
Abstract
Aim:
The purpose of this study was to characterize the New Delhi metallo-beta lactamase (NDM)-7-producing Enterobacteriaceae isolated in the Arabian Peninsula.
Methods:
Enterobacteriaceae identified to carry blaNDM-7 in a collection of 157 NDM-producing isolates from Kuwait, Oman, Saudi Arabia, and the United Arab Emirates (UAE) were investigated for their antibiotic and disinfectant susceptibility, and resistance gene content. The virulence profile, phylogenetic and sequence types of the isolates were also determined. The plasmids carrying the blaNDM-7 were transferred, and their complete nucleotide sequence was determined.
Results:
Four NDM-7-producing Escherichia coli isolated in Kuwait, Oman, and the UAE, respectively, were identified. They were clonally unrelated, carried a few virulence determinants only, and belonged to clonal complexes CC10 and CC23, or ST448. They were all multi-drug resistant but remained susceptible to fosfomycin, tigecycline, and colistin. In all isolates, blaNDM-7 was located on IncX3 type plasmids of a variable size, not harboring any further resistance genes. The plasmids exhibited a high degree of similarity to each other and to pKpN01-NDM7 from Canada, with various size deletions and insertions.
Conclusions:
Our findings show that IncX3 type plasmids play an important role in the spread of the currently rare NDM-7 variant in the Arabian Peninsula. This association of blaNDM-7 with the IncX3-type plasmid is particularly worrisome, as this type of plasmid was proved to spread other carbapenemases in various species of Enterobacteriaceae worldwide at a high efficiency.
Introduction
S
Although NDM-producing Enterobacteriaceae are endemic in many countries of the Arabian Peninsula, 4 little is known about the variants of the enzyme and their genetic support. In the present study, we aimed at revealing the molecular context of blaNDM-7 in Enterobacteriaceae isolates from Kuwait, Oman, Saudi Arabia, and the UAE.
Materials and Methods
Strains
A collection of 157, independent NDM-producing Enterobacteriaceae isolated between 2009 and 2013 were included in the study. Of this collection, 100 strains, that is, 6 from Kuwait, 30 from Oman, 32 from the Kingdom of Saudi Arabia, and 32 from the UAE, were reported earlier as NDM producers 4 whereas 16 and 41 isolates from Kuwait and from the UAE, respectively, were new isolates. All samples were taken based on clinical indications, and strains were submitted to us coded, without any patient identifiers.
Antibiotic susceptibility assay
Susceptibility to most antibiotics was tested by microdilution and interpreted by CLSI standards. 17 Fosfomycin susceptibility was tested by agar dilution. For the majority of antibiotics, CLSI clinical breakpoints were used for interpretation 17 with the exception of colistin and fosfomycin, which were interpreted by using the EUCAST criteria. 18 The minimum inhibitory concentration (MIC) of chlorhexidine, benzalkonium, and irgasan was also established by agar dilution. 19
Clinical data
Data on age, gender, sample type, antibiotic treatment of the patient, along with history of travel and/or hospitalization within one year prior the isolation of the strains were obtained from databases of the respective laboratories.
Molecular typing
The XbaI-digested pulsed-field gel electrophoresis pattern and multi-locus sequence type of the isolates was established by standard methods.4,20
Determination of the NDM allele
The immediate surroundings of the blaNDM genes were amplified, and the amplicons were subjected to direct sequencing. 2
Detection of antibiotic resistance genes
The presence of the resistance genes (blaTEM, blaCTX-M, blaSHV, blaPER, blaAmpC, blaNDM, blaOXA-48-like, blaKPC, blaVIM, blaIMP, armA, rmtA, rmtB, rmtC, rmtD, rmtE, rmtF, qnrS, qepA, aac6-1b-cr) was established as described.2,21 The specific alleles of beta-lactamase genes were determined by direct sequencing of the respective amplicons on a 3130X genetic analyzer (Applied Biosystems). Genes implicated in biocide resistance (emrE, qacΔE, qacF, qacE, mdfA, ydgE, ydgF, sugE(c), sugE(p), cepA) were also detected by PCR. 19 The presence of the virulence factor genes (cdt, hlyA, papC, papA, papEF, fimH, PAI, fyuA, bmaE, sfa/foc DE, iutA, traT, focG, cvaC, gafD, sfaS, cnf1, afa/dra, nfaE, rfc, ibe A, iha, upaH, upaB, Kps MTII, K5, K1, Kps MTIII, sat, ireA, iroN, usp) was tested as described. 22
The phylogenetic group of isolates was determined by multiplex PCR. 23
Plasmid analysis
Mating out assays were performed with the four clinical isolates by using an azid-resistant derivative of rifampicin-resistant E. coli J53 (J53RAZ) as recipient. Transconjugants were selected on Tryptic Soy Agar containing 8 mg/L ceftazidime and 100 mg/L azide. 21 In case of unsuccessful conjugation, or conjugal transfer of more than one episome, the plasmid content of the wild-type strain was purified by using the Plasmid Maxi Prep kit (Qiagen, Germany) and the plasmid DNA was transferred into competent E. coli DH5α by heat shock transformation. 21 Tryptic Soy Agar containing 8 mg/L ceftazidime was used for selection of transformants harboring the blaNDM-7-carrying plasmid. Plasmids from single NDM-plasmid-carrying transconjugants or transformants were purified by using the Plasmid Maxi Prep kit (Qiagen, Germany). The complete DNA sequence of the pOM26-NDM-7 plasmid was obtained by using the 454-Genome Sequencer FLX procedure (Roche Diagnostic, Monza, Milan). The other three blaNDM-7-bearing plasmids were submitted for commercial next-generation sequencing by Illumina MiSeq performed at the CCIB DNA Core Facility in Massachusetts General Hospital (Cambridge, MA). The gaps between the contigs assembled were closed by PCR and direct sequencing of the amplicons on a 3130X genetic analyzer (Applied Biosystems). The incompatibility type of the plasmids was defined by using PlasmidFinder. 24 Sequences were annotated by using Sequin (www.ncbi.nlm.nih.gov/Sequin) and submitted to GenBank.
Results
Of the 157 NDM-positive strains, 144 (91.7%) carried NDM-1, 9 (5.7%) carried NDM-5, and the NDM-7 allele was found in 4 (2.5%) isolates. The clinical data of these latter four isolates are summarized in Table 1. All four NDM-7 producer E. coli were isolated in 2012 in different hospitals from Kuwait (KW53T), Oman (OM26), and Dubai (ABC218) and Abu Dhabi (ABC133) Emirates of the UAE, respectively. Two of the patients travelled to India one year prior the isolation of the NDM-7 producer E. coli. Before the isolation of the carbapenemase producer organism, three patients received broad-spectrum antibiotic therapy, which in one case included carbapenem as well.
During the year before the isolation of the strain.
F, female; KSA, Kingdom of Saudi Arabia; M, male; UAE, United Arab Emirates.
The isolates were uniformly susceptible to tigecycline, colistin, and fosfomycin, variably susceptible to aminoglycosides, co-trimoxasole, chloramphenicol, and tetracycline, and all were resistant to ciprofloxacin and all beta-lactams tested, including aztreonam (Table 2).
AMK, amikacin; AZT, aztreonam; BAC, benzalkonium; CAZ, ceftazidime; CHL, chloramphenicol; CHX, chlorhexidine; CIP, ciprofloxacin; COL, colistin, CTX, cefotaxime; ETP, ertapenem; FOS, fosfomycin; GEN, gentamicin; IMP, imipenem; IRG, irgasan; KAN, kanamycin; MEM, meropenem; MIC, minimum inhibitory concentration; NDM, New Delhi metallo-beta lactamase; R, Recipient; STR, streptomycin; TC, Transconjugant; TET, tetracycline; TF, Transformant; TGC, tigecycline; TMP-SMX, trimethoprim-sulphamethoxazole; W, Wild type.
Genotyping results (Fig. 1 and Table 3) showed that the strains were clonally unrelated and did not belong to virulent extra-intestinal E. coli phylogenetic groups, although all belonged to E. coli clonal complexes that were associated with multi-drug resistance. 25 All isolates harbored an AmpC cephalosporinase and aac6′-1b-cr. Moreover, three of them possessed blaCTX-M-15 and the Kuwaiti strain (KW53T) also carried 16S methylase gene rmtB.

PFGE comparison of the NDM-7 producer Escherichia coli isolates from the Arabian Peninsula. NDM, New Delhi metallo-beta lactamase. PFGE, pulsed field gel electrophoresis.
Genes printed in bold are co-located on the NDM-7 plasmid.
CC, clonal complex; MLST, Multi-locus sequence typing; NA, not available; ND, not determined; NT, not typable; Phyl, phylogenetic group; RT, replicon type; ST, sequence type.
Although all strains carried genes that were associated with disinfectant resistance 19 (Table 3), no increase in MIC to the disinfectants tested was detected compared with the respective E. coli K12 recipient strains (Table 2), in which we also detected certain disinfectant resistance genes: in E. coli J53RAZ, emrE, mdfA, ydgE, ydgF, and sugE(c) and in E. coli DH5α, mdfA, ydgE, ydgF, and sugE(c).
From three isolates, plasmids carrying the blaNDM-7 gene were successfully transferred into E. coli J53RAZ by conjugation. Conjugation failed for ABC133, and pABC133-NDM was transferred by transformation into E. coli DH5α. The KW53T transconjugants contained multiple plasmids. Therefore, pKW53T-NDM was also transferred by transformation into E. coli DH5α. Plasmid typing showed that the four NDM-7 plasmids belonged to IncX3 incompatibility type. The complete sequences of the plasmids were determined. Beyond blaNDM-7, the plasmids did not carry any other resistance gene. The immediate surrounding of the blaNDM-7 gene was identical in all four plasmids. The largest of the four plasmids, pKW53T-NDM-7 of 46,161 bp size, was almost identical to the blaNDM-7 plasmid of EcN01 (GenBank CP012990), with the exception of a single nucleotide change at position 13857 (T to G), resulting in an Ile106 to Leu change in the transposase of IS26. However, compared with pKW53T-NDM-7, pOM26-1 lacked the 5′ end of the ISAba125 located downstream of IS5 in pKW53T-NDM-7. In pABC218-NDM-7, a 11,758 bp deletion was observed, including the 5′ end of the ISAba125, a Tn3 and another transposase, the mpr gene, genes coding for a hypothetical protein, a H-NS histone family protein, a DNA topoisomerase III, and the 3′ end of an ATPase gene, flanked by two 4-bp repeats (CCTT). A similar, although smaller (9,867 bp), deletion, flanked by the same repeats, but only truncating the topB gene, was observed in the non-conjugative pABC133-NDM-7. In this plasmid, further changes affecting the conjugative apparatus were detected: The pilX8 gene was disrupted by an IS1 element, and the pilX3 gene suffered a single base deletion (Fig. 2).
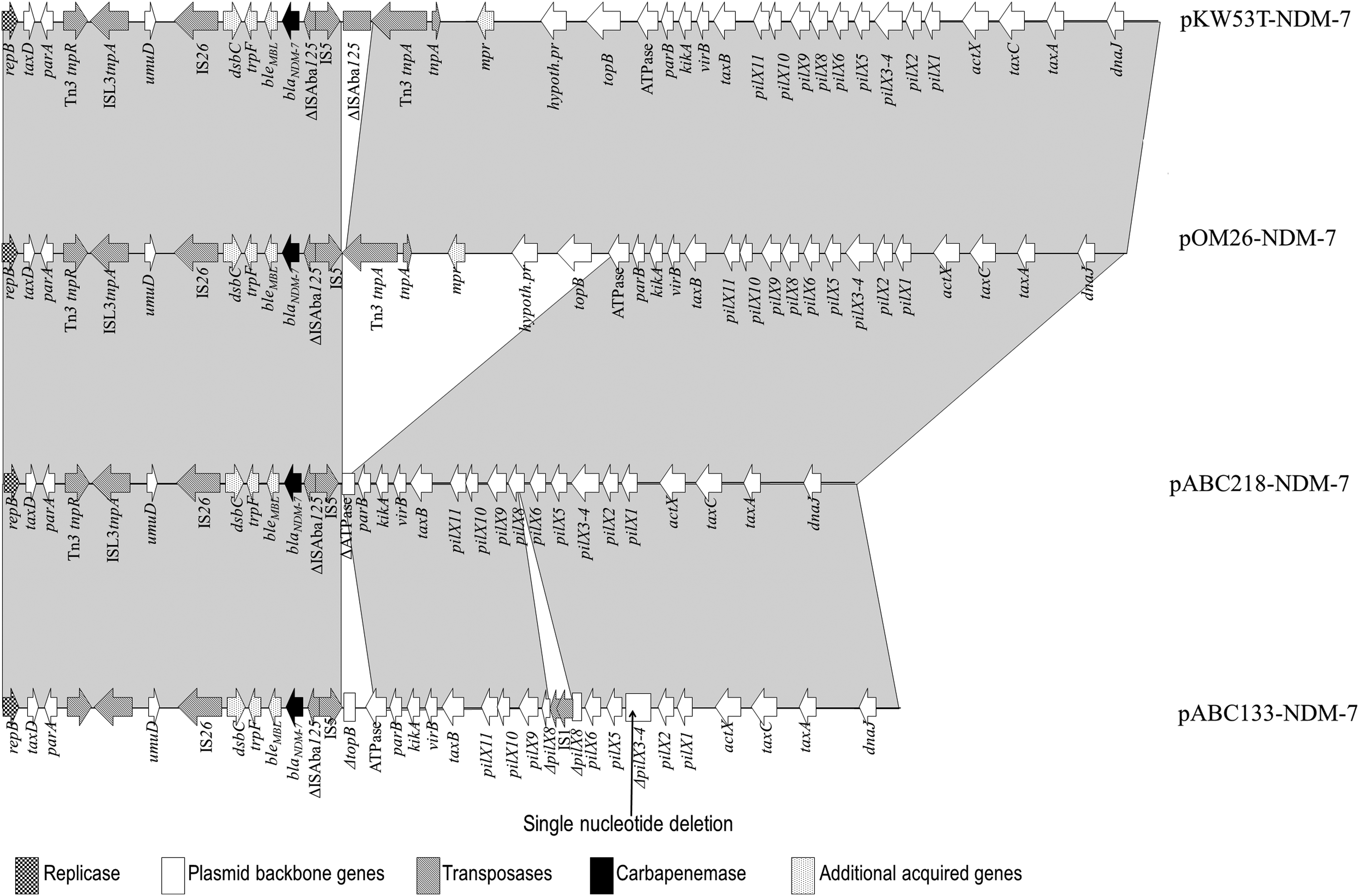
Comparison of the NDM-7-carrying IncX3 plasmids from the Arabian Peninsula.
Discussion
Although NDM-7 carbapenemase is more efficient in hydrolyzing imipenem,
9
and it shows a global distribution, yet it is considered a rare variant of NDM that is mostly linked to travel to India.
16
According to our study, the NDM-7 allele in the Middle East is also comparatively rare (2.5% of all NDM producers in our collection). In this region, NDM-7-producing organisms were not exclusively related to travel to the Indian subcontinent. Unlike in Spain,
14
these strains carrying the NDM-7 allele were not clonally related. Importantly, however, all isolates belonged to E. coli clonal complexes that were associated with multi-drug resistance.
26
Indeed, all four isolates described possessed the plasmid
The MIC values of benzalkonium, chlorhexidine of the wild-type strains considerably exceeded that of E. coli DH5α, but not of E. coli J53RAZ. Although an investigation of the molecular background of this observation was beyond the scope of this study, we surmise that this could be due to the absence of qacEΔ1 or emrE, encoding small multidrug resistance family (SMF) proteins 19 in DH5α, while present in the carbepenemase-producing wild-type strains and in E. coli J53RAZ (Table 3). Nevertheless, neither the pNDM-7 transformants nor the transconjugants exhibited any increased resistance to any of the disinfectants, in line with the fact that none of these plasmids carried any disinfectant resistance genes (Tables 2 and 3 and Fig. 2).
The comparison of our current data on NDM-7-producing E. coli with those reported earlier (Table 3) shows that the blaNDM-7 is a plasmid-encoded gene that is carried mostly on self-conjugative, 46 to >100 kb-sized plasmids of the IncX3, IncF, and IncA/C incompatibility groups.6,7,9,13,15,16,27 IncX3 incompatibility type plasmids associated with blaNDM-7 were reported earlier from Germany, 6 Canada, 15 and China. 27 Of particular interest is that in Canada, a plasmid almost identical to the one in the Kuwaiti isolate, pKW53T-NDM-7 was detected in several enterobacterial species. 15 Interestingly, in the Omani and in the two UAE isolates, the NDM-7 plasmids were also related to the pKW53T-NDM-7 plasmid but lacked mobile genetic elements or even the topoisomerase III gene (Fig. 2).
Importantly, three of the four plasmids were transferable by conjugation. In case of pABC133-NDM-7, the single nucleotide deletion and the IS1 insertion in the conjugation machinery explain the lack of mobility. We surmise that in this patient the carbapenem therapy administered had a role in maintaining the carbapenemase gene in ABC133 sputum isolate. Nevertheless, apparently highly similar IncX3-type plasmids play an important role in the spread of the currently rare NDM-7 variant in the Arabian Peninsula, as well as in other parts of the world. This association of blaNDM-7 with the IncX3-type plasmid is particularly worrisome, as this type of plasmid has been shown to spread NDM-, KPC-, and OXA-type carbapenemases in various species of Enterobacteriaceae worldwide with a high efficacy.2,4,21,28–32
Footnotes
Acknowledgments
This work was supported by the United Arab Emirates University grants NRF-31M085 and CMHS-31M251 awarded to AS and UPAR-25143 awarded to TP.
Disclosure Statement
No competing financial interests exist.