resistance of all non-O157 STECs isolated in this study was as follows: sulfonamide (S3) 99%; tetracycline (TE) 82%; colistin (CT) 59%; ampicillin (AMP) 53%; cephalothin (KF) 31%; neomycin (N) 26%; kanamycin (K) 21%; trimethoprim (W) 17%; trimethoprim–sulfamethoxazole (SXT) 16%; ceftiofur (EFT) 16%; furazolidone (SAM) 11%; cefpodoxime (CPD) 6%; chloramphenicol (C) 5%; amikacin (AK) 4%; gentamicin (CN) 2%; florfenicol (FFC) 1%; amoxicillin–clavulanic acid (AMC) 0%; cefpirome (CPO) 0%; ciprofloxacin (CIP) 0%; and nalidixic acid (NA) 0%. The distribution of resistance in the farm and abattoir environments is presented in Fig. 1. Resistance to AK, CPD, KF, CT, N, SXT, and W occurred more frequently among the abattoir isolates compared with the farm isolates. Resistance to AMP was more prevalent among farm isolates.
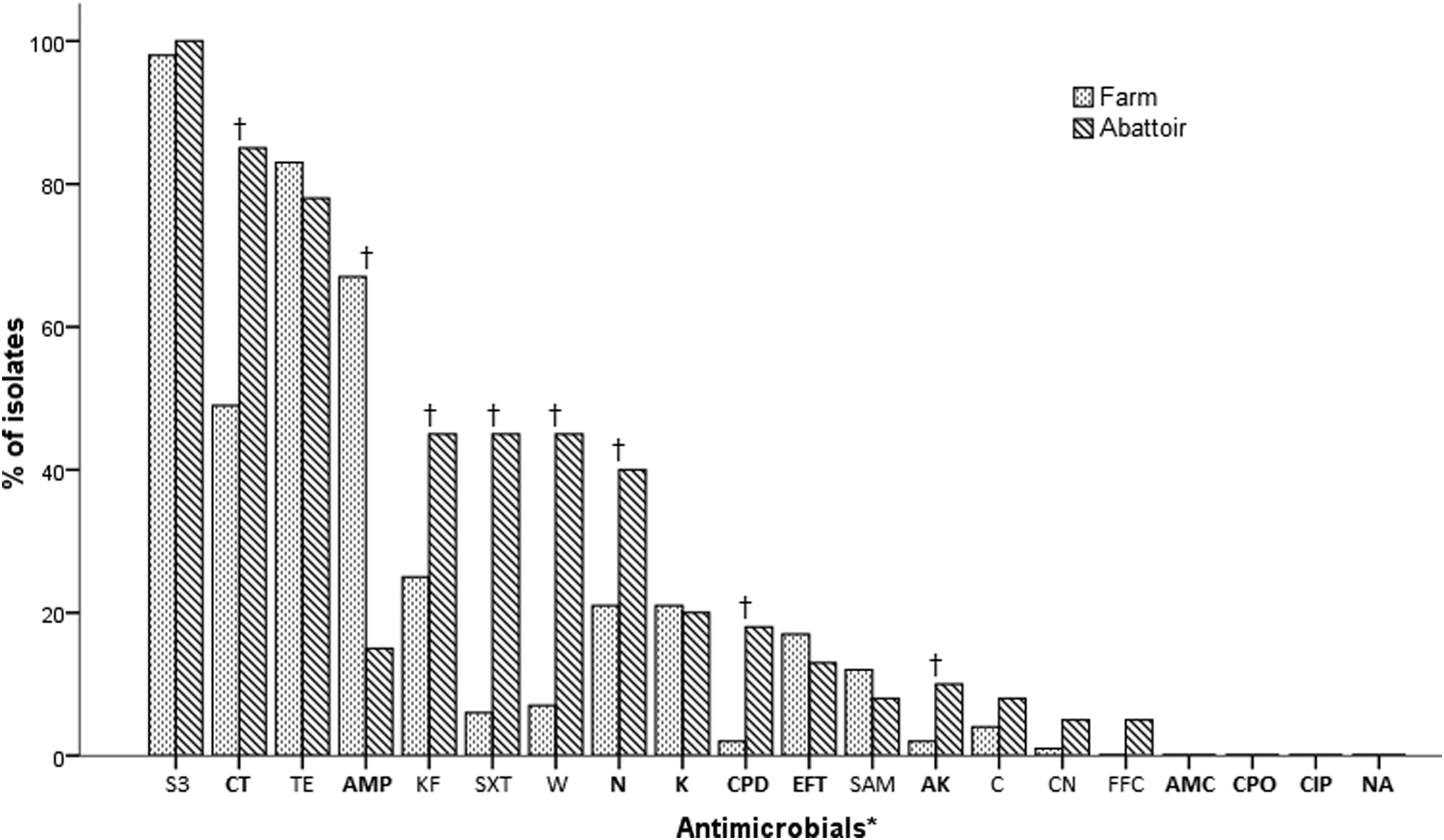
Prevalence of antimicrobial resistance among farm and abattoir non-O157 STEC isolates. The panel of antimicrobials tested consisted of *
The majority of the non-O157 STEC isolates (82%) demonstrated MDR. MDR isolates were resistant to between 3 and 10 antimicrobial compounds with 64 different resistance profiles recognized. Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr) in the Supplementary Data presents these results in detail. Of the MDR isolates, the most frequent resistance profile identified was CT/S3/TE, present in 14% of isolates, followed by AMP/S3/TE (4%), AMP/CT/S3/TE (4%), AMP/CT/KF/S3/TE (3%), CPD/CT/KF/N/S3/SXT/TE/W (3%), AMP/K/N/S3/TE (3%), AMP/CT/EFT/S3 (3%), CT/KF/N/S3/SXT/TE/W (2%), AMP/K/KF/N/S3/TE (2%), and AMP/CT/S3 (2%). The three most common MDR profiles among farm isolates had resistance to up to four antimicrobials, AMP/S3/TE (15%), CT/S3/TE (9%), and AMP/CT/S3/TE (6%). The most common MDR profiles among abattoir isolates had resistance to between three and eight antimicrobials. CT/S3/TE (25%), CPD/CT/KF/N/S3/SXT/TE/W (10%), and CT/KF/N/S3/SXT/TE/W (8%).
Phylogenetic groups
Seven percent of the isolates belonged to the phylogenetic group B2 associated with virulent extraintestinal strains. Three percent of the isolates were assigned to phylogenetic group D, which is associated with pathogenic bacteria, although less frequently than group B2. Sixty-nine percent and 21% of the non-O157 STEC isolates belonged to the phylogenetic groups B1 and A, respectively. 32 All, but one, virulence-associated isolate originated from the farm environment.
Frequencies of resistance markers
A summary of the frequency of resistance markers identified in all of the farm and abattoir isolates is presented in Table 2. Supplementary Table S1 in the Supplementary Data details the resistance genotypes from the isolates in the collection.
Only two of the five β-Lactam-resistant genes screened were detected in this study, in 65% of β-lactam-resistant isolates, with the ampC gene most prevalent (39%), followed by blaTEM (34%). A total of 16 isolates contained the ampC and blaTEM resistance markers simultaneously.
Six chloramphenicol-resistant isolates were detected, with two isolates carrying the floR gene encoding the chloramphenicol/florfenicol efflux pump. Both isolates conferring resistance to florfenicol also carried the floR gene, one of which was coresistant to chloramphenicol. A further seven isolates possessed the floR gene without exhibiting resistance to either drug.
The tetracycline resistance marker tet(A) was identified in 18% of tetracycline-resistant isolates, while tet(G) was identified in one isolate. Interestingly, two tetracycline-susceptible isolates carried the tet(A)gene.
sul1 was identified in 31% of the sulfonamide-resistant isolates.
Integrons and gene cassettes
The genetic markers associated with class 1 integrons were identified in 21% of isolates, with int1, qacEΔ1, and sul1 detected in 18%, 17%, and 20% isolates, respectively (Supplementary Table S1). Fifteen of these isolates were positive for all three markers, although four did not contain a class 1 gene cassette. One of the int1-positive isolates did not carry the sul1 gene and did not contain a variable gene cassette. Gene cassettes carried by the int1-positive isolates had the following sizes; 800 bp (n = 7), 1 kb (n = 3), and 2.7 kb (n = 1). A further three int1-negative isolates were positive for a variable gene cassette, one of which also contained the sul1 gene. No isolates carried class 2 integrons, as determined by the absence of an integrase (int2) amplicon.
Conjugal transfer of resistance
Resistance transfer by conjugation was observed for 8 of the 17 integron-associated MDR isolates that were subjected to conjugation assays (Table 3). In all cases, ampicillin, tetracycline, and trimethoprim resistance could be transferred to the recipient, while chloramphenicol resistance transfer only occurred in one isolate. Two of the donor strains transferred resistance to ampicillin, kanamycin, neomycin, sulfonamide, trimethoprim–sulfamethoxazole, tetracycline, and trimethoprim (AMP/K/N/S3/SXT/TE/W). Another pair of isolates transferred resistance to neomycin, sulfonamide, trimethoprim–sulfamethoxazole, tetracycline, and trimethoprim (N/S3/SXT/TE/W). One isolate (A77) could transfer its entire MDR resistance repertoire (K/KF/N/S3/SXT/TE/W).
Plasmid incompatibility
Plasmids present in the transconjugants were assigned to the following incompatibility groups; IncFIB, IncA/C, IncP, IncF, and IncB/O. The characteristics of the donors and their corresponding transconjugants are listed in Table 3.
Discussion
Non-O157 STEC strains are increasingly recognized as important food-borne pathogens worldwide, with the number of non-O157 STEC cases now equaling if not exceeding the number of O157 STEC cases in many countries.5,6 In this study, the majority (87%) of non-O157 STECs isolated from farm and abattoir environments were classified as being MDR, expressing resistance to three or more antimicrobials commonly used during veterinary and agricultural practice. 31 Resistance to antimicrobials was at a higher frequency in the abattoir isolates compared with the farm, highlighting the abattoir as a critical point at which strategies aimed at reducing the overall MDR non-O157 STEC bacterial contamination in the food chain would be most effective.
Generally, E. coli belonging to phylogenetic groups B2 and D are considered more likely to be pathogenic, while groups A and B1 are considered nonpathogenic commensal strains. 32 Interestingly, most of the MDR non-O157 STECs (90%) isolated in this study belong to the nonvirulence-associated phylogenetic groups. This is supported by findings from other studies showing that non-O157 STEC isolates from healthy cattle belong predominantly to lineages A and B1. 34 This suggests that MDR non-O157 STECs are emerging as a result of nonpathogenic E. coli acquiring virulence and resistance genes from bacteria in the same ecological niche. This acquisition may convey a certain competitive advantage allowing non-O157 STECs to preferentially colonize cattle over other enteric bacteria when antimicrobial selective pressures are present, thereby leading to an increased prevalence.10,11,13,15 This could ultimately result in greater contamination of food with MDR STECs. In 2011, the USDA Food Safety and Inspection Service declared six STECs (O26, O45, O103, O111, O121, and O145) as adulterants in raw, nonintact ground beef products. 35 Of the isolates belonging to the virulence-associated phylogenetic groups in this study, four belonged to the human clinically important O2:H27 and O119:H5 serotypes.2,29 Only one of the pathogenic isolates was from the abattoir, which is encouraging since the majority of virulent strains were not isolated from the processing chain. Less encouraging is the fact that all, but one virulence-associated isolate, were MDR, suggesting evidence of coselection for antibiotic resistance and virulence. Further refinement of this phylogenetic classification has recently been documented by Clermont et al. 36
Resistance to antimicrobials was at a higher frequency in the abattoir isolates compared with the farm isolates. The most prevalent abattoir STEC MDR profiles demonstrated resistance to up to eight antimicrobials, twice as many as the most prevalent farm STEC MDR profiles. It is tempting to speculate that increased fecal shedding due to stress induced by animal transportation, long wait times in the lairage, higher throughput of animals in the abattoir, and the presence of a greater diversity of animals from different farm origins could have contributed to this. This could also explain the higher frequency of resistance and greater diversity of non-O157 MDR STECs in the abattoir compared with the farm. More importantly, this study highlights the abattoir as an important point at which strategies which could both detect and subsequently reduce the overall MDR non-O157 STEC bacterial populations would be most effective. There is a correlation between feces and carcass/hide contamination that is a result of cross-contamination between animals and cross-contamination between hide and carcass at the abattoir.37–39
The frequency of resistance to newer, more clinically important antimicrobial compounds, such as fluoroquinolones and cephalosporins, was low in this study. Production of β-lactamase is considered the main mechanism of resistance to cephalosporins (β-lactams) in gram-negative bacteria. 40 Extended-spectrum cephalosporins are an important class of drug used in human and veterinary medicine. In this study, various types of narrow and extended-spectrum β-lactamase-encoding genes were investigated. Only two were identified, ampC and blaTEM. Narrow-spectrum TEM-1 is common in non-O157 STECs in animals, and consequently, the high rates of ampicillin-resistant blaTEM-positive isolates were to be expected. 26 The results of this study are similar to a previous report by Scott et al., 15 where blaTEM was identified in non-O157 STEC isolated from cattle feces and soil samples, but none of the other blaOXA, blaSHV, and blaCTX-M variants could be detected. Poultry-associated STEC is more likely to carry the blaSHV variant, 41 and blaOXA, blaSHV, and blaCTX-M have been previously reported in STEC O157:H7 serotypes. 42
The high levels of resistance to sulfonamides, tetracycline, and ampicillin detected in the non-O157 STEC examined are supported in the wider literature.3,26,43 In this study, the tet(A) marker was the predominant tetracycline resistance gene identified in the collection (43%), which is one of the most widespread tet genes found in Enterobacteria. However, to the best of our knowledge, this is the first report of the presence of tet(G) in an non-O157 STEC isolate.
The frequency of resistance to chloramphenicol (5%) and florfenicol (1%) was low in the non-O157 MDR STEC examined. Resistance to florfenicol, a drug closely related to chloramphenicol, is mediated by the floR gene, which encodes a florfenicol/chloramphenicol efflux pump. 44 Although florfenicol is not approved for human use, it is related to chloramphenicol and can select for cross-resistance in bacterial pathogens. In the United States, floR was present in 43% of chloramphenicol-resistant E. coli isolates from retail meats. 45 In this study, floR was identified at a higher frequency in the abattoir non-O157 STEC isolates (26%) compared with the farm (5%) further highlighting the abattoir as critical in the dissemination of antimicrobial resistance throughout the food chain.
One concerning finding in this study was that over half of all the non-O157 STEC isolates examined were resistant to the antibiotic, colistin. We consider this rate of incidence to be high given that low levels of resistance to colistin (3 of 150 E. coli beef isolates) were identified in France and Germany. 46 Our data on colistin resistance must be interpreted with caution due to the fact that colistin diffuses poorly in agar. 47 While this colistin data requires further validation, it is of concern. The lack of novel antibiotics with which to combat MDR gram-negative bacteria has led to the revival of polymyxins, such as colistin, as a last-line treatment.48–50 Since its discovery in the 1940s, colistin was used therapeutically in Japan, Europe, and the United States. It was gradually abandoned during the 1980s due to reports of nephrotoxicity and neurotoxicity.51,52 Recent studies of patients who received intravenous polymyxins for the treatment of Pseudomonas aeruginosa and Acinetobacter baumannii infections have supported the conclusion that these antibiotics have acceptable effectiveness and less toxicity than previously reported.48,53 Resistance to colistin may occur due to alterations in the bacterial cell outer membrane or an efflux pump/potassium system.48,54–56 Conjugative transfer of colistin resistance was not demonstrated in this study. However, not all plasmids transfer by conjugation. 57 The plasmid-mediated polymyxin resistance mechanisms, MCR-1 and MCR-2, have recently been characterized.58,59 Further investigation into the basis of colistin resistance should focus on the characterization of novel colistin resistance genes and their mechanism of transfer.
Class 1 integrons are thought to be one of the mechanisms responsible for the rise in MDR STECs in animal production environments.8,26,27 Integrons are gene-capture systems that play a fundamental role in the dissemination of antimicrobial resistance genes and are thought to be one of the mechanisms responsible for the rise in MDR STECs in animal production environments.8,26,27 In this study, class 1 integrons linked to 11 MDR profiles were detected in non-O157 STEC isolates from both the farm and abattoir environments. The results of this study demonstrated a similar incidence of integrons (16%) previously reported in the United States, where 18% of STECs (including O157:H7) isolated from cattle were positive for class 1 integrons. 8
The occurrence of conjugation events is feasible under the conditions present in animal farm and abattoir environments.13,60,61 Conjugative transfer of plasmids belonging to incompatibility groups IncP, IncB/O, and IncFIB was observed in this study. This is interesting given that both IncB/O and IncFIB plasmids were previously found to be relatively common among avian pathogenic E. coli (APEC) isolates as well as in both human commensal and uropathogenic E. coli (UPEC) isolates. 62 Plasmids of the IncP group are recognized to be a significant antibiotic resistance reservoir due to their self-transmissibility and their broad host range. 63 In this study, conjugation experiments produced successful transconjugates with MDR to aminoglycosides, β-lactamases, phenicols, sulfonamides, trimethoprim, and tetracycline. IncP replicons in the transconjugants were almost always accompanied by IncB/O. It is noteworthy that the plasmids identified did not belong to incompatibility groups considered epidemic, such as IncFII, IncA/C, IncL/M, or IncI1, which are ubiquitous in various geographical regions and bacterial hosts. 18
The current focus on pathogens at abattoirs in developed countries is on animal health, production loss, or major food-borne pathogens. 64 However, the development of government-led collaborative surveillance programs, which are focused around abattoir data collection, establishment of performance targets, and the public dissemination of results, is focusing industrial attention on this problem. The publication of these surveillance data may help to encourage the implementation of microbial reduction measures in abattoirs. Efforts to design, develop, and implement abattoir-based surveillance for antimicrobial resistance on a continuing basis have resulted in a number of global programs. Monitoring trends in antimicrobial resistance is now performed at various points in the food chain, including the abattoir, on a global scale. This is performed by many cooperating government and academic programs, including the Canadian Integrated Program for Antimicrobial Resistance Surveillance, National Antimicrobial Resistance Monitoring System, the Danish Integrated Antimicrobial Resistance Monitoring and Surveillance, Colombian Integrated Program for Antimicrobial Resistance Surveillance and World Health Organization Global Reports on Surveillance of Antimicrobial Resistance.64,65
In conclusion, MDR non-O157 STECs in cattle farms and abattoirs are present as a result of nonpathogenic E. coli acquiring virulence and resistance genes. The colonization of cattle with these MDR non-O157 STECs is a public health concern as they have a certain competitive advantage when antimicrobial selective pressures are present, consequently leading to an increase in contamination of food with MDR non-O157 STECs. This study further highlights the need for coordinated global action with regard to antimicrobial use in food animal production.
Footnotes
Acknowledgment
Funding for this research was provided under the National Development Plan, through the FIRM fund, administered by the Department of Agriculture, Fisheries and Food, Ireland.
Disclosure Statement
The authors of this article have no commercial associations that might create a conflict of interest in connection with the submitted article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.