
Abstract
In our earlier in vitro and in vivo studies, synergistic effects were observed when itraconazole or voriconazole were combined with tetrandrine (TET) against Aspergillus fumigatus, and the synergistic mechanism was related to inhibition of the drug efflux pump. Posaconazole (PCZ) is a broad-spectrum triazole antifungal agent used for the treatment of diverse fungal infections, including aspergillosis and candidiasis. Herein, the antifungal effects of TET are further investigated in vitro and in vivo alone or combined with PCZ against 20 clinical isolates of A. fumigatus. We found that the minimal inhibitory concentrations (MICs) of PCZ were decreased one- to twofold and three- to fivefold across a series of concentration gradients in vitro in presence of TET. Time-killing curves revealed that the synergy was dependent on TET and PCZ concentrations as well as incubation time. The combination could further downregulate the expression of MDR2, MDR3, MDR4, and ATRF in PCZ-resistant strain, however, it has subtle effects on TET-synergized mechanism. In addition, TET in combination with PCZ significantly prolonged mice survival time and reduced kidney and brain tissue burdens in vivo. Our data in vitro and in vivo demonstrate that TET is an effective synergist with azoles against A. fumigates.
Introduction
I
Our earlier studies have demonstrated that TET acted synergistically with fluconazole against A. fumigatus and Candida albicans in vitro and in vivo.5,6 The synergistic mechanism of action may be related to inhibition of the drug efflux pump and downregulation of the expression of drug efflux pump genes.4,7 The present study is to further investigate the synergistic effect and mechanism of TET with PCZ against the invasive filamentous A. fumigatus.
Materials and Methods
Organisms and sources
A. fumigatus CMCC (F) A.1a was obtained from Institute of Dermatology, Chinese Academy of Medical Sciences (Nanjing, China). A total of 19 clinical isolates were obtained from Guangzhou General Hospital under Guangzhou Area Command (Guangzhou, China). Candida parapsilosis ATCC 22019 was used as a control isolate as recommended by the Clinical and Laboratory Standards Institute (CLSI). According to the interpretive breakpoints for PCZ (≤0.125, and ≥0.5 μg/mL), 8 A. fumigatus isolates AF-17 and AF-28 were sensitive to PCZ; AF-7, AF-8, AF-13, AF-20, AF-46, and AF-0193 were dose-dependent sensitive to PCZ; and AF-9, AF-10, AF-19, AF-21, AF-23, AF-24, AF-26, AF-27, AF-29, AF-30, AF-31, and AF-CMCC were resistant to PCZ.
All isolates were retrieved from frozen glycerol stocks and subcultured on potato dextrose agar (PDA) at 37°C (Gibco) 5 days before use. For all experiments, conidial suspensions were dislodged from hyphal mats by dispersal in 0.9% saline with 0.05% Tween 80 (Sigma-Aldrich) and filtered through sterile gauze to remove hyphal fragments, as described elsewhere. 8
Antifungal susceptibility testing
Susceptibility testing was performed according to the checkerboard microdilution procedure based on the Clinical and Laboratory Standards Institute (CLSI) M38-A2 document 9 and previously published protocol. 10
In brief, twofold serial dilutions of drugs were prepared in RPMI 1640 medium at four times the final desired concentration. Serial dilutions of 50 μl of PCZ (final concentration ranged from 0.015625 to 8 μg/ml) were added to the wells of 96-well flat-bottomed microtiter plate in columns 2–12, while serial dilutions of 50 μl of TET (final concentration ranged from 16 to 512 μg/ml) were added to the wells of rows G–A. Next, 50 μl of RPMI 1640 was added to the wells of column 1 and row H, followed by the addition of 100 μl of conidia suspensions (final concentration 5 × 103 cells/ml) to all wells except well A12, to which 100 μl of RPMI 1640 was added.
Next, the plates were incubated at 37°C for 48 h. Visual reading of minimal inhibitory concentrations (MICs) was performed, and OD490 was measured by microplate reader. All experiments were performed in triplicate on two separate days.
Drug interaction interpretation
To evaluate the in vitro effects of combined PCZ and TET, the data obtained from the checkerboard tests were analyzed by nonparametric models based on the Loewe additivity model (LA).
The fractional inhibitory concentration index (FICI) was defined by the equation: ∑ FIC = FICTET+FICPCZ = MICTET+PCZ/MICTET+MICPCZ+TET/MICPCZ, where MICTET and MICPCZ represent the MICs of TET and PCZ when tested alone, and MICTET+PCZ and MICPCZ+TET represent the MICs of TET in combination with PCZ. The results were categorized according to the previously described method using the following rules: synergism (FICI ≤ 0.5), indifference (0.5 < FICI ≤ 4), and antagonism (FICI > 4.0).
Time-kill curves
To further investigate the effect of concentration and exposure time on the activity of PCZ alone or in combination with TET, time-kill curves were performed using an adaptation of previously published methods.11,12
Cell viability (i.e., the OD) versus time was measured using the XTT assay for the aforementioned fungal biomass-associated metabolic activity. Defined concentrations of 1 × 105 cells/ml conidia of both the PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC were added to 96-well microtiter plates and incubated with defined concentrations of each compound (PCZ, 0.5, 1, and 2 μg/ml; TET, 8, 16, and 32 μg/ml) at 37°C. At each time point (0, 1, 2, 4, 8, 12, and 24 h) after incubation, 100 μl of XTT–menadione solution was added. The plates were then incubated in the dark for 2 h at 37°C. Colorimetric changes of the XTT reduction product (formazan) were measured at 490 nm with a microplate reader. All experiments were performed in triplicate on 2 separate days.
In vivo experiments
Female BALB/c mice (18–22 mg; Guangdong Medical Laboratory Animal Center, Foshan, Guangdong, China) were used for all experiments. The mice were housed in filter-top cages with free access to food and water.
Mice were immunosuppressed with cyclophosphamide (200 mg/kg) administered intravenously on day 3 before fungal challenge. 13 Mice were challenged on day 0 via the lateral vein with conidia suspensions containing 7.5 × 104 cells of the PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC. Groups of 10 mice were randomly selected for each strain and treatment.
The in vivo dosing regimens, including dose level, dose interval, and treatment duration were chosen from previous studies.13–16 On day 1 postinfection, mice were administered PCZ (25/50 mg/kg/d) orally and/or TET (40/60 mg/kg/d) intraperitoneally for 7 days. Controls received saline orally or intraperitoneally. For survival studies, mice were checked daily for 22 days after infection. For tissue burden studies, mice were sacrificed on day 5 postinfection, and kidneys and brains (the main target organs) were aseptically removed, weighed, and homogenized. 17 Serial 10-fold dilutions of the tissue homogenates were plated on PDA, and colonies were counted after 2 days of incubation.
Mortality was plotted with Kaplan-Meier survival curves. The tissue fungal burdens are quantitatively ranked at the log scale and compared using the Mann–Whitney test or the Kruskal–Wallis H-test in cases where use of the Mann–Whitney test is not possible. All animal experiments were performed in accordance with the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication No. 85–23, Revised 1996) and were carried out with permission from the Ethical Committee of Institute of Zoonosis, Jinan University, Guangdong, China.
Quantitative real-time PCR
Approximately 1 × 105 cells/ml of the PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC were incubated with shaking (200 rpm) at 37°C for 16 h. Subsequently, subinhibitory concentrations of PCZ and TET were added to the media and incubated for additional 8 h under the same growth conditions.
Total RNA was extracted and purified using the RNeasy Plant and filamentous fungi Mini Kit (Qiagen, Inc.) and Turbo DNase I (Ambion, Inc.). cDNA was synthesized using the PrimeScript™ RT Master Mix (TaKaRa Dalian). All Quantitative real-time PCR (RT-PCR) were performed in triplicate using SYBR Green qPCR SuperMix (Invitrogen). The PCR were carried out with a denaturation step at 95°C for 2 min, followed by 40 cycles of amplification and quantification at 95°C for 15 sec, 60°C for 32 sec, and 72°C for 15 sec and extension at 72°C for 1 min. The results were analyzed using the 2−ΔΔCT method, and statistical significance was determined using the one-way ANOVA test. The primer sequences of efflux pump genes and reference genes used in the RT-PCR were designed as previously described. 18
Results
Antifungal susceptibility testing
The MICs of PCZ alone and in combination with TET against A. fumigatus are shown in Table 1. In the case of TET in combination with PCZ, the overall MICs of PCZ were reduced from 0.0625–1 μg/ml to 0.03125–0.25 μg/ml, and the MICs of TET were reduced from 256–512 μg/ml to 32–128 μg/ml in 20 strains with a 90% reduction of growth. The results of the drug interaction are also interpreted by the FICI value in Table 2. When PCZ was administered in combination with TET, synergistic interactions were observed in 85% (17 of 20) of the isolates. These results demonstrate that the in vitro activities of TET act synergistically with PCZ against A. fumigatus as well as itraconazole or voriconazole.
MIC, minimal inhibitory concentration; TET, tetrandrine; PCZ, posaconazole.
For the FICI model, synergy was defined as FICI ≤0.5, antagonism was defined as FICI > 4.0, and independence was defined as FICI between 0.5 and 4 (i.e., no interaction).
FICI, fractional inhibitory concentration index; INT, interpretation; SYN, synergism; IND, independence.
Time-killing curves
A. fumigatus cells were treated with PCZ alone and in combination with TET at 0.5-, 1-, or 2-fold higher than the plasma concentrations to validate the results of the checkerboard microdilution analysis. The time-killing curves are shown in Fig. 1. The percentage of viability in both strains show a pronounced reduction after treatment with TET in combination with PCZ when compared with PCZ alone. Overall, the results show that TET strikingly enhances the antifungal effects of PCZ against A. fumigatus because of reduced cell viability over a shorter time period.
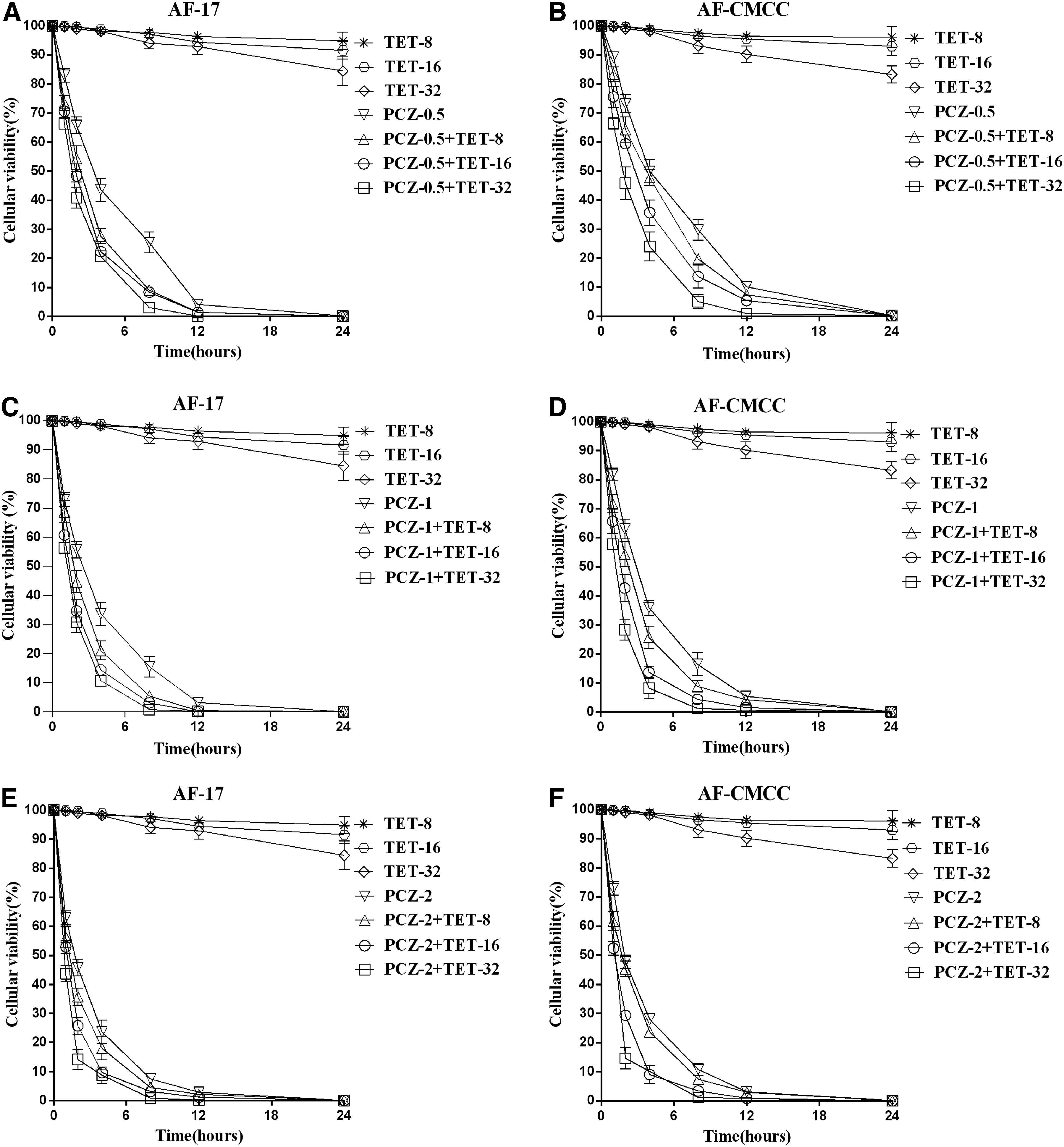
Time-kill curves of TET (8, 16, and 32 μg/ml) alone and in combination with PCZ at 0.5 μg/ml
Quantitative real-time PCR
To determine whether the synergistic mechanism of TET with PCZ against A. fumigatus is also involved in downregulation of efflux pump genes, we used RT-PCR to analyze efflux pump genes in both the PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC.
First, the inherent mRNA levels of MDR1, MDR2, MDR3, MDR4, and ATRF were measured and compared between PCZ-resistant strain AF-CMCC and PCZ-sensitive strain AF17. As shown in Fig. 2A, all MDR-related efflux genes tested here are upregulated in PCZ-resistant strain AF-CMCC except MDR1. Intriguingly, this MDR1 expression can be effectively induced by PCZ alone in PCZ-sensitive (∼1.75-fold) and resistant strains (∼3.0-fold) in Fig. 2B, C. A similar inducible MDR1 expression is also seen with TET treatment in PCZ-sensitive strain (Fig. 2B) at smaller scale. This upregulated MDR1 sustains about same level of PCZ treated alone in the course of two drugs treatment, suggesting that MDR1 is highly responsible to PCZ.

Relative expression levels of the drug efflux pump genes in PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC
Similar to MDR1, the MDR3 is also upregulated with each drug treated alone with a slightly higher expression in PCZ-resistant strain than sensitive strain. Further evaluation of MDR3 expression showed that this gene can be repressed while PCZ and TET are used together, but the repressed expression of MDR3 is actually slightly higher than the one observed in control cells. On the contrary, while each agent alone has no effect on expression of MDR2, MDR4, or ATRF, the mRNA level of each gene is effectively repressed in both PCZ-resistant and PCZ-sensitive strains in presence of PCZ and TET combination (p < 0.05).
These results reveal that PCZ-resistant mechanism was more relative to upregulation of the efflux pump genes MDR2, MDR3, MDR4, and ATRF (Fig. 2A). However, the observed synergism of TET with PCZ in both PCZ-resistance and sensitive strains suggests that the synergistic mechanism has very little, if any, to do with inhibition of these efflux genes in A. fumigatus.
In vivo experiments
In vivo, the survival curves demonstrated that both the PCZ-sensitive strain AF17 and PCZ-resistant strain AF-CMCC caused acute and lethal infections in mice. When treated infected mice with a high dose of PCZ (50 mg/kg/d) alone, we observed 70% and 60% survival rates for PCZ-sensitive and PCZ-resistant strains, respectively, which were higher than the control mice (p < 0.05). In the case of TET (60 mg/kg/d) in combination with a low dose of PCZ (25 mg/kg/d), the survival rates were around 90% and 80%, respectively, which significantly improved the survival rates when compared with PCZ (25 mg/kg/d) alone (p < 0.05) (Fig. 3).

Survival curves of mice infected systemically with PCZ-sensitive strain AF17
In agreement of survival rates, while treatment with PCZ at various concentrations significantly reduced kidney and brain tissue burdens (p < 0.05), treatment with TET (60 mg/kg/d) in combination with PCZ at various concentrations further decreased kidney and brain tissue burdens versus PCZ alone (p < 0.05) (Fig. 4).
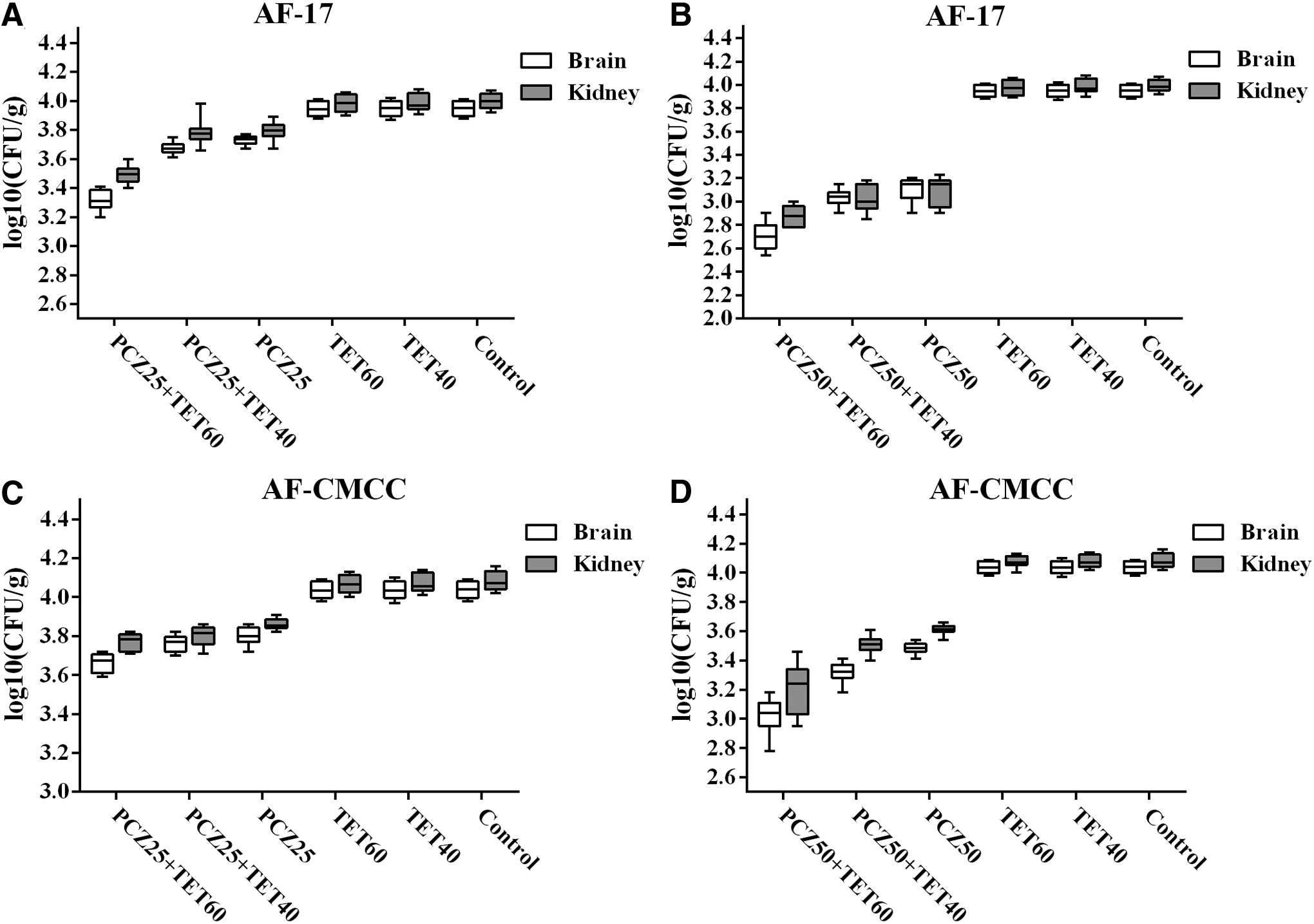
Box plot of changes in fungal tissue burdens of mice infected with PCZ-sensitive strain AF17
Discussion
Our results showed that the MICs of PCZ in combination with TET were decreased one- to twofold and three- to fivefold across a series of concentration gradients in vitro compared with PCZ and TET alone, respectively. Time-killing curves further revealed that the synergy was dependent on PCZ and TET concentrations and incubation time. Furthermore, PCZ in combination with TET significantly prolonged mice survival time and reduced kidney and brain tissue burdens in vivo versus PCZ alone. These results are in agreement with the findings from the previous studies, of which the synergistic effect of PCZ with immunosuppressants (FK506, cyclosporine, and rapamycin) or antifungal agents (amphotericin B, voriconazole, and caspofungin) have been documented against A. fumigatus.19,20
The fungistatic nature and prolonged use of azole to treat fungal infections, however, have been noted by a possibility to promote the selection of drug-resistant fungal strains. In clinical settings, combination therapy has become a potential alternative to treat invasive fungal infections as it can improve clinical efficacy of existing drugs such as azole by reducing their side effects to host when lower doses are used. 20 Our previous studies demonstrated that TET acted synergistically with fluconazole against filamentous A. fumigatus and yeast Candida albicans in vitro and in vivo.7,21,22
The azole resistance mechanisms of A. fumigatus were mainly mediated by overexpression of the drug efflux pump and its associated genes.18,23–25 Morschhauser reported that overexpression of the drug efflux pump and its associated genes can promote A. fumigatus to develop azole resistance. 26 To date, four ABC transporters (MDR1, MDR2, MDR4, and ATRF) and one major facilitator (MDR3) of A. fumigatus have been cloned, and our previous studies demonstrated that the drug efflux pumps in itraconazole/voriconazole-resistant strain were higher than that in azole-sensitive strain. 22 Also, Zhang et al. 21 demonstrated that the synergistic mechanism of TET with fluconazole against C. albicans was due to inhibition of the drug efflux pump.
However, RT-PCR results in this study suggest that the molecular mechanism of the observed TET synergistic effects with PCZ in A. fumigatus cannot be simply explained by suppression of these overexpressed efflux pumps, since MDR2, MDR3, MDR4, and ATRF are repressed by PCZ, TET, and combination of two compounds in both PCZ-resistant and sensitive strains. In contrast to synergized effects of TET on suppression of these efflux genes, the MDR1 level can be induced in both strains with PCZ treatment, which cannot be withdrawn by TET addition. The expression of MDR3 is further stimulated with PCZ treatment in resistant strain that can be substantially repressed with combination treatment even though absolute levels of MDR3 in both strains are still above control baseline.
These findings suggest that TET and PCZ combination can further reduce these MDR efflux pumps except MDR1. As MDR3 is a major contributor of resistance as we mentioned above, less inducible roles of TET on MDR1 and MDR3 may benefit A. fumigatus cells from azole resistance developed with high-dose azole treatment.
We suspect that two possibilities may explain MDR1 and MDR3 behaviors. First, the azole resistance mechanisms of the A. fumigatus are not mediated by overexpression of MDR1 that can be supported by this study and others. Tobin et al. 25 also failed to detect an overexpression of MDR1 in azole resistant A. fumigatus. Second, mode of TET has no action on PCZ-inducible efflux pumps such as MDR1 or MDR3, but targets on different efflux pumps. To draw the conclusion, the time courses of MDR1 and MDR3 expression in resistant A. fumigatus and their implications on resistance require further exploration. Also, the dosage of TET on these genes expression needs to be optimized to rule out its roles on MDR3-mediated resistance. We believe that a study at larger scale regarding to these matters will help us fully explain the synergistic mechanism of TET.
In summary, TET can act synergistically with PCZ against A. fumigatus in vitro and vivo, and part of their synergistic mechanisms may be due to the downregulation of some drug efflux pump genes such as MDR2, MDR4, and ATRF.
Footnotes
Acknowledgments
We thank Institute of Dermatology, Chinese Academy of Medical Sciences (Nanjing, China) and Guangzhou General Hospital under Guangzhou Area Command (Guangzhou, China) for generously providing A. fumigatus isolates. This work was supported by the National Natural Science Foundation of China (81471995/81171542).
Authors' Contributions
S.L., Y.S., and H.Z. designed the research; S.L., Y.S., L.J., Y.Z., and H.G. performed the experiments. S.L., Y.S., L.J., Y.Z., H.G., D.L., K.Z., and H.Z. analyzed data and wrote the article. All authors read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.