
Abstract
We characterized extended-spectrum β-lactamases (ESBL) enzymes among Salmonella strains isolated in Brazil from 2009 to 2014. Salmonella recovered from both clinical and nonhuman (food, poultry, and environment) sources were subjected to antimicrobial susceptibility testing. β-lactamases genes were detected by polymerase chain reaction/sequencing; plasmid profiles and transferability were assessed by S1-pulsed field gel electrophoresis (PFGE). Genetic diversity was evaluated by XbaI-PFGE. Out of 630 Salmonella strains screened, 46 displayed ESBL phenotype, distributed across 11 different serotypes. blaCTX-M-8 and blaCTX-M-2 genes were detected at frequencies of 47% and 41%, respectively. blaSHV-5 and blaSHV-2 were also detected but in lower frequencies (4%, 2%). blaTEM-1 gene was detected in 22% of the strains. Most of the ESBL genes were transferable by conjugation, and the respective blaESBL gene was detected in the recipient strain, indicating the location of ESBL determinants on transferable plasmids. XbaI-PFGE revealed genomic diversity of Salmonella Typhimurium bearing blaCTX-M-2, blaCTX-M-8, blaTEM-1, and blaSHV-2 genes. Salmonella Muenchen (harboring blaCTX-M-2) and Salmonella Corvallis (blaCTX-M-8 and blaSHV-5) showed clonal relatedness within respective serotypes. Our findings underscore the occurrence of diverse ESBL genes in several Salmonella serotypes, reinforcing the need for continuous surveillance of resistance genes circulating in human and nonhuman sources.
Introduction
N
The resistance to expanded-spectrum cephalosporins is mainly mediated by production of acquired extended-spectrum β-lactamases (ESBL), particularly by CTX-M, TEM, and SHV enzymes, and also by production of acquired AmpC β-lactamases, 2 which often are plasmid encoded.
In recent years, ESBL-producing Salmonella enterica serotypes have emerged globally from human and food-producing animals,2,6–9 leading to strong public health concern.2,10 Previously, we have reported clonally related ESBL-producing Salmonella Typhimurium, carrying the blaCTX-M-2 gene, from clinical and nonhuman sources across different Brazilian regions. 11
To assess the current distribution of ESBL in non-typhoid Salmonella, the aims of this study were to evaluate the occurrence of ESBL-producing S. enterica serotypes, to identify the genes involved with expanded-spectrum cephalosporin resistance and to determine the genetic relatedness among isolates, both from clinical and nonhuman sources in Brazil.
Materials and Methods
Bacterial strains
From 2009 to 2014, a total of 3387 Salmonella isolates from human (n = 1296, nonduplicate isolates from stool, blood, body fluid, and urine) and nonhuman sources (n = 2091, including poultry and its environment, food-producing animals, other foodstuffs, environment samples [sludge], and unknown sources) were received for serotyping at Adolfo Lutz Institute, a reference laboratory of public health in Brazil. These isolates had been isolated and identified in regional public health laboratories, private and public hospitals, animal pathology and food microbiology laboratories from different regions of the State of São Paulo, on a voluntary basis. As part of routine surveillance, 630 of these Salmonella isolates were subjected to antimicrobial susceptibility testing: 495 from human sources (stool, 235; blood, 173; body fluids, 50, and urine, 37) and 135 from nonhuman sources [poultry samples: poultry meat (carcasses) (49), organs (12), poultry feed (08), and poultry environment (18); foodstuffs: eggs “in natura” (02), raw eggs-containing foods including homemade mayonnaise (06) and desserts (03), poultry sausage (07), pork meat (05) and sausage (06), seafood (03), raw kibbeh (02), beef burger (04), milk (01), drinking water (02), and black pepper (02); and sludge (05)].
All the human Salmonella isolates recovered from extra-intestinal sources, and representative samples, mainly associated with suspected outbreaks, of gastroenteritis infections received from diversified cities in the State of São Paulo were also submitted to AST. From the nonhuman sources, strains submitted to AST were proportionally selected based on the most prevalent contemporary serotypes identified during the study period: Salmonella Typhimurium, 37; Salmonella Enteritidis, 30; S. I 4,5,12:i:-, 23; and 16 other serotypes represented by more than one isolates (45 strains). Isolates that displayed ESBL phenotype were included in this study.
Serotyping
Salmonella serotyping was performed according to the 9th edition of White-Kauffmann-Le Minor scheme12–14 on the basis of somatic O and H flagellar antigens by agglutination tests with antisera (prepared in the Laboratory of Enteric Pathogens, Adolfo Lutz Institute, São Paulo).
Antimicrobial susceptibility test
Antimicrobial susceptibility was determined by the disk-diffusion method using Mueller Hinton agar plates (Merck, Darmstadt, Germany), according to the recommendations of the Clinical and Laboratory Standards Institute (CLSI).15,16 CLSI breakpoints 15 were employed to define the susceptible, intermediate, and resistance categories.
All the strains submitted to antimicrobial susceptibility test were also tested for ESBL phenotype, detected by double disk diffusion testing: the keyhole effect observed between any cephalosporin and amoxicillin-clavulanate disks indicated partial or total restoration of the activity of ceftazidime or cefotaxime by clavulanic acid. 17 Minimum inhibitory concentrations (MIC) were determined for ampicillin, ceftazidime, cefotaxime, ceftriaxone, and cefepime by E-test (BioMérieux, Marcy-l'Étoile, France), according to the manufacturer's recommendations. Escherichia coli ATCC 25922, E. coli ATCC 35218 and Klebsiella pneumoniae ATCC 700603 were used as the quality control strains for antimicrobial susceptibility testing.
Conjugation and transformation assays
Conjugal transfer of plasmids was carried out in mixed broth cultures with E. coli K12 strain J53 (sodium azide-resistant) as the recipient. Transconjugants were selected on MacConkey (Merck, Darmstadt, Germany) agar plates containing cefotaxime (10 mg/L; Sigma-Aldrich, St. Louis) and sodium azide (100 mg/L; Sigma-Aldrich, St. Louis).
For the strains in which transfer of plasmids encoding cefotaxime resistance was not achieved by conjugation, transformation experiments were conducted: total plasmids were extracted using commercial kits (Qiagen, Venlo, Netherlands) according to the manufacturer's instruction. Purified plasmids were transformed using Electromax (Gibco Invitrogen) and E. coli DH10B competent cells. Transformants were selected on Trypticase soy agar (Merck, Darmstadt, Germany) plates containing cefotaxime (10 mg/L; Sigma-Aldrich, St. Louis). The resistance profile of transconjugants and transformants was determined by the disk diffusion method as described above.
Plasmid analysis
Plasmid DNA was visualized from all Salmonella and E. coli transconjugant and transformant strains after linearization with S1 nuclease (Promega, Wisconsin-Madison) and electrophoresis in a CHEF-III equipment (Bio-Rad, Hercules, CA). Electrophoresis was for 17 hr with initial switch time of 1.0 sec and final switch time of 40.0 sec. Plasmid sizes were estimated with the Bionumerics v. 7.5 software (Applied Maths, Sint-Martens-Latem, Belgium) by using as references the plasmids contained in strains E. coli HB101 APr (RP4) (36 MDa); Salmonella Typhimurium C5, TTr (60 MDa); E. coli 39R861 (24, 50, 98 MDa); and E. coli R27 (110 MDa).
Pulsed field gel electrophoresis
Genetic relatedness among strains within the same serotypes was determined by Pulsed field gel electrophoresis (PFGE) using the XbaI enzyme (New England Biolabs, Beverly, Mass) following the standardized protocol for PulseNet pathogens, 18 and using the Salmonella Braenderup H9812 strain 19 as molecular size marker. The genetic profiles were analyzed with Bionumerics software (Applied Maths, Sint-Martens-Latem, Belgium), applying the Dice coefficient and the Unweighted Pair Group method with Arithmetic Mean method, optimization and tolerance defined in 1.5%, parameters as recommended by PulseNet for usual analysis (CDC, 2000). The 100% similarity cutoff was used to define a restriction profile.
Polymerase chain reaction analysis and DNA sequencing
The total DNA of Salmonella strains was extracted from overnight culture by lysing the bacteria at 95°C and collecting the supernatant after centrifugation. Initially, blaCTX-M, blaSHV, and blaTEM were sought by using primers previously described. 20 Subsequently, the amplification of the blaCTX-M-2 genes was performed by using the primer sets described previously 20 and blaCTX-M-8 genes were detected by using the set primers designed in house (M.F.B., personal communication, October 1, 2014). After polymerase chain reaction (PCR) amplification, all DNA fragments were purified with ExoSapIT (Affymetrix, Inc., Santa Clara, CA) and the amplification products obtained were sequenced with an ABI 3100 sequencer (Applied Biosystems Waltham, MA). The nucleotide sequences obtained were compared to those registered in GenBank (www.ncbi.nlm.nih.gov/blast).
Results
During the study period, 46 (7.3%) of 630 Salmonella enterica strains were β-lactamase producers, of which 30/495 (6.1%) were of human origin, and 16/135 (11.9%; Yates Qui Square = 4.435; p = 0.018) were of nonhuman origin: poultry meat (13), poultry environment (01), beef burger (01), and sludge (01). The most frequent serotype among ESBL producers identified was Salmonella Muenchen (n = 13). followed by Salmonella Typhimurium, including monophasic variant strains (n = 9), and Salmonella Corvallis (11). The remaining strains (n = 10) were identified as belonging to nine serotypes: Salmonella Agona, Salmonella Albany, Salmonella Infantis, Salmonella Enteritidis, Salmonella Kentucky, Salmonella Newport, Salmonella Poona, and Salmonella Saintpaul. Three Salmonella Typhimurium produced only the non-ESBL β-lactamase TEM-1. The distribution of serotypes identified and the isolation source are summarized in Table 1.
Minimum inhibitory concentrations (MICs), mg/L, were determined for ampicillin (R ≥ 32 mg/L), ceftazidime (R ≥ 16 mg/L), cefotaxime (R ≥ 4 mg/L), ceftriaxone (R ≥ 4 mg/L), and cefepime (R ≥ 16 mg/L).
Resistance phenotype defined according to CLSI, 2015.
Genes conferring resistances to β-lactams identified by polymerase chain reaction and sequencing.
Nontransferred plasmid neither by conjugation nor by transformation.
NAL, nalidixic acid; AMI, amikacin; AMC, amoxicillin/clavulanic acid; AMP, ampicillin; ATM; aztreonam, CAZ, ceftazidime; CTX, cefotaxime; CRO, ceftriaxone; CPM, cefepime; CFX, cefoxitin; CIP, ciprofloxacin; C, chloramphenicol; S, streptomycin; GEN, gentamicin; IMP, imipenem; SXT, trimethoprim-sulfamethoxazole; SSS, sulfonamide; and TET, tetracycline.
Antimicrobial resistance
ESBL and TEM-1-producing Salmonella strains revealed high level of resistance to ampicillin (MIC ≥256 mg/L), cefotaxime (MIC range, 16–256 mg/L), ceftriaxone (MIC ≥32 mg/L), and cefepime (MIC range, 24–256 mg/L). For ceftazidime, only two strains (4.3%) were resistant (MIC range, 16–24 mg/L), one carrying the blaCTX-M-8 and blaTEM-1 and one blaSHV-5. Among the 46 Salmonella strains, resistance to sulfonamide (58.7%), tetracycline (58.7%), streptomycin (56.5%), nalidixic acid (54.3%), aztreonam (54.3%), gentamicin (41.3%), trimethoprim-sulfamethoxazole (21.7%), amoxicillin-clavulanate (17.4%), and chloramphenicol (32.6%) was observed by the disk-diffusion method. One strain (2.2%) presented ciprofloxacin resistance. All strains were susceptible to amikacin and to cefoxitin (Table 1).
Resistance genes
The occurrence of ESBL and blaTEM-1 genes in the Salmonella strains is detailed in Table 1. The most prevalent gene detected was blaCTX-M-8 (n = 20), identified mainly in Salmonella Corvallis (10), Salmonella Typhimurium, and monophasic variant (4), Salmonella Enteritidis (2), Salmonella Saintpaul (1), Salmonella Newport (1), Salmonella Albany (1) and Salmonella Kentucky (1), followed by blaCTX-M-2 (n = 19), identified in Salmonella Muenchen (13), Salmonella Typhimurium (3), Salmonella Saintpaul (1); Salmonella Agona (1), and Salmonella Infantis (1).
Association of blaCTX-M-2 and blaTEM-1 genes was detected in three strains (Salmonella Typhimurium, Salmonella Agona, and Salmonella Infantis), along with blaCTX-M-8 and blaTEM-1 in another three strains (S. I 4,5,12:i:-, Salmonella Newport and Salmonella Enteritidis). The occurrence of blaSHV-2 and blaTEM-1 was identified in one Salmonella Typhimurium strain. The blaSHV-5 gene was identified in Salmonella Poona (1) and Salmonella Corvallis (1), while the non-ESBL β-lactamase blaTEM-1 gene alone was detected in Salmonella Typhimurium (3).
Conjugation, plasmid profiles
ESBL and TEM-1 genes were plasmid-encoded in 37 (80.4%) Salmonella strains, as determined by conjugation experiments and confirmed by the detection of respective genes in the transconjugants. Plasmid sizes determined by S1-PFGE ranged from 43 to 232 MDa. S1-PFGE also revealed the occurrence of multiple plasmids (three to four) in three strains; the other 34 strains carried one or two plasmids. Despite several attempts, the remaining nine CTX-M-2-producing Salmonella strains were not able to transfer the ESBL phenotype by conjugation or transformation. The antimicrobial resistance profiles obtained for the transconjugants revealed that diverse non-β-lactam resistance phenotypes were also co-transferred (Table 1).
PFGE
The XbaI PFGE patterns of β-lactamase-producing Salmonella strains are shown in Figure 1. Among the 12 Salmonella Typhimurium and Salmonella Typhimurium monophasic (S. I 4,5,12:i:-) strains, 12 different restriction profiles were detected. For the 13 Salmonella Muenchen strains, 9 of them (69%) belonged to a predominant profile (including the 4 food isolates) and the other 2 profiles presented close relation with the predominant profile (overall similarity higher than 95%). For the 13 Salmonella Corvallis strains, only one restriction profile was identified, as well for the 2 Salmonella Enteritidis strains. PFGE was not useful in discriminating the Salmonella Saintpaul strains since the DNA suffered degradation even with addition of 50 μM thiourea into the electrophoresis buffer.
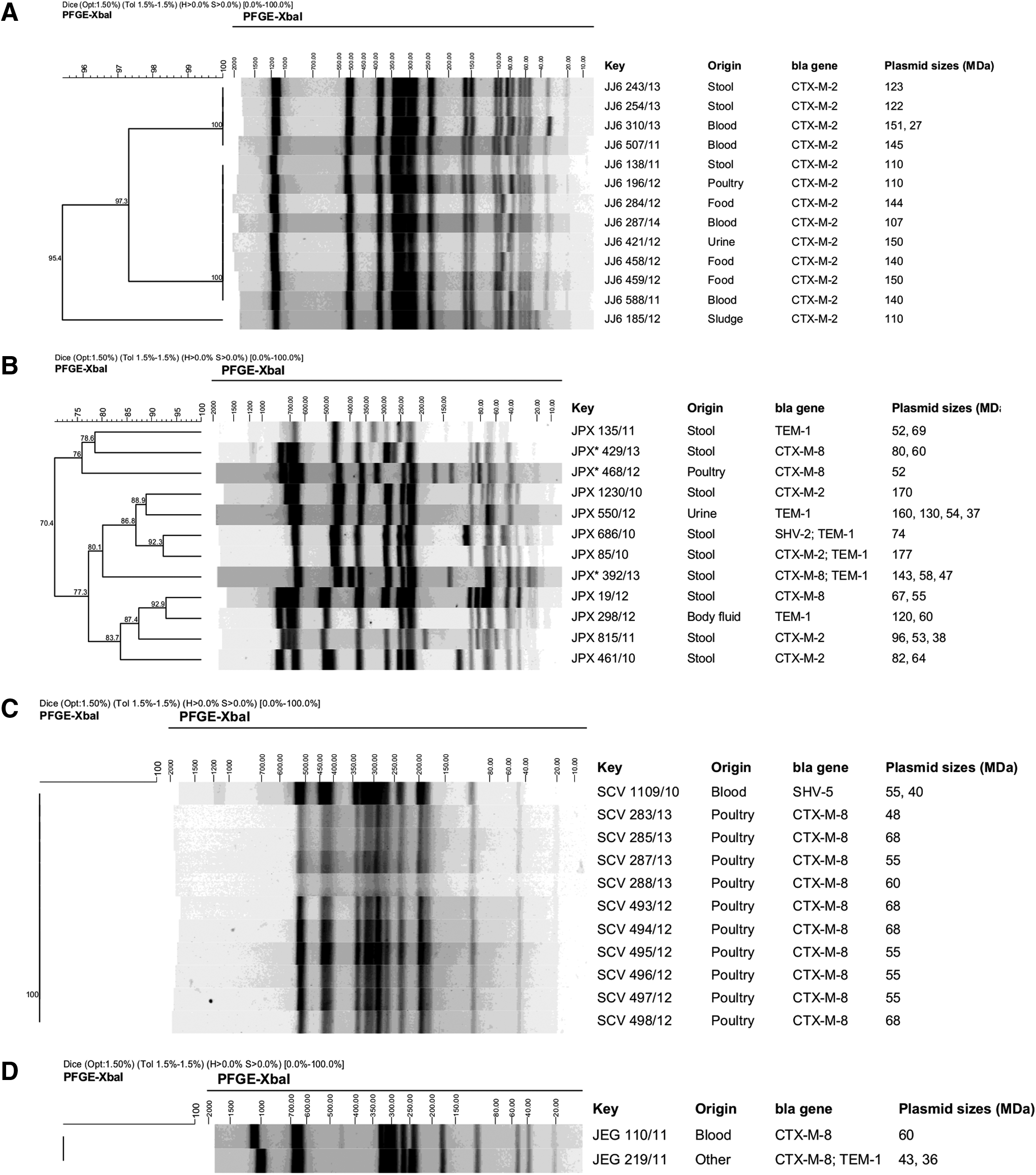
Dendrograms analysis of XbaI-PFGE profiles for β-lactamase producing Salmonella enterica.
Discussion
This study describes the occurrence of ESBL enzymes, mainly CTX-M-2 and CTX-M-8, present among diverse Salmonella serotypes in Brazil over the last 6 years. Most of the ESBL-producing Salmonella strains were recovered from human sources and belonged to 11 different serotypes. Besides the identification of prevalent serotypes in Brazil, Salmonella Typhimurium (and monophasic variant S. I 4,5,12,i:-) and Salmonella Enteritidis,21,22 the occurrence of emerging human serotypes was noteworthy, such as Salmonella Muenchen recovered from invasive infections and nonhuman sources and Salmonella Corvallis from poultry.
ESBL-producing Salmonella serotypes are increasing in prevalence worldwide and represent an emerging problem in the community. 23 The main resistance determinants detected in these strains are the CTX-M enzymes, codified by diverse blaCTX-M genes.1,4,7,24–27
We detected the occurrence of the blaCTX-M-2 gene in isolates belonging to various serotypes: Muenchen, Typhimurium, Saintpaul, Infantis, and Agona. The occurrence of CTX-M-2 in Salmonella Muenchen, an infrequent serotype identified in Brazil,21,22 presenting only three distinguishable patterns as identified by PFGE, with an overall similarity of 95%, suggests the circulation of closely related strains among human and nonhuman sources. The blaCTX-M-2 gene was identified in Salmonella Typhimurium strains isolated from stool samples. Unlike our previous findings, in which we detected the presence of clonal CTX-M-2-producing Salmonella Typhimurium in different Brazilian states, 11 Salmonella Typhimurium strains presented diverse PFGE patterns.
We also detected the blaCTX-M-8 gene among Salmonella Typhimurium and monophasic variant, Salmonella Saintpaul, Salmonella Agona, Salmonella Enteritidis, Salmonella Newport, Salmonella Albany, and Salmonella Kentucky, from human disease, and Salmonella Corvallis from poultry. In the serotypes Salmonella Typhimurium and monophasic variant, blaCTX-M-8-positive strains were genetically unrelated, ruling out the possibility of clonal dissemination. On the other hand, the CTX-M-8 production in 10 Salmonella Corvallis strains revealed to be clonal, but restricted to a single slaughterhouse 28 ; indeed, the presence of other ESBL genes in Salmonella Corvallis from poultry has already been reported. 29 The CTX-M-8-type ESBL was described for the first time in Enterobacteriaceae isolated in Brazil, representing a new variant of CTX-M ESBL. 30 In Salmonella, blaCTX-M-8 gene has been only rarely reported.31–33 Our finding of blaCTX-M-8 in several Salmonella serotypes, most of them associated with plasmids, and the recent reports of change in epidemiology of CTX-M enzymes in Enterobacteriacae from Argentina 9 are a cause of concern for the possible emergence of this resistance determinant in our region.
In lower frequencies, we also identified TEM-1, SHV-2, and SHV-5 enzymes. TEM-1 is not an ESBL, however, the blaTEM-1 gene was detected in different serotypes, including Salmonella Typhimurium belonged to different PFGE patterns. Most of the strains harboring the blaTEM-1 gene also carried blaCTX-M-2, blaCTX-M-8, or blaSHV-2 ESBL genes. Salmonella serotypes co-producing CTX-M and TEM-type β-lactamase have been documented in few case reports, also in Brazil. 33 Besides the occurrence of SHV-2 in only one Salmonella Typhimurium strain, this isolate also carried blaTEM-1 gene, evidencing the presence of multiple β-lactamase genes in a single isolate. Furthermore, blaSHV-5 gene was detected in Salmonella Poona and Salmonella Corvallis serotypes, both of them recovered from human patients with illness.
Successful spread of ESBL genes may arise from their ability to be captured and mobilized by genetic structures, particularly insertion sequences and integrons, often into conjugative plasmids that play an important role to facilitate their dissemination.34,35 Plasmids have been associated with the spread of several β-lactamases genes, mostly ESBL genes identified in strains isolated from human and poultry sources,8,36 underscoring the need to monitor ESBL-encoding plasmids in addition to the ESBL-producing strains among Salmonella enterica from human and nonhuman sources.
We observed a great variability in the numbers and sizes of plasmids circulating among human and nonhuman Salmonella serotypes. Transferability of ESBL phenotypes to recipient strains (Table 1) very likely indicates the location of the ESBL genes on these plasmids, since we verified that all obtained transconjugants harbored the corresponding blaESBL gene.
Besides the cephalosporin resistance conferred by ESBL, we identified additional antimicrobial resistance to non β-lactams agents (sulphonamide, gentamicin, chloramphenicol, tetracycline, sulfamethoxazole, and nalidixic acid) among the ESBL-producing Salmonella isolates, some of them transferable by conjugation or transformation together with the ESBL-encoding plasmids (Table 1). Multiple drug resistance has already been observed among Salmonella Muenchen,37,38 and Salmonella Typhimurium strains. 11 This finding becomes even more worrisome considering that some Salmonella Muenchen strains were isolated from bloodstream infections, negatively impacting available therapeutic options for the treatment of invasive infections. The resistance rates to nalidixic acid, indicating decreased susceptibility to ciprofloxacin, a key antimicrobial used in the treatment of human salmonellosis, was also notable. Indeed, resistance to ciprofloxacin (MIC = 6 mg/L) was detected in CTX-M-8-producing Salmonella I 4,5,12:i:-, a major serotype already reported with different quinolone resistance determinants in our region. 39
The large number of ESBL-producing Salmonella studied herein provides novel information regarding the dissemination of blaESBL genes; remarkably, we report for the first time in Brazil the occurrence of blaCTX-M-8 in the serotypes Typhimurium including monophasic variant, Enteritidis, Corvallis, Newport, Saintpaul, Albany, and Kentucky from human sources (blood, body fluids, urine, and stool), and in Salmonella Corvallis from poultry. Detection of blaCTX-M-2 gene in Salmonella Muenchen and blaSHV-5 gene in Salmonella Poona and Salmonella Corvallis are also reported for the first time in this country here.
In summary, we reported the predominance of CTX-M-2 and CTX-M-8 among several serotypes of Salmonella enterica from human and nonhuman sources. In addition to cephalosporin resistance, resistances to other antimicrobial drugs were observed, reducing the available antimicrobial options. Public health measures are urgently needed to contain the spread of Salmonella serotypes resistant to the key antibiotic classes employed for treatment of severe salmonellosis.
Footnotes
Acknowledgment
This work was supported by the Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP), Brazil (Grants numbers 2012/21709-5 and 2013/14907-8).
Disclosure Statement
Y.D. is a member of the following advisory boards: Meiji, Tetraphase, Merck, Achaogen. Research grant: The Medicines Company. All other authors declare no conflicts.