
Abstract
The extensive use of β-lactam antibiotics has led to emergence and spread of extended-spectrum β-lactamases (ESBLs). This study was conducted to investigate the prevalence of 7 different ESBL genes (blaTEM, blaSHV, blaCTX-M, blaVEB, blaPER, blaGES, and blaOXA-10) and O25b-ST131 high-risk clone among 61 clinical isolates of Escherichia coli. Also, one broad-spectrum β-lactamase (blaOXA-1) was investigated. This study was also constructed to evaluate iodometric overlay method in detection of ESBL production. Phenotypic identification of E. coli isolates using API 20E revealed 18 distinct biotypes. DNA fingerprinting using enterobacterial repetitive intergenic consensus polymerase chain reaction (ERIC-PCR) differentiated all isolates into 2 main phylogenetic groups with 60 distinct genetic profiles. Elevated values of minimal inhibitory concentration (MIC)50 and MIC90 for third- and fourth-generation cephalosporins were observed. Phenotypic tests revealed that 85.24% of isolates were ESBL producers. The incidence rates of blaTEM, blaSHV, blaCTX-M, blaGES, blaOXA-1, and blaOXA-10 among E. coli ESBL producer phenotype were 69.23%, 25%, 96.15%, 3.85%, 11.54%, and 48%, respectively. On the other hand, blaVEB and blaPER were not detected. Sequencing of blaTEM and blaSHV revealed that blaTEM-214 and blaSHV-11 were the most prevalent variants. Group characterization of blaCTX-M revealed that blaCTX-M-1 was the most prevalent group of blaCTX-M family. It was found that 30.77% of E. coli ESBL producers belonged to O25b-ST131 clone harboring blaCTX-M-15. This study concluded that iodometric overlay method was 100% sensitive in detection of ESBL production. To our knowledge, this is the first Egyptian study that declares the emergence of E. coli O25b-ST131 harboring blaGES.
Introduction
T
Both TEM-1 and SHV-1 β-lactamases are class A narrow-spectrum β-lactamases according to the Ambler classification. 6 After discovery of TEM-1 and SHV-1 β-lactamases, new variants of these enzymes with an extended spectrum of activity were discovered. 7 The extended spectrum activity of the new discovered TEM and SHV β-lactamases was found to originate from a mutation in DNA sequence of genes coding for these two enzymes. 8
The extended spectrum activity of new TEM and SHV β-lactamases was found to include resistance to third- and fourth-generation cephalosporins and monobactams; therefore, the term extended-spectrum β-lactamases (ESBLs) were introduced to clinical settings. Both of TEM and SHV β-lactamases are inhibited by clavulanic acid (CA). 6 Another commonly occurring ESBL is the CTX-M type, which also classified as Ambler class A β-lactamases. CTX-M β-lactamases was first discovered in an E. coli isolate recovered from Munich, Germany, in 1989. 9 The CTX-M enzymes exhibit a high hydrolytic activity on cefotaxime, ceftriaxone, and aztreonam. 6 The CTX-M β-lactamases were further classified according to both DNA and amino acid sequences into five groups (CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9, and CTX-M-25). CTX-M β-lactamases are encoded by the chromosome in Kluyvera spp. and by plasmids in E. coli. Recently, new ESBLs of class A β-lactamases, which are not derived from either TEM or SHV β-lactamases, had been reported. These new ESBLs are blaPER, blaVEB, and blaGES/IBC. 10
Pseudomonas extended resistance (PER) ESBLs share only around 25–27% homology with known TEM and SHV-ESBLs. PER-1 was first detected in P. aeruginosa and later in S. enterica serovar Typhimurium and Acinetobacter isolates as well. 7
Vietnamese extended β-lactamase (VEB-1) was originally recovered from a Vietnamese infant hospitalized in France. The gene encoding for blaVEB-1 was found to not be a simple point mutant of any known β-lactamases. VEB-1 β-lactamase has been detected in E. coli, K. pneumoniae, E. cloacae, E. sakazakii, and P. aeruginosa isolates. 7 VEB-1 β-lactamase was found to confer a higher level of resistance to ceftazidime than cefotaxime. 11
Guiana extended spectrum (GES-1) was first described in 2000 in K. pneumoniae isolate recovered from French Guiana. 12 The hydrolytic profile of GES-1 is similar to other class A ESBLs, but unlike most ESBLs, GES-1 was found to not hydrolyze aztreonam. 13
Apart from class A ESBLs, OXA-type ESBLs, which are categorized as class D β-lactamase, 6 were originally detected in P. aeruginosa isolates recovered from a single hospital in Ankara, Turkey. 14 OXA-derived ESBLs have more hydrolyzing ability to ceftazidime than cefepime.7,15
Most OXA-type β-lactamases do not hydrolyze the extended-spectrum cephalosporins to a significant degree and are not regarded as strong ESBLs. However, OXA-10 hydrolyzes (weakly) cefotaxime, ceftriaxone, and aztreonam, resulting in a reduced susceptibility to these antibiotics. 16
Recently, a problematic high-risk clone of E. coli has been reported worldwide, representing a major public health problem. This clone was found to belong to extraintestinal phylogenetic group B2, sequence type 131 and serotype O25b. The E. coli O25b-ST131 clone was found to have high potentials for virulence and antibiotic resistance. This clone was found to be highly associated with blaCTX-M, especially blaCTX-M-15.17,18
To our knowledge from available cited literature, few studies have been done in Egypt, investigating ESBL genes among E. coli clinical isolates belonging to O25b-ST131 clone; therefore, the present study was conducted to investigate the prevalence of blaTEM, blaSHV, blaCTX-M, blaPER, blaVEB, blaGES, blaOXA-1, blaOXA-10, and O25b-ST131 clone among E. coli clinical isolates recovered from Egyptian patients. Also, this study aimed to evaluate the iodometric overlay method in detection of ESBL production.
This study indicated a high prevalence of E. coli O25b-ST131 clone harboring ESBL-encoding genes among genetically diverse E. coli clinical isolates genotyped by ERIC-PCR. Iodometric overlay method was found to be a sensitive method for detection of ESBL production.
Patients and Methods
Bacterial strains
A total of 61 nonduplicate clinical isolates of E. coli were selected from 301 isolates of Gram-negative bacilli, which were recovered from 153 cases. The 61 isolates of E. coli were collected from 55 cases (31 females and 24 males aging between of 7 months and 85 years) who admitted or attended to different medical departments at Ain-Shams University hospital, Cairo, Egypt, over a period of 1 year (from May 2012 to April 2013). Isolates of E. coli were recovered from ascitic fluid (n = 1), blood (n = 4), throat swab (n = 4), endotracheal tube (n = 1), sputum (n = 10), urinary catheter (n = 1), urine (n = 14), wound (n = 24), pus (n = 1), and cerebrospinal fluid (n = 1).
Isolation and identification of E. coli isolates
All strains of E. coli were primarily isolated on MacConkey's agar (Oxoid, United Kingdom) and then on eosin methylene blue agar (Scharlau, Spain). The isolated strains were identified phenotypically using API 20E (Biomerieux, France) and genotypically through amplification of specific E. coli uidA gene and sequencing of 16S rRNA gene using primers and cycling conditions as listed in Table 1.
F, forward; R, reverse; TEM, Temoneira; SHV, sulfhydryl variable; CTX-M, cefotaximase-Munich; GES, Guiana extended spectrum; PER, Pseudomonas extended resistance; VEB, Vietnamese extended spectrum; OXA, oxacillinase; uidA, glucourindase; ERIC, enterobacterial repetitive intergenic consensus; pabBspe, para-amino benzoic acid.
Genotyping of clinical isolates
Clonal relatedness between clinical isolates of E. coli was determined by ERIC-PCR. The primers were obtained from Macrogen (Korea, Geumcheon-gu, Seoul) .Gene amplification was carried out according to cycling conditions listed in Table 1 using Mastercycler® personal (Eppendorf, CA).
Fingerprint pattern analysis
Banding pattern generated by ERIC-PCR was analyzed using BioNumerics7.5 software (Applied Maths, Kortrijk, Belgium) using Dice (similarity) coefficient. Cluster analysis was performed based on unweighted pair-group method with arithmetic averages (UPGMA) with position tolerance at 0.15.
Antimicrobial susceptibility testing
All E. coli isolates were tested for their susceptibility against 13 β-lactams and 3 β-lactams/β-lactamase inhibitor combination. Antimicrobial susceptibility testing was determined by Kirby-Bauer disc diffusion method using Mueller-Hinton agar (MHA) (Oxoid, United Kingdom). 19 Broth microdilution method was also performed using the cation-modified Mueller-Hinton broth (Oxoid, United Kingdom) to determine the minimal inhibitory concentration (MIC) values of tested antibiotics. 20 The results were interpreted according to guidelines of clinical laboratory standards institute (CLSI), 2014. 21 Thirteen different β-lactam antibiotics representing all classes of β-lactams as well as 3 β-lactams/β-lactamase inhibitor combination were also tested. Both E .coli ATCC 25922 and Klebsiella pneumoniae ATCC 700603 were used as quality control strains.
Initial screening of ESBL production
Initial detection of potential ESBL production was determined according to guidelines of CLSI, 2014, that is, resistance to either cefotaxime or ceftazidime or ceftriaxone or aztreonam, 22 and the iodometeric overlay method as described below.
Confirmation of ESBL production according to CLSI, 2014
Confirmation of ESBL production according to guidelines of CLSI, 2014, was briefly done by determining the MIC of cefotaxime (CTX) and ceftazidime (CAZ) alone and in combination with 4 μg/ml CA .Isolates which showed ≥3 twofold decrease in MIC values of either cefotaxime or ceftazidime after addition of CA were considered to be ESBL producers. 22
Confirmation of ESBL production by double disc synergy test
Briefly, a disc of amoxicillin/CA (30 μg) was placed at the center of MHA plate and one disc for each of CAZ (30 μg) and CTX (30 μg) was placed 25–30 mm apart from each side of amoxicillin/CA disc (Fig. 1). The plates were incubated at 37°C for 18 hr. An isolate was considered to be an ESBL producer when an expanded zone of inhibition was observed around either ceftazidime and/or cefotaxime disc from side toward the amoxicillin/clavulanic disc. 23

Phenotypic tests for detection of ESBL production. Tests:
Confirmation of ESBL production by modified CLSI ESBL confirmatory test
The modified confirmatory test was performed by placing discs of CAZ or CTX loaded with boronic acid (BA) and EDTA at one side of MHA plate inoculated with the tested isolate. Another group of discs contained CAZ or CTX loaded with CA, BA, and EDTA at the other side of the plate (Fig. 1). Positive result was considered when an augmentation of ≥5 mm in the inhibition zone diameter, of either CTX-CA-BA-EDTA or CAZ-CA-BA-EDTA, compared to the inhibition zone diameter of either CTX-BA-EDTA or CAZ-BA-EDTA, respectively. 24
Iodometric overlay method for initial detection and confirmation of ESBL production
All clinical isolates of E. coli were toothpicked onto MHA plates and allowed to grow for 18 hr at 37°C. The plates were then overlaid with 10 ml of 1% agarose containing 0.05% toluene, 0.2% soluble starch, and 1% of antibiotic substrate (cefotaxime, ceftazidime, and ceftriaxone). The plates were then incubated at 37°C for 15 min. The plates were then covered with Lugol's iodine solution for 2–4 min at room temperature, and then Lugol's iodine solution was discarded out. 25 The same procedure was repeated with the addition of 0.04% CA to the previously mentioned overlay mixture containing each substrate. The isolates were considered to be potential ESBL producer when well-defined halo zones were observed around toothpicked colonies in plates containing the antibiotic substrate only (Fig. 1). On the other hand, the isolates were confirmed to be ESBL producers when either well-defined halo zones around toothpicked colonies disappeared or the diameter of halo zone around the toothpicked colonies decreased in the plate containing the substrate and CA, when compared to the plate containing substrate only.
Genotypic detection of ESBLs and broad spectrum β-lactamase
All clinical isolates of E. coli ESBL phenotype were screened for class A ESBLs (blaTEM, blaSHV, blaCTX-M, blaVEB, blaPER, and blaGES) and class D ESBL (blaOXA-10). Broad-spectrum blaOXA-1 was also investigated. All the previously mentioned β-lactamase genes were investigated using primers and cycling conditions listed in Table 1.
Group characterization of blaCTX-M
All isolates that tested positive for blaCTX-M were subjected to further characterization to identify the genetic group of detected blaCTX-M using primers and cycling conditions listed in Table 1. All isolates that tested positive for group I blaCTX-M were also screened for blaCTX-M-15.
PCR and preparation of DNA template
Colony PCR method was employed as previously described by Mirhendi et al. in which a colony of bacterial isolate was picked up with a small tip of micropipette and dipped onto the bottom of 0.2 ml PCR tube; this colony represented the DNA template 26 to which the PCR reaction mix (HOT FIREPol® Blend Master Mix, Solis BioDyne, Tartu, Estonia) was added. The PCR reaction was done in a final reaction volume of 20 μl according to the instructions of the manufacturer. The 20 μl reaction mixture contained 4 μl of 5× master mix, 0.6 μl of forward primer (10 pmol/μl), 0.6 μl of reverse primer (10 pmol/μl), and 14.8 μl distilled water.
Extraction and purification of PCR products for DNA sequencing
All PCR products of blaTEM, blaSHV, and 16S rRNA of E. coli were extracted from agarose gel using the GenElute Gel Extraction® kit (Sigma-Aldrich). The extracted PCR products were purified using the ExoSAP-IT One Step PCR Clean-up® Kit (GE Healthcare Limited).
DNA sequencing
The purified extracted PCR products were sequenced by Macrogen (Korea, Geumcheon-gu, Seoul) using the 3730xl DNA Analyzer® (Thermo Fischer Scientific) and forward primers used in amplification step. The released DNA sequences were corrected using molecular evolutionary genetic analysis (MEGA®) 6 software (Biodesign institute, Tempe).
Identification of corrected DNA sequences
The corrected DNA sequences were uploaded to National Center of Biotechnology and Information (NCBI) using the nucleotide Basic Local Alignment Search Tool (BLAST) to identify the highly similar cited sequences to tested sequences. 27
Phylogenetic analysis of sequenced genes
All corrected DNA sequences of blaTEM and blaSHV were subjected to phylogenetic analysis using JalViwe 2® software (University of Dundee, Scotland, United Kingdom) in which multiple sequence alignment was performed using the ClustalO web service of JalViwe 2 software to construct neighbor-joining tree to determine genetic relatedness between sequenced genes. 28
Molecular detection of E. coli O25-ST131 clone
All identified ESBL producers were screened for ST131 clone using pabBspe primers to amplify pabB gene. The pabB gene is one of the genes used in the Institut Pasteur MLST (multilocus sequencing typing). The nucleotides at the 3′ end of both forward and reverse pabBspe primers (underlined nucleotides) were designed to be only specific for the B2 subgroup. Also, all ESBL producers were screened for O25b serotype through amplification of specific rfbO25 gene using primers and cycling conditions as listed in Table 1.
Results
Isolation and identification
Out of the 301 recovered isolates, 61 (20.27%) were E. coli, 114 (37.87%) were Klebsiella spp., 48 (15.95%) were Proteus spp., 38 (12.62%) were P. aeruginosa, 12 (3.99%) were A. baumannii, 9 (2.99%) were S. marcescens, 6 (1.99%) were Ent. cloacae, and 3 (0.99%) were single isolates of P. stuartii, B. cepacia, and A. hydrophilia.
Phenotypic identification of E. coli strains
All E. coli isolates were phenotypically identified using API 20E revealing 18 different biotypes (Fig. 2). The most prevalent phenotypes were 5144572, 5144552, 7044572, 5044552, 7144552, and 5044572, which occurred at a prevalence rate of 29.51% (18/61), 11.48% (7/61), 11.48% (7/61), 9.84% (6/61), 8.20% (5/61), and 4.92% (3/62), respectively. Other detected biotypes were 7144572, 1044532, and 7044570, which occurred at a prevalence rate of 3.28% (2/61) for each. The lowest detected biotypes were 7144510, 7140512, 7044552, 5144532, 5144512, 1044552, 4144552, 5044542, and 5044563, which occurred at a prevalence rate of 1.64% (1/61) for each.
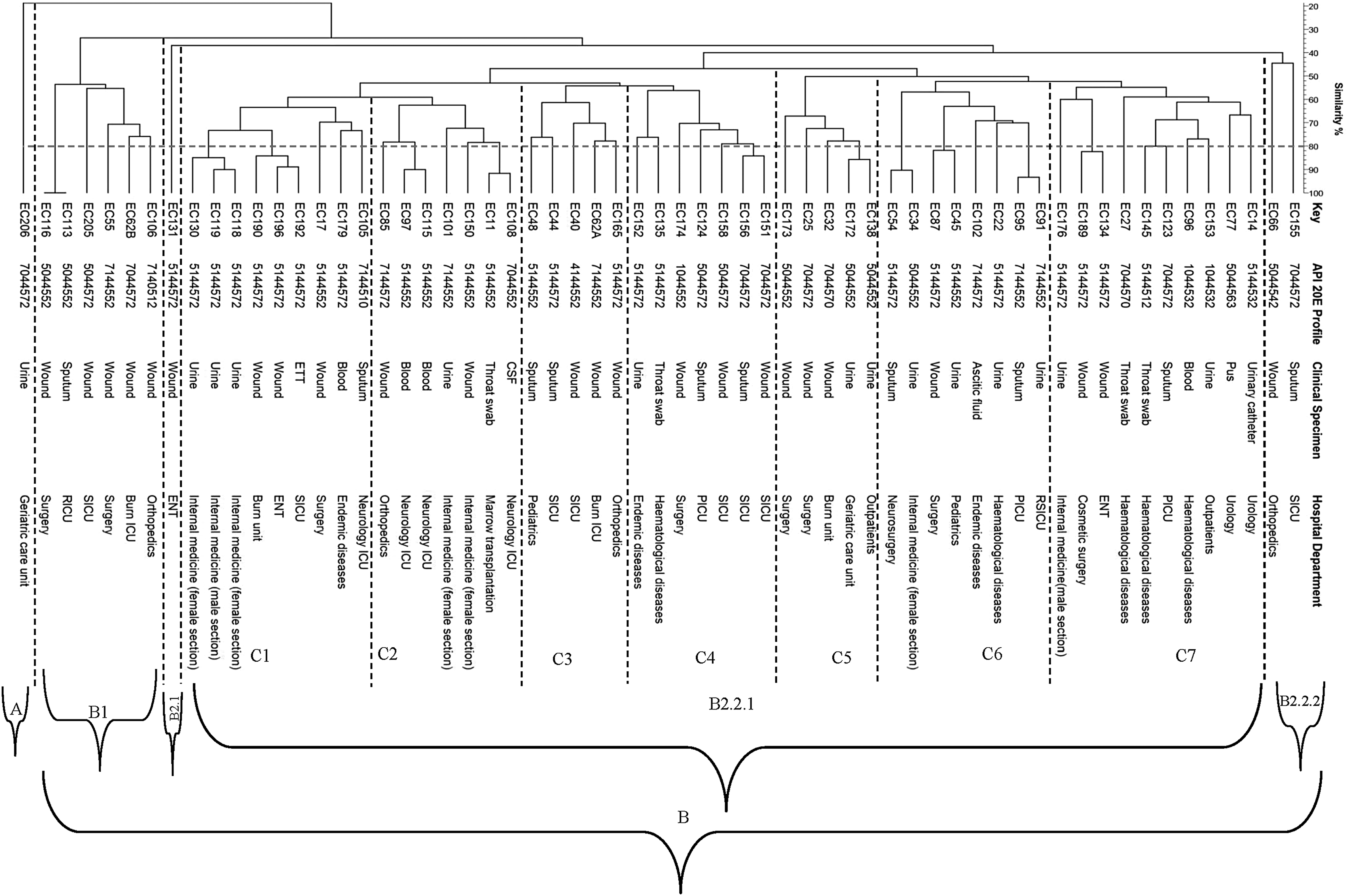
Dendrogram showing clonal relationship between clinical isolates of Escherichia coli.
Genotypic identification of phenotypically identified E. coli isolates
All phenotypically identified E. coli isolates were 100% (61/61) E. coli by genetic identification methods in which 82.24% (52/61) of isolates were tested positive for the specific E. coli uidA gene, while 17.76% (9/61) tested negative. The nine isolates that tested negative for the specific E. coli uidA gene were subjected to amplification and sequencing of 16S rRNA gene, which confirmed that these isolates were E .coli.
Genotyping of E .coli clinical isolates using ERIC-PCR
It was found that ERIC-PCR was able to differentiate the 61 E. coli isolates into 60 unique profiles. It was found that the two isolates (EC113 and EC115) showed an identical profile, although they recovered from different specimens and different hospital departments.
Fingerprint pattern analysis
Generated UPGMA dendrogram according to Dice (similarity) coefficient revealed that all tested E. coli isolates were classified into two major phylogenetic groups (group A and group B). Phylogenetic group A was found to include only one isolate (EC206), while phylogenetic group B was found to include the remaining 60 isolates. Phylogenetic group B was further classified into two major subgroups (group B1 and B2). Subgroup B2 was further classified into two subgroups (B2.1 and B2.2). Subgroup B2.2 was found to include seven clusters (Fig. 2).
Antimicrobial susceptibility
Apart from the two tested carbapenems (imipenem and meropenem) to which 93.44% (57/61) and 95.08% (58/61) of isolates were, respectively, sensitive, the present study revealed that 47.54% (29/61) to 90.16% (55/61) of isolates were resistant to the other tested β-lactams, whether they tested individually or in combinations with β-lactamase inhibitors, as shown in Table 2.
β
MIC, minimal inhibitory concentration.
The highest incidence of resistance was accounted for amoxicillin and ampicillin, where 90.16% (55/61) of the isolates were resistant to them as shown in Table. 2. The incidences of resistance for the tested β-lactam/β-lactamase inhibitor combinations were 83.61%, 75.41%, and 47.54% for ampicillin/sulbactam, amoxicillin/CA, and cefoperazone/sulbactam, respectively.
The incidence of multiple resistance to 2-15 β-lactams was detected in 93.44% of isolates (57/61), while resistance to 5-15 β-lactams was detected in 83.61% of isolates (51/61) as shown in Table 3. Three isolates were resistant to all the tested β-lactams and four isolates were resistant to only one β-lactam. Only 2 isolates were sensitive to all the tested 15 β-lactams. Apart from imipenem and meropenem, the MIC50 and MIC90 of tested β-lactams ranged between 32 to >1,024 μg/ml and 128 to >1,024, respectively, as shown in Table 2.
EC, Escherichia coli; AMX, amoxicillin; AMC, amoxicillin/clavulanic acid; SAM, ampicillin/sulbactam; CE, cephradine; CEF, cefazolin; CXM, cefuroxime; CTX, cefotaxime; CAZ, ceftazidime; CPZ, cefoperazone; CPZS, cefoperazone/sulbactam; CRO, ceftriaxone; ESBL, extended-spectrum β-lactamase; FEP, cefepime; ATM, aztreonam; IPM, imipenem; MEM, meropenem.
Initial screening of ESBL production
According to guidelines of CLSI, 2014, and the iodometric overlay method, this study revealed that 85.24% (52/61) of isolates were considered to be potential ESBL producers.
Phenotypic confirmatory tests of ESBL production
This study revealed that 61.53% (32/52) of potential ESBL producers were confirmed to be ESBL by DDS test, while 100% (52/52) of potential ESBL producers were confirmed to be ESBL producers by the modified CLSI ESBL confirmatory test and iodometric overlay method.
Genotypic detection of ESBLs and broad spectrum β-lactamase
Investigation of phenotypically confirmed E. coli ESBL producers revealed that 69.23% (36/52), 25% (13/52), 96.15% (50/52), 3.85% (2/52), 11.54% (6/52), and 48% (25/52) of these isolates were carrierrs for blaTEM, blaSHV, blaCTX-M, blaGES blaOXA-10, and blaOXA-1, respectively, as shown in Table 3. On the other hand, blaVEB and blaPER were not detected.
Group characterization of blaCTX-M
This study revealed that group I of blaCTX-M family was the most prevalent group in which 82% (41/50) of blaCTX-M producers belonged to blaCTX-M-1. On the other hand, blaCTX-M-2 and blaCTX-M-9 were detected at a prevalence rate of 30% (15/50) and 20% (10/50), respectively, as shown in Table 3.Both blaCTX-M-8 and blaCTX-M-25 were not detected. The most prevalent variant of group I blaCTX-M was blaCTX-M-15 in which 95.12% (39/41) of blaCTX-M-1 producers were tested positive for blaCTX-M-15 as shown in Table 3.
DNA sequencing of blaTEM and blaSHV
The sequencing of all detected blaTEM revealed seven different variants (blaTEM-34, blaTEM-57, blaTEM-135, blaTEM-176, blaTEM-206, blaTEM-207, and blaTEM-214), which occurred at a prevalence rate of 2.78% (1/36), 13.89% (5/36), 5.56% (2/36), 2.78% (1/36), 13.89% (5/36), 5.56% (2/36), and 55.56% (20/36), respectively. On the other hand, sequencing of all detected blaSHV revealed three different variants (blaSHV-11, blaSHV-12, and blaSHV-48), where blaSHV-11 was the most prevalent variant that was detected at a rate of 84.62% (11/13). Both blaSHV-12 and blaSHV-148 occurred at a prevalence rate of 7.69% (1/13) for each.
DNA sequencing of 16S rRNA gene of phenotypically identified E. coli isolates
Sequencing of 16S rRNA gene of all E. coli isolates that tested negative for the specific E. coli uidA gene revealed that these isolates were E. coli in which the BLAST results showed that the sequence of 16S rRNA gene of tested isolates have 96–99% sequence similarity to 16S rRNA sequence of different E. coli strains as cited in GeneBank. On the other hand, 16S rRNA gene of only one isolate showed a relatively low sequence similarity (81%) with E-value = 2e−23 with 16S rRNA gene of E. coli strain CRMY9 (GeneBank: KF574806.1).
Phylogenetic analysis of sequenced genes
Phylogenetic analysis of detected blaTEM showed that all identified blaTEM were classified into two phylogenetic groups in which group I was found to include one cluster, while group II was found to include two clusters. On the other hand, phylogenetic analysis of detected blaSHV showed that all detected variants of blaSHV were classified in one phylogenetic group containing two main clusters.
Molecular detection of E. coli O25b-ST131 clone harboring blaCTX-M-15
All E. coli ESBL producers were screened for the O25b-ST131 clone, where it was found that 44.23% (23/52) of ESBL producers belonged to the O25b serotype and 92.31% (48/52) belonged to the ST131 clone. Overall, 42.31% (22/52) of E. coli ESBL producers belonged to O25b-ST131 clone. It was found that 30.77% (16/52) of E. coli ESBL producers belonged to the O25b-ST131 clone harboring blaCTX-M-15 as shown in Table 3.
Discussion
Infections caused by ESBL-producing bacteria have increased in recent years. Detection of ESBL production among hospital and community isolates has great importance for infection control practitioners and clinicians to select the most appropriate antibiotic as well as establish new policies to restrict the spread of such isolates, especially in a hospital environment. 29
Great attention has been focused on ESBL production among E. coli isolates as they are the most frequently recovered enterobacterial isolate from both hospitalized and nonhospitalized patients. 30
This study revealed a high prevalence of ESBL genes among E. coli clinical isolates recovered from patients who were hospitalized or attended to Ain-Shams University Hospital, Cairo, Egypt, between May 2012 and April 2013, where this study demonstrated that 85.25% (52/61) of investigated isolates were found to harbor ESBL genes. A similar high prevalence rate of ESBL genes among E .coli clinical isolates was reported by a Korean study. This Korean study was performed on 142 clinical isolates of E. coli recovered from infected or colonized patients who were hospitalized or attended to Daejeon St Mary's Hospital, Korea, between January 2008 and September 2009. The Korean study revealed that 97.89% (139/142) of investigated isolates were found to harbor ESBL genes. 31
Compared to this study, relative low prevalence rates of ESBL producers were observed in two recent Egyptian studies, which demonstrated that 54.5% (30/55) and 52% (47/90) of E. coli were, respectively, found to be ESBL producers.32,33 The previously mentioned rates of ESBL producers among E. coli isolates were greater than the rates detected in Khartoum teaching hospital, where 45.1% (32/71) of E. coli isolates were ESBL producers. 34 All the previous findings were far from the findings of an Iranian study, which demonstrated a low rate of 30% (100/327) of ESBL producers among E. coli isolates. 35
This study revealed that the DDS test was not a sensitive method for detection of ESBL production in which only 61.53% (32/52) of genetically identified ESBL producers were tested positive for the DDS test, indicating the coexistence of other types of β-lactamase (MBL and/or AmpC β-lactamase) along with ESBL, which masked the detection of ESBL by the DDS test. 24
The previous finding was made sure when the modified CLSI ESBL confirmatory test was performed, where it was found that 100% (52/52) of genetically identified ESBL were also tested positive for the CLSI ESBL confirmatory test. So, we recommend the use of BA and EDTA along with CA to detect ESBL production which can be masked by MBL and/or AmpC β-lactamase. 24
This study revealed that the sensitivity of DDS test for detection of ESBL was inconsistent with the findings of an Indian study, which revealed that the sensitivity of DDS test was 95% (123/129). 36
On the other hand, the sensitivity of modified CLSI ESBL confirmatory test for detection of ESBL production among genetically identified E. coli ESBL producers was closely related to the findings of a Greece study, which showed that 97.5% (158/162) of isolates that genetically identified as ESBL producers were tested positive for the modified CLSI ESBL confirmatory test. 24
The iodometric overlay is considered to be an accurate, reproducible, easily readable, and economical method for detection of β-lactamase production25,37; this method was used only for detection of β-lactamase production using penicillin G as a substrate without testing its ability to detect ESBL production. 38
To our knowledge, this study is considered the first to employ and evaluate the iodometric overlay method for detection of ESBL production using third-generation and fourth-generation cephalosporins as substrates and CA as an ESBL inhibitor.
The sensitivity of iodometric overlay method for ESBL detection among ESBL producers was 100% (52/52). Overall, the present study revealed that the iodometric overlay method is more sensitive for detection of ESBL production than the DDS test. Also, this study showed that the sensitivity of iodometric overlay method is similar to the sensitivity of modified CLSI ESBL confirmatory test.
Until the end of the previous millennium, ESBL genes were commonly detected among of K. pneumoniae than E. coli isolates, but during the last decade, it was observed that ESBL genes became mainly detected among E. coli isolates. 39
This study revealed that the most prevalent ESBL gene among E. coli ESBL producers was blaCTX-M followed by blaTEM in which the prevalence rates of blaCTX-M and blaTEM were 84.62% (44/52) and 62.23% (36/52), respectively. The previous findings were inconsistent with the findings of Rezai et al. in which blaTEM was more prevalent than blaCTX-M in which 49% (49/100) and 28% (28/100) of E. coli ESBL producers were, respectively, blaCTX-M and blaTEM producers. 35
On the other hand, a recent Egyptian study was found to be consistent with our finding in which 87.8% (65/74) of E. coli isolates were blaCTX-M producers. 40 In addition to the previous findings, a very high prevalence rate of blaCTX-M was demonstrated in a Thai study in which 99.6% (234/235) of ESBL-producing E. coli were blaCTX-M producers. 41
This study revealed a low prevalence rate of blaSHV, 25% (13/52), a similar low prevalence rate of blaSHV was reported in a recent Egyptian study, which declared that the prevalence of blaSHV was 14% (6/42). 40 The previous findings were relatively contradicted with two Iranian studies, which revealed relative high prevalence rates of blaSHV, 34.8% (48/138) and 44% (44/100);35,42 also, our results were contradicted with the Egyptian and Thai studies, which did not detect blaSHV among E. coli isolates.32,43
This study revealed that blaPER was not detected; this finding was consistent with the finding of Moghaddam et al. who also did not detect blaPER among E. coli isolates. 44 This study did not detect blaVEB; this finding was contradicted with the findings of Udomsantisuk et al. and Rezai et al. in which the prevalence rates of blaVEB were 16.7% (6/36) and 8% (8/100), respectively.35,43
To our knowledge, there is no data about prevalence of blaGES among E. coli strains in Egypt. This is first Egyptian study that announces the detection of blaGES among clinical isolates of E. coli. Only 3.85% (2/52) of ESBL-producing isolates were found to harbor blaGES-1. The previous findings were contradicted with the finding of a Thai study which did not detect blaGES. 41
This study declared a high prevalence rate of blaCTX-M-15 in which 75% (39/52) of E. coli ESBL producers were found to harbor blaCTX-M-15; this finding was closely related to the finding of a Canadian study, which revealed that 71% (148/209) of E. coli ESBL producers were blaCTX-M-15 producers. 45 A relative high prevalence of blaCTX-M-15 was also observed in a German study in which blaCTX-M-15 was detected in 55.55% (55/99) of E. coli ESBL producers. 46
To our knowledge, the prevalence rates of blaOXA-1 and blaOXA-10 among clinical isolates of E. coli were not determined in Egypt due to limited number of surveillance studies examining such two genes among E. coli isolates. Overall, this study revealed that 11.54% (6/52) of ESBL-producing isolates were found to harbor blaOXA-10; this finding was closely related to a recent Thai study, which demonstrated that 8.1% (19/235) of ESBL-producing E. coli were carriers for blaOXA-10. 41
Interestingly, blaOXA-1-like was also investigated among ESBL-producing isolates where it was found that 48% (25/52) of isolates were tested positive. The previous finding was contradicted with the findings of recent Moroccan and Indian studies, which, respectively, demonstrated that 90% (9/10) and 75% (9/12) of ESBL-producing E. coli isolates were carriers for blaOXA-1.47,48
The E. coli ST131 clone was found to be associated with a high ability to resist a wide range of antimicrobial agents; also, this clone is correlated with high virulence and ability to spread. The E. coli ST131 clone was found to be strongly associated with ESBLs. 49
This study revealed that 92.30% (48/52) of E. coli ESBL producers belonged to the ST131 clone and 45.83% (22/48) of this clone belonged to the O25b serotype. The previous findings were less comparable to the findings of a Danish study, which declared that 38% (44/115) and 95% (42/44) of ESBL-producing E. coli, respectively, belonged to the ST131 clone and O25b serotype. 50 In contrast to the previous findings, a low prevalence rate of E. coli ST131 was detected in a recent Turkish study, which revealed that 12% (35/294) of E. coli isolates belonged to the ST131 clone. 49
This study showed that 72.73% (16/22) of E. coli isolates that belonged to the O25b-ST131 clone were blaCTX-M-15 producers; a similar high prevalence was observed in a Bulgarian study in which 96% (129/134) of E. coli O25b-ST131 clone were blaCTX-M-15 producers. 51
To our knowledge, this is the first Egyptian study that addressed the emergence of blaGES among E. coli isolates recovered from Egyptian patients. This study demonstrated that the rate of blaGES was 3.85% (2/52). The previous finding was contradicted with the Iranian study that did not detect blaGES among ESBL-producing E. coli isolates. 35 Also, our findings were inconsistent with the results of a recent Lebanese study carried out on 242 clinical isolates of E. coli recovered from fecal specimens from patients with diarrhea between February 2013 and May 2014; this study did not detect blaGES among investigated isolates. 52
Genotyping of clinical isolates using ERIC-PCR is considered to be faster and more cost-effective than pulsed-field gel electrophoresis or multilocus sequencing for providing conclusive data about the genetic or clonal similarity between investigated bacterial isolates; also, ERIC-PCR possesses a higher discriminatory ability compared with other quick-typing techniques. 53
Genotyping of E. coli clinical isolates using ERIC-PCR revealed that E. coli isolates were genetically diverse and heterogeneous. This finding was expected as the isolates were collected from different buildings and medical department at Ain-Shams University hospital. A similar finding was reported by Britain, Malaysian, and Italian studies.54–56
This study revealed that 96.72% (59/61) of isolates displayed a single profile, indicating different origins of dissemination; whereas only 3.28% (2/61) of isolates showed an identical DNA fingerprint pattern, indicating a similar origin of dissemination.
ERIC-PCR profiles did not demonstrate that there is no genetic relationship between 59 isolates. Therefore, infection or colonization by these isolates was not clonally spread and results from individual cases reflecting good infection control practice inside Ain-Sham University hospital. Similar findings were observed in an Iranian study. 57
This study revealed that the amplification of uidA gene for genotypic identification of E. coli isolates was not 100% sensitive, where only 85.24% (52/61) of phenotypically identified E. coli isolates were tested positive for the uidA gene. The previous finding was consistent with the finding of Martins et al. who reported that 97.3% (425/435) of E. coli isolates collected from different sources were tested positive for the uidA gene, that is, 2.7% (10/435) cannot be identified by amplification of uidA gene. 58
Conclusion
There is an obvious high prevalence of ESBL genes among genetically diverse E. coli isolates included in this study. DNA sequencing revealed that blaTEM-214 and blaSHA-11 were the most prevalent variants of detected TEM and SHV ESBLs. This is the first study that detects ESBL production by the iodometric overlay method with 100% sensitivity. Genotypic identification of E. coli through amplification of specific E. coli uidA gene was not 100% sensitive. This study reports the first emergence of O25b-ST131 clone harboring blaGES in Egypt.
Footnotes
Acknowledgment
I would like to express my sincere thanks and deepest gratitude to Dr. Ahmed S. Abdel-Moneim, Professor of Virology, Faculty of Veterinary Medicine, Beni-Suef University, for his continuous guidance regarding interpretation of DNA sequencing results. Authors thank Mr.Youhana Ekladious Takyi, Mrs.Wafaa Ibrahim Noor, Mr. Khalid kamel Abd-Elkhalek, and Mr. Yassin Farag Mahmoud, Department of Microbiology, Ain Shams University educational hospital, Cairo, Egypt, for their great help in collection of clinical specimens and clinical data.
Disclosure Statement
No competing financial interests exist.