
Abstract
Currently, echinocandins are first-line drugs for treatment of invasive candidiasis. However, data on how serum influences killing activity of echinocandins against uncommon Candida species are limited. Therefore, the killing activity of micafungin in RPMI-1640 and in 50% serum was compared against Candida guilliermondii, Candida lusitaniae, and Candida kefyr. Minimum inhibitory concentration (MIC) ranges in RPMI-1640 were 0.5–1, 0.12–0.25, and 0.06–0.12 mg/L, respectively. In 50% serum, MICs increased 32- to 256-fold. In RPMI-1640 ≥ 0.25, ≥4, and 32 mg/L micafungin was fungicidal against all four C. kefyr (≤4.04 hours), two of three C. lusitaniae (≤16.10 hours), and two of three C. guilliermondii (≤12.30 hours), respectively. In 50% serum, all three species grew at ≤4 mg/L. Micafungin at 16–32 mg/L was fungicidal against all C. kefyr isolates (≤3.03 hours) and at 32 mg/L was fungistatic against one of three C. lusitaniae isolates. Two C. lusitaniae isolates and all three C. guilliermondii grew at all tested concentrations. Adding human serum to susceptibility test media drew attention to loss of fungicidal or fungistatic activity of micafungin in the presence of serum proteins, which is not predicted by MICs in case of C. kefyr and C. lusitaniae in RPMI-1640. Our results strongly suggest that micafungin and probably other echinocandins should be used with caution against rare Candida species.
Introduction
A
Currently, echinocandins such as micafungin are first-line drugs for treatment of invasive candidiasis. 7 Echinocandins are highly protein bound with 0.2–3% free drug, that is, the serum and tissue concentrations may be low. 8 Decreased killing activity of echinocandins in 50% serum against medically important Candida species was confirmed.9–12 However, data on how serum influences killing activity of echinocandins against uncommon Candida species are limited.9–11,13 To extend our knowledge about the in vitro activity of echinocandins against uncommon Candida species, the killing activity of micafungin in RPMI-1640 and in RPMI-1640 plus 50% serum (50% serum) was compared with C. guilliermondii, C. lusitaniae, and C. kefyr.
Materials and Methods
Bloodstream-derived clinical isolates (Table 1) were identified using MICRONAUT-Candida and API ID32C methods. 14 All strains were isolated before antifungal administration. Micafungin pure powder was kindly provided by Astellas. Minimum inhibitory concentrations (MICs) in RPMI-1640 and in 50% serum (serum from a human male, type AB; Sigma, Budapest, Hungary) were determined using the standard broth macrodilution method. Final concentration ranged between 0.03 and 32 mg/L. MIC values were read visually after 24 hours using the partial inhibition criterion. 15
MIC, minimum inhibitory concentration.
Time kill studies were performed in both media at 1, 4, 16, and 32 mg/L at least twice. In case of C. kefyr 0.25 mg/L micafungin was also tested. The starting inocula were 3.5–5 × 105 CFU/ml. Aliquots of 100 μl were removed at 0, 4, 8, 12, 24, and 48 hours, serially diluted 10-fold, plated onto Sabouraud agar and incubated at 35°C for 48 hours. The same method was used to test growth kinetics.9–12,16,17
Fungicidal activity of micafungin was defined as 99.9% or higher reduction in viable cell count.9–13,16,17 Killing kinetics were analyzed in both media (RPMI-1640 with and without serum) as described previously.11,12 Briefly, an exponential equation was fitted to the mean data at each time point: Nt = N0 × e−kt, where Nt is the number of viable yeasts at time t, N0 is the number of viable yeasts in the initial inoculum, k is the killing rate, and t is the incubation time; reducing time points of each killing curve to one value (k). Positive k values indicate killing, negative values indicate growth. The goodness of fit was assessed by the r2 value (r2 > 0.8). The mean times to achieve the fungicidal endpoint (T99.9 = 3/k) were calculated from the k values for each isolate and concentration in both media.11,12
Killing kinetics among isolates and concentrations in either RPMI-1640 or 50% serum were compared using one-way ANOVA with Tukey's post testing; growth rates and the efficacy of the same micafungin concentrations in different media were compared by T-test (with Welch's correction, where appropriate).11,12
Results
C. guilliermondii isolates were susceptible to micafungin in RPMI-1640 (Table 1). 18 No breakpoint is available for C. kefyr and C. lusitaniae. However, MICs of isolates of these two species were not higher than the epidemiological cut off values (0.12 and 0.5 mg/L for C. kefyr and C. lusitaniae, respectively). 19 In 50% serum, MICs increased 32- to 256-fold. All C. kefyr isolates and C. guilliermondii isolate 5540 grew significantly better in RPMI-1640 than RPMI-1640 plus serum (p < 0.05).
Representative time-kill plots are shown in Fig. 1. In RPMI-1640, killing exerted by micafungin was rapid against C. kefyr; mean times to achieve 99.9% growth reduction were shorter than 4.04 hours for all isolates (Table 2). Contrastingly, 0.25, 1, and 4 mg/L micafungin did not produce any killing against C. kefyr isolates in serum (k values were negative and did not differ significantly from those of the drug-free controls) (Fig. 2 and Table 3). Higher concentrations (16 and 32 mg/L), however, produced fast (≤3.03 hours) killing in 50% serum as well.
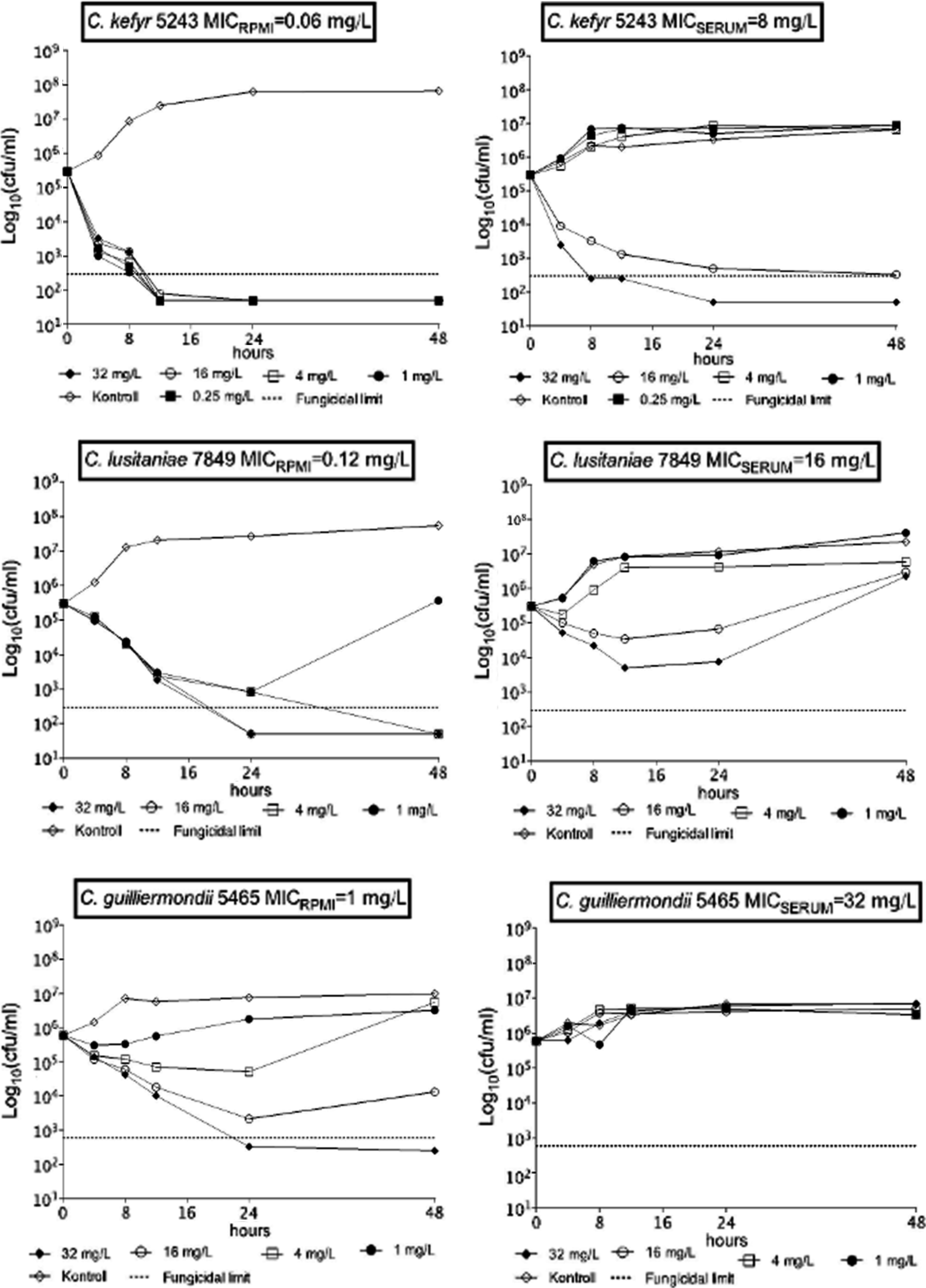
Representative time–kill curves against Candida kefyr, Candida lusitaniae and Candida guilliermondii in RPMI-1640 with (MICSERUM) and without 50% serum (MICRPMI).

Killing rates of micafungin and the corresponding adjusted regression lines (dashed lines) against four C. kefyr bloodstream isolates in RPMI-1640 with (serum) and without 50% serum (RPMI). Positive and negative k values indicate the decrease and increase, respectively, in viable cell numbers.
—, not determined; NA, not achieved.
Regrowth occurred.
CFU, colony-forming unit.
Micafungin at 4, 16, and 32 mg/L was fungicidal against two of three C. lusitaniae isolates in RPMI-1640 (Tables 2 and 3); the killing activity of micafungin was strain dependent, but concentration independent (i.e., k values did not differ significantly) (Fig. 3). In 50% serum, micafungin produced only transient colony-forming unit (CFU) decreases even at 16–32 mg/L against C. lusitaniae isolates (except isolate 3834 at 32 mg/L) (Table 3). Positive k value (0.2369 1/h, indicating killing) was noticed only at 32 mg/L in case of isolate 3834 (Fig. 3).

Killing rates of micafungin and the corresponding adjusted regression lines (dashed lines) against three–three C. lusitaniae (left) and C. guilliermondii (right) bloodstream isolates in RPMI-1640 (RPMI) and RPMI-1640 plus 50% serum (serum). Positive and negative k values indicate the decrease and increase, respectively, in viable cell numbers.
Although micafungin was fungicidal at 32 mg/L against C. guilliermondii isolates 5465 and 21060 in RPMI-1640, the inhibitory effect was only transient (regrowth occurred) at lower concentrations (1, 4, and 16 mg/L) (Table 3). In RPMI-1640, k values at 16 and 32 mg/L were strain dependent, but concentration independent (Fig. 3). In 50% serum, killing curves for C. guilliermondii at any concentrations were similar to controls (k values were always negative, indicating growth) (Figs. 1 and 3).
Discussion
It was previously shown that C. guilliermondii is intrinsically less susceptible to echinocandins, whereas wild-type C. kefyr and C. lusitaniae MIC values are close to those obtained with innately echinocandin-susceptible C. albicans or C. glabrata.6,19 However, mortality rates caused by C. guilliermondii, C. lusitaniae, and C. kefyr among seriously ill patients are extremely high (53–83%), suggesting that currently used therapeutic approaches, including echinocandin therapy, are insufficient.1–4 Furthermore, infections caused by C. lusitaniae and C. kefyr have become increasingly prevalent in parallel with the continuous increase in echinocandin use.1,6,20
The CLSI introduced clinical break-points for the five most important Candida species (C. albicans, C. glabrata, C. parapsilosis, C. tropicalis, and C. krusei), but not for uncommon (rare) Candida species (with the exception of C. guilliermondii).15,18 However, it is not taken into account that RPMI-1640 is a serum-free medium, thus in case of the highly protein-bound echinocandins, their in vitro potency against these Candida species may be overestimated in spite of the very low MIC values in standard susceptibility tests.6,18 Epidemiological cutoff values for the species against which official breakpoints has not yet been published are also based on the standard susceptibility testing using RPMI-1640.18,19
Serum-based testing of echinocandin susceptibility, which mimics the in vivo conditions occurring in the blood or tissues more closely, has some advantages. 21 First, in vivo efficacy of micafungin showed better correlation with MICs by serum-based than by standard RPMI-1640 susceptibility tests. 22 Second, 50% serum may help to differentiate between wild-type isolates and isolates harboring resistance mutations.6,21 Third, serum eliminates the paradoxically decreased killing activity at high concentration of echinocandins in RPMI-1640.12,18,21,23
Killing activities in this study were found to be markedly lower than against C. albicans 9 in the presence of serum, that is, no killing by 4 mg/L micafungin; even higher concentrations proved to be consistently fungicidal only against C. kefyr. Consequently, reported median peak and trough plasma levels (15.15 and 4.85 mg/L, respectively) produced by the elevated 150 mg micafungin daily doses, 24 predict inadequate eradication and increased risk of breakthrough infections 1 by these three species. Concordantly, only weak eradication capacity of echinocandins was reported in neutropenic animal models against these species even after 7–10 days echinocandin therapy (i.e., internal organs were not sterilized by echinocandins).25–27 In addition, inadequate serum echinocandin levels may facilitate development of acquired resistance to echinocandins. 6
In summary, adding human serum to susceptibility test media drew attention to loss of killing activity in the presence of serum proteins, which is not predicted by standard MICs in case of C. kefyr and C. lusitaniae in RPMI-1640. Even elevated therapeutic micafungin doses (150 mg/day) do not produce serum drug levels sufficient for reliable fungicidal or fungistatic activity against any of the three studied species, which may explain the observed high probability of therapeutic failure in infections with these species. This issue is especially important in case of patients with neutropenia or other immunosuppression. Although serum-based susceptibility testing has not yet been standardized, our results strongly suggest that micafungin and probably other echinocandins should be used with caution against rare Candida species.
Footnotes
Disclosure Statement
No competing financial interests exist.