
Abstract
Multidrug-resistant Escherichia coli can contaminate food meat during processing and cause human infection. Phenotypic and genotypic characterization of the antimicrobial resistance were conducted for 45 multidrug-resistant E. coli isolates from 208 samples of beef carcasses. The mechanisms of resistance were evaluated using polymerase chain reaction and sequencing methods, and the clonal relationship among isolates was evaluated using multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE). Different variants of bla, tet, flo, dfrA, and aadA genes were detected in most of the strains resistant to β-lactam, tetracycline, chloramphenicol, sulfonamides, and aminoglycosides, respectively. Extended-spectrum β-lactamase (ESBL)-producing E. coli was found in 42.2% of the 45 E. coli isolates and the most commonly detected ESBL genotypes were CTX-M group 1 and 9. Class 1 integrons with nine different arrangements of gene cassettes were present in 28 of 45 E. coli isolates. Twenty-nine PFGE groups and 24 MLST types were identified in their clonal structure. This study revealed that E. coli isolates from beef contained high diversity of antimicrobial resistance genes, integrons, and genotypes. These results highlighted the role of beef meat as a potential source for multidrug-resistant E. coli strains and the need for controlling beef safety.
Introduction
M
The increasing prevalence of extended-spectrum β-lactamase-producing E. coli (ESBL-E. coli) has brought great trouble to the physicians in treating infectious diseases. Plasmid-mediated AmpC β-lactamase-producing E. coli strains are also an emerging problem in human and veterinary medicine because of the mobile genetic element-mediated transmission of resistance genes. 4
The risk for humans to acquire antimicrobial-resistant bacteria from food animals is much lower than from receiving antimicrobial therapy or hospital-acquired infection. However, resistant bacteria of food animals still can transmit to humans by direct contact in farm and slaughterhouse workers and veterinarians, and consumption of raw milk or not fully cooked meat. 3 In addition, a previous study suggested that a large proportion of multidrug-resistant E. coli isolates causing blood stream infections in the people may come from food animals, especially poultry and pigs. 5 It is now generally accepted that the key factors for increased resistance in bacteria can be traced to the misuse or overuse of antibiotics. Therefore, the antibiotic use in animal husbandry has become a subject of major global concern.
Several studies have described that mobile and mobilizable genetic elements, such as integrons, play an important role in the carriage and dissemination of antibiotic resistance. 6 According to the sequence of integrase genes, the integrons are classified into a few groups or classes, of which class 1 integron has received the most attention because it is most frequently found in Gram-negative isolates of humans and various animal hosts. 7 More than 100 different gene cassettes carried by class 1 integrons have been found. 6 The cassettes with various array patterns have been reported in most bacterial species from different world regions, and the variations and evolution of cassettes carried by integrons may still be created. The dissemination of class 1 integrons usually is related to their location in plasmids and/or transposons that enhance the spread of resistance genes.
The prevalence and contamination levels in food production animals, particularly in cattle, have been monitored in western countries.1,5 However, the current epidemiology of antibiotic resistance patterns of E. coli from beef meat in Taiwan is poorly understood. High percentages of antimicrobial resistance and transmitter factors among common pathogenic bacteria in humans have been reported in Taiwan. 8 Therefore, the objective of this study was to provide information of prevalence of ESBL positivity and the correlation between antimicrobial resistance and integrons in different E. coli isolated from various beef carcasses in Taiwan.
Materials and Methods
Sample collection and E. coli isolation
In this study, E. coli isolates from beef carcass surface were collected from two different abattoirs (A, B) located in central and southern Taiwan. Samples were collected and analyzed following the U.S. Department of Agriculture Food Safety and Inspection Service standardized method. 9 All isolates from fresh beef samples were sent to the laboratory at 4°C. Here, the samples were immediately processed by both culture and identification. The isolates were identified on the basis of routine microbiologic methods, and species identification was confirmed using the VITEK system (BioMerieux Vitek, Inc., Hazelwood, MO). E. coli J53 (sodium azide resistant) was used as a recipient in the conjugation experiment.
Antimicrobial susceptibility testing and confirmatory tests of ESBL phenotype
Susceptibilities to 17 antimicrobial agents were determined by disc diffusion method according to Clinical and Laboratory Standards Institute (CLSI) guidelines. 10 Minimal inhibitory concentration (MIC) testing, including cefotaxime, cefotaxime/clavulanic acid, ceftazidime, ceftazidime/clavulanic acid, and imipenem, was undertaken to confirm ESBL production as described previously 11 and CLSI guidelines. 10 Quality control was assured by testing E. coli ATCC 25922 and Klebsiella pneumoniae 700603.
Identification of antimicrobial resistance genes
Total DNA of E. coli isolates was prepared using a Wizard genomic DNA preparation kit (Promega, Madison, WI) with 0.5 mg/ml of lysostaphin and 0.3 mg/ml of RNase for the lysis step. Polymerase chain reaction (PCR) amplification was used to identify the presence of blaTEM, blaSHV, blaCTX-M, blaAmpC, insertion sequence, plasmid-mediated quinolone resistance (PMQR) genes (qnrA, qnrB, qnrC, qnrS, qepA, and aac(6')-Ib), and quinolone resistance-determining regions of the gyrA and parC genes. Primers for blaCTX-M family, 12 and group-specific primers for blaCTX-M-1 group 13 and blaCTX-M-9 group, 14 the most common blaCTX-M genes of E. coli in Taiwan, 11 were used to screen the presence of blaCTX-M.
The plasmid DNA was extracted using a commercial plasmid DNA purification kit (Genemark, Taichung, Taiwan). The oligonucleotide primer sets specific for the antibiotic resistance genes and integrons in the PCR were as previously described11–19 and listed in Table 1. The PCR was performed using a conditioned program: denaturation at 94°C for 5 min, followed by 30 cycles of 30 sec at 94°C, annealing for 40 sec at 55°C, and extension at 72°C for 40 sec for the class 1 integrase gene and 4 min for the variable region. The resulting fragments were separated by electrophoresis using a 1% agarose gel. Both DNA strands of the PCR product were sequenced by the Mission Biotech, Taiwan. Sequence analyses were performed online at the National Center for Biotechnology Information website (www.ncbi.nlm.nih.gov).
Transfer of resistance determinants
Transfer of the β-lactam resistance element from study E. coli isolates to E. coli J53 (Azir) was conducted by solid mating assays. Transconjugants were selected on trypticase soy agar plates (Becton, Dickinson and Company, Sparks, MD) supplemented with 100 μg/ml sodium azide and 2 μg/ml cefotaxime (Sigma, St Louis, MO). Plasmids were extracted from E. coli J53 transconjugants using a Plasmid Miniprep Purification Kit (Amersham Biosciences, Uppsala, Sweden).
Clonal relatedness
Clonal relatedness among E. coli isolates was assessed by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). Whole chromosomal DNA of beef isolates embedded in agarose gel plugs (FMC Bioproducts, Philadelphia, PA) was treated with proteinase K and restriction endonuclease XbaI according to the manufacturer's recommendations (New England Biolabs, Ipswich, MA). The restriction fragments were separated by electrophoresis in a CHEF-DR III apparatus (Bio-Rad Laboratories, Hercules, CA) for 24 hr. XbaI-digested DNAs from Salmonella enterica subsp. enterica serotype Braenderup H9812 were used as molecular size markers. PFGE running conditions were as follows: 1.0% SeaKem LE agarose, 14°C, 21 hr, 6 V/cm, and 2.2–54.2-sec linear switch ramp.
PFGE patterns were compared using standard published criteria. 20 PFGE patterns were also analyzed using GelCompar II 6.5 software (Applied Maths). Similarity was calculated with the Dice coefficient using parameter settings of 1.0% tolerance and 0.5% optimization. The isolates were also analyzed by MLST as described previously. 21
Results
Antimicrobial resistance profiles of E. coli isolates
For survey of antimicrobial resistance profiles, a total of 208 E. coli isolates were obtained from 94 beef samples. Ninety-six isolates (46.1%) were resistant to at least one of the antimicrobial agents tested. Those with resistance to two or more antimicrobials (multidrug resistance) were collected for this study, consisting 21.6% (45/208) unrelated isolates (one isolate from each beef sample was collected). A low resistance to ceftazidime (13%), nalidixic acid (19%), amikacin (22%), ciprofloxacin (24%), and enrofloxacin (22%) was found. Resistance to tetracycline, aminoglycosides, cephalexin, and ampicillin was the most common combination from 40 (19.2%) isolates. Table 2 shows the resistance to 17 antimicrobial agents of 45 antibiotic multidrug-resistant beef isolates.
Characterization of β-lactamase resistance genes
Among 45 multidrug-resistant isolates, a total of 42 isolates with ampicillin or cephalosporin resistance were screened by PCR for the most predominant β-lactamase determinants, TEM-, SHV-, CTX-M-, OXA-, and CMY-2. All of 42 isolates harbored a blaTEM-1 gene. Other β-lactamase genes were detected: blaCTX-M group (five isolates), blaCMY-2 (one isolate), blaOXA (two isolates), and blaCTX-M group plus blaCMY-2 (one isolate). The blaSHV enzyme was not detected in this survey.
All 26 cefotaxime-resistant and cefotaxime-intermediate isolates were further screened for ESBL production. E-test ESBL screen revealed a significant reduction (≥3 doubling dilutions) of cefotaxime MIC by the presence of clavulanic acid for 19 isolates. Of these ESBL producers, three isolates (E7, E26, and E28) exhibited high-level resistance to cefotaxime (MIC ≥128 μg/mL). Distribution of MICs for tested antimicrobial agents and their susceptibilities against 19 ESBL producers are shown in Table 3. The ESBL-producing isolates were all susceptible to imipenem (MIC, 0.125–0.25 μg/ml) and resistant to cefotaxime (MIC, 16–256 μg/ml).
MICs were determined by agar dilution method.
MIC, minimal inhibitory concentration; AMC, amoxicillin–clavulanic acid; AMP, ampicillin; CAZ, ceftazidime; CAZ/CLA, ceftazidime/clavulanic acid; CIP, ciprofloxacin; CL, cephalexin; CTX, cefotaxime; CTX/CTL, cefotaxime/clavulanic acid; C, chloramphenicol; Do, doxycycline; ENR, enrofloxacin; FFC, florfenicol; NA, nalidixic acid; S, streptomycin; SXT, sulfamethoxazole–trimethoprim.
PCRs and sequencing with primers specific for blaCTX-M and blaCMY-2 genes were performed for the 19 ESBL isolates in this study. The most common type of class A ESBLs identified was CTX-M-14 (n = 7). Genes encoding CTX-M-3 (n = 6), CTX-M-1 (n = 2), and CTX-M-9 (n = 4) were also detected. CMY-2 β-lactamases combined with genes encoding CTX-M were detected in one isolate. CTX-M-14 and CTX-M-3 were the most frequently isolated genotypes in beef at 38.9% and 33.3% of the total ESBLs, respectively. Conjugation experiments were performed for all ESBL producers, and all the transconjugants expressed the ESBL-related resistance phenotypes (data not shown). The results presented show the wide distribution of blaCTX-M on resistance plasmids of E. coli and emphasize their high potential to contribute to the efficient spread of antibiotic resistance.
Integron and gene cassettes in multidrug-resistant E. coli isolates
Of the 45 multiple antibiotic-resistant E. coli isolates tested, 28 (62%) were positive for the class 1 integrase gene (intI1). These isolates with class 1 integrons showed different resistance phenotypes and most of them were resistant to streptomycin and trimethoprim. After sequencing and RFLP analysis, nine groups of integrons with gene cassettes were detected (Table 4).
Antimicrobials: Lin, lincosamide; Tp, trimethoprim; Amp, ampicillin; Sm, streptomycin; Gen, gentamicin.
Gene cassettes that conferred resistance to streptomycin/spectinomycin (aadA1, aadA2, aadA5, and aadB) were presented in 92.6% integron-positive isolates, and resistance to trimethoprim (dfrA1, dfrA17, and dfrA12) in 77.8% of the isolates. The cassette group 7 with two amplicons, carrying dfr12-orfF-aadA2/dfr12 genes that cause streptomycin and trimethoprim resistance, was the most common one (25%, 7/28) in the integron-positive isolates. One isolate carried three amplicons with dfr12-orfF-aadA2/aadA1-blaOXA-10/dfr12 and showed aminoglycoside, trimethoprim and ampicillin resistance. Conjugation experiments showed that all the class 1 integrons in our E. coli isolates were located on conjugative plasmids.
Detection of tetracycline, chloramphenicol, and quinolone resistance genes
All of the 45 E. coli isolates demonstrated low susceptibility or full resistance to tetracycline, and these isolates were screened by PCR and sequencing for the tetracycline determinants. Of the tetracycline-resistant isolates, the tet gene was found in all of the multidrug-resistant strains, 21 isolates with tet(A) being the most prevalent gene, followed by tet(B) in 12 isolates, 3 isolates with tet(A+B), and 9 isolates with tet(M). tet(C), tet(D), and tet(E) were not detected in our study isolates. The results of our study also showed that 14.5% of the tetracycline-resistant and tetracycline-intermediate isolates contained two tet genes and 5.5% with tet(M) gene, respectively.
The presence of the cmlA and floR genes was investigated in the isolates resistant to chloramphenicol and florfenicol. Of 29 chloramphenicol and/or florfenicol-resistant and florfenicol-intermediate isolates, 17 were found to harbor floR and 3 had cmlA genes. None of E.coli isolates was carrying both the floR and cmlA genes. Nine isolates did not carry the cmlA and floR gene, implying there may be other genes for chloramphenicol and florfenicol resistance, which were not detected by the primers used in this study.
Among 27 quinolone-resistant or quinolone-intermediate isolates, 6 isolates (no. 20, 22, 28, 40, 42, and 43) had simultaneous mutations at codon 83 (serine to leucine) and 87 (aspartic acid to asparagine) in GyrA and codon 80 (serine to isoleucine) in ParC. In addition, two isolates had only one mutation at codon 83 (serine to leucine) in GyrA. Mutation at codon 84 in ParC and PMQR was not detected in these 27 isolates.
Phylogenetic groups and clonal analysis
Among the 45 strains, PFGE analysis identified 37 pulsotypes and could be grouped into 29 groups using XbaI macrorestriction endonuclease. The predominant groups were H and K with seven and six isolates, respectively. The remaining PFGE groups corresponded to one to three isolates. MLST analyses identified 25 sequence types (STs). The most common STs were ST4025 and ST10, both identified in seven isolates. One hundred percentage (7/7) ST4205 and 86% (6/7) ST10 isolates belonged to PFGE group H and group K, respectively. The two STs were all present in the A site and exhibited high correlation with PFGE patterns. The results of MLST and PFGE patterns analyses were summarized in Figure 1.
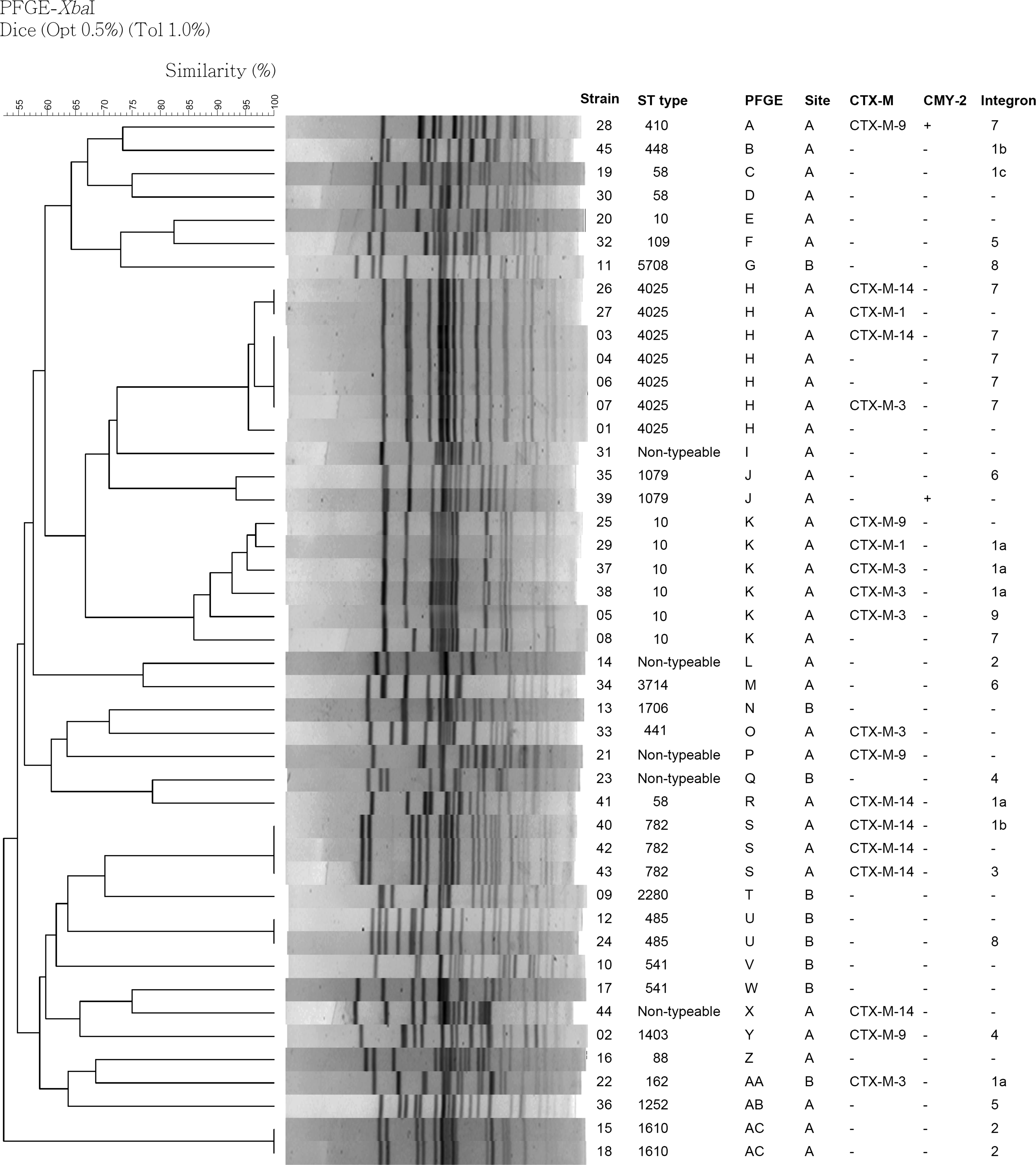
Pulsed-field gel electrophoresis patterns, sequence types, slaughter sites, CTX-M, CMY-2, and integron groups of 45 multidrug-resistant Escherichia coli isolates obtained from beef carcasses in Taiwan.
Based on the PFGE patterns, the 19 ESBL-producing isolates could be grouped into 9 groups. Most of them belonged to ST4025/PFGE group H (n = 7) and ST10/PFGE group K (n = 6). Eighteen out of 19 ESBL-producing isolates were obtained from site A. In two predominant MLST clones, five out of seven ST4205 E. coli isolates carried the gene cassette 7 and three out of seven ST10 isolates carried the gene cassette 1a.
Discussion
Isolates with resistance to various antibiotics are frequently found in livestock and humans. There is a considerable variation reported from abattoir surveys on the prevalence of antimicrobial resistance of E. coli in cattle.1,22 We aimed to study the prevalence of antimicrobial resistance of E. coli in beef carcasses sold in Taiwan and their antimicrobial resistance patterns. These results showed a resistance of E. coli in 46.1% (94/208) of beef samples, which is similar to the findings in Japan. 23 More than half of the resistant isolates showed resistance to at least two antibiotics. It indicated a high rate of antimicrobial resistance among Taiwanese beef E. coli isolates.
Among the 45 multidrug-resistant isolates, 93.3% were positive for blaTEM. The result of high prevalence of ampicillin-resistant and blaTEM-positive E. coli was consistent with the prior study and suggested that the narrow-spectrum β-lactamase TEM is common in Enterobacteriaceae of dairy cattle. 24 ESBL-producing E. coli emerged as a serious problem in human and veterinary medicine in recent years.5,8,25 We found ESBL-producing E. coli in 9.1% (19/208) of beef samples. Although ESBL-producing E. coli is a common cause of nosocomial infection in humans, ESBL-producing isolates from animal food are still rare.
Many clinical ESBL-producing E. coli isolates and ESBL-encoding genes, including SHV-5, SHV-12, CTX-M-3, and CTX-M-14, have been reported in Taiwan. 8 Although these β-lactamases have not been identified in food pathogens in Taiwan, 26 CTX-M-3 and CTX-M-14 β-lactamases were found in our beef isolates. ESBL-producing E. coli with CTX-M-1 group (CTX-M-1/15/55) and CTX-M-9 group (CTX-M-9/14/27) have been identified in food-producing animals in Western European countries and Japan.25,27 Although the CTX-M-3 is rarely reported in animals,28,29 we found six cases in this study.
As far as we know, this is the first report of E. coli in food-producing animal CTX-M-3 and CTX-M-14 ESBLs in Taiwan, emphasizing their high potential to contribute to the efficient spread as well as persistence of ESBL-producing E. coli strains outside clinical settings. All isolates that produce CTX-M exhibited resistance to cephalexin, ceftiofur, and cefotaxime; however, all of them were susceptible to imipenem.
The CMY-2 is the most common plasmid-mediated AmpC β-lactamase in food-producing animals and in meat worldwide. CMY-2-producing E. coli is the most common finding in the meat samples, but still with low prevalence in beef or cattle samples.30,31 In this study, the prevalence of CMY-2 in E. coli remained low. Among our isolates, two were detected with blaCMY-2 gene, and the overlap between ESBL/pAmpC (CTX-M-9/CMY-2) genotypes was found in only one isolate. This would suggest that plasmid-encoded AmpC cephalosporinase had limited additional effects on the ESBL population in these isolates.
The prevalence of tetracycline-, chloramphenicol-, and quinolone-resistant genes are also of our concern. Generally, resistance of Enterobacteriaceae to tetracycline is mainly mediated by tet efflux genes and found to be common in E. coli isolates from humans and animals. Screening for resistance determinants shows that the majority of tetracycline-resistant E. coli isolates from cattle harbored the tet(A) efflux gene, followed less frequently by tet(B) and tet(M). Prevalence of tet(A) is detected mainly in E. coli isolates of livestock origin, 32 while tet(B) is predominant in cattle isolates in North America to be higher than tet(A) and tet(C). 33 However, our results agree with a previous study 34 that, E. coli isolated from cattle harbor a higher prevalence of tet(A) gene.
Chloramphenicol is an antibiotic useful for the treatment of dairy farm applications, but was banned for use in all food animals in Taiwan in 2002 because of toxicity to blood and damage to bone marrow. However, florfenicol, a fluorinated synthetic analog of chloramphenicol, has continued to be legally used in veterinary in Taiwan. Concerning florfenicol/chloramphenicol, floR and cmlA resistance genes were found in our isolates, respectively. They are widespread in many bacteria, suggesting a coselection of chloramphenicol resistance with other antimicrobial-resistant phenotypes. 35 The high incidence of florfenicol resistance in the E. coli isolates analyzed in this study was in agreement with a recent study that reported a high prevalence of the floR gene among E. coli isolates. 36 They showed a similar tendency for tetracycline and chloramphenicol resistance. Simultaneous mutations in GyrA and ParC are associated with high-level quinolone resistance.37,38
Among our study isolates, simultaneous mutations in GyrA and ParC were detected in six quinolone-resistant E. coli isolates. All tested PMQR were not found in our quinolone-resistant E. coli isolates and this may reflect the low prevalence of PMQR in cattles. 39 In this study, 19 quinolone-resistant E. coli isolates did not carry PMQR genes or common mutations in GyrA and ParC. Further characterization of the resistance mechanisms to enrofloxacin/ciprofloxacin assessed in this study is needed. Taken together, these data indicate a global variation in tetracycline, chloramphenicol, and quinolone resistance rates, probably due to different practices and policies.
The increasing drug resistance of E. coli isolates in food animals may be explained by the abuse of antimicrobials and widespread presence of integrons. Class 1 integrons, most frequently found in resistant bacteria, were analyzed in our multidrug-resistant isolates. Our results revealed that resistance determinants to aminoglycosides and trimethoprim were prevalent among beef isolates. These findings were similar to another report, indicating cassettes for aminoglycoside and trimethoprim resistance were frequently detected in class 1 integrons, being common in E. coli from cattle. 40
We found that the prevalence of class 1 integrons was significantly higher in ESBL E. coli isolates from beef carcasses. The most prevalent array dfrA12-orfF-aadA2 was identified not only in single amplicon but also in two or three amplicons in this study. The array dfr12-orfF-aadA2 is also the most prevalent in different bacterial species worldwide and appeared to become stably integrated. 40
The gene cassette blaOXA-10-aadA1 has been detected in a composite transposon-located integron of E. coli. The gene cassette that contained aadB and/or blaOXA-10 in class 1 integron, is earlier also detected in E. coli (AF205943.1) and a few other bacteria such as Aeromonas spp. (JX141473.1) and Vibrio spp. (KU356480.1).The cassette array “aadB-blaOXA-10” is only reported in Pseudomonas aeruginosa (accession nos: AY536742); however, we identified this gene cassette in one E. coli beef isolate. Furthermore, the rare combination of aadA2-linF was found in five of the E. coli isolates. The array aadA2-linF has been reported in clinically isolated E. coli in Malaysia 41 and Salmonella Anatum strain isolated from poultry in the Netherlands. 42 This implicates the efficiency of transmission of various gene cassettes to different parts of the world.
ST131 is the most widespread ST of multidrug-resistant E. coli 43 ; however, ST131 E. coli was not identified in this study. Prior study results showed that the ratio of identical ST of E. coli between human–beef is significantly lower than between human–chicken. 44 The study results of Dahmen et al. also showed few same STs of E. coli between pigs, cattle, and farm workers. 25 The above-mentioned results may explain why ST131 E. coli was not found in this study. Our results are consistent with the prior studies. Among our isolates, MLST and PFGE patterns showed a good correlation. In addition, 71% (5/7) ST4205 and 43% (3/7) ST10 isolates carried antibiotic resistance gene cassettes 7 and 1a, respectively. These results suggested the probability of clonal spread.
Conclusion
In conclusion, this study characterized the genetic basis of antimicrobial resistance in E. coli isolated from beef carcasses in Taiwan. To the best of our knowledge, this is the first report of isolation and molecular characterization of multidrug-resistant isolates of E. coli from beef meat in Taiwan. The inclusion of some resistance genes inside integrons constitutes an effective means to spread antibiotic resistance among bacteria from food products to humans. Therefore, it is important to monitor the risk of expanding antibiotic resistance through the food chain.
Footnotes
Acknowledgments
This work was supported, in part, by grants from the China Medical University (CMU105-S-24). We thank Prof. Kee-Ching Jeng, head of medical research of Tungs' Taichung MetroHarbor Hospital, for editing this article.
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.