
Abstract
Proteus mirabilis is the second most common cause of urinary tract infections and is also an important cause of nosocomial infections. TEM-type and CTX-M-type extended-spectrum β-lactamases (ESBLs) are the most widely distributed in this bacterial species, but minor ESBLs such as the VEB-type have also been identified. The aim of this study was to analyze the genetic environment of the blaVEB-4 gene found in a P. mirabilis clinical isolate recovered in Spain. P. mirabilis N2231 showed resistance to penicillins, cephalosporins, and aminoglycosides, remaining susceptible to imipenem, cefoxitin, β-lactamases inhibitors, and quinolones. Southern blot analysis revealed that blaVEB-4 was located in the chromosome. Analysis of the blaVEB-4 genetic context revealed a 15 kb segment 98% identical to the multidrug resistance (MDR) region of a Salmonella genomic island 1 (SGI1), which included a class 1 integron belonging to the In104 family, previously described in blaVEB-6-producing P. mirabilis VB1248. blaVEB-4 was surrounded by repeat elements, transposon Tn1721, and located on a class 1 integron containing aacA4-aadB-dfrA1-orfC genes. The blaVEB-4 gene was inserted in a complex structure of a class 1 integron, which is part of an MDR region of an SGI1, possibly involved in the mobilization of the gene and homologous recombination.
Introduction
P
Resistance to extended-spectrum cephalosporins in P. mirabilis is mainly due to the production of extended-spectrum β-lactamases (ESBLs) that belong to Ambler class A. TEM-type and CTX-M-type ESBLs are the most widely distributed among this species, but minor ESBLs, such as the VEB type, have also been identified. 2
VEB-type β-lactamases, which represent one of the smaller subgroups of class A β-lactamases,3,4 classified as group 2be enzymes 5 are a rare type of enzyme responsible for conferring high-level resistance to ceftazidime, cefotaxime, and aztreonam, although not carbapenems. There are 16 variants of VEB enzymes reported to date, derived from VEB-1 by one or more amino acid substitutions (www.lahey.org/studies/other.asp#table1). The blaVEB genes have been isolated from various species of Enterobacteriaceae and nonfermenter gram-negative bacilli (Pseudomonas aeruginosa, Acinetobacter baumannii, Achromobacter xylosoxidans)4,6–8 from Southeast Asia, Europe, the Middle East, Africa, and North and South America. 9 In P. mirabilis three variants have been reported: VEB-1, 2 VEB-4, 10 and VEB-6. 11 They are usually inserted within the variable regions of class 1 integrons and horizontally transmitted at the intra- and interspecific levels. 12 Genetic analysis of blaVEB-1 has revealed both chromosome and plasmid locations. 13 Interestingly, blaVEB-1 has also been reported on a class 1 integron located in an 86 kb resistance island. 14 The blaVEB-1 gene was first reported in an Escherichia coli isolate from a Vietnamese patient, where it was plasmid- and integron-located, 15 and subsequently in two P. aeruginosa isolates from Thailand, where it was chromosomally and integron-located. 11
We report in this study the genetic environment of the blaVEB-4 gene in a P. mirabilis clinical isolate N2231 recovered from a patient treated in a Spanish hospital.
Materials and Methods
Bacterial strain and antibiotic susceptibility testing
The P. mirabilis N2231 strain resistant to extended-spectrum β-lactams was isolated during a survey from February 2000 to December 2005 at Hospital de la Santa Creu i Sant Pau in Barcelona. Among 1423 P. mirabilis clinical isolates studied to determine the prevalence of ESBLs, we had found one VEB-4-producing isolate, the N2231, recovered from a urine culture. Reported in this study is the first description of this new variant (Accession No. EF136375). 10
Antimicrobial susceptibility testing was performed by the broth microdilution method, using GNX2F-layout plates (Sensititre; Thermo Fisher Scientific, Inc.) and the results were interpreted by Clinical and Laboratory Standards Institute guidelines (CLSI). 16
Conjugation experiments
Transferability of the resistance phenotype was studied by conjugation assays using a broth mating method at 37°C without shaking. The P. mirabilis N2231 VEB-4-producing strain was used as a donor and the modified E. coli Hb101 (UA6190) strain (rifampicin- and aminoglycoside-resistant, lactose-negative, and green fluorescent protein-producing) as a recipient strain. Transconjugants were selected based on fluorescence production and grown on LB agar plates supplemented with 10 mg/L ceftazidime and 100 mg/L rifampicin.
Genetic localization of the blaVEB-4 gene
The genetic location of blaVEB-4 was analyzed by pulsed-field gel electrophoresis (PFGE) and Southern blot. First, the genomic DNA was digested with the S1 nuclease to define plasmid localization. Second, the genomic DNA was digested with the I-CeuI restriction enzyme (New England Biolabs) to observe a chromosomal localization. After Southern transfer to a Hybond-N+ membrane (GE Healthcare) the fragments were hybridized with PCR-generated probes derived from purified DNA products obtained from the PCR of the blaVEB 10 and 16S rDNA genes (primers: 27F: 5′-AGAGTTTGATCMTGGCTCAG-3′; 907R: 5′-CCGTCAATTCMTTTRAGTTT-3′) marked with the PCR DIG probe synthesis kit (Roche, Spain). Detection was performed with antidigoxigenin antibody conjugated to alkaline phosphatase and CDP-Star chemiluminescence substrate (Roche) according to the manufacturer's instructions.
Genetic context of the blaVEB-4 gene
The genetic context of the blaVEB-4 gene in P. mirabilis N2231 was analyzed by inverse PCR and sequencing as previously described. 17 In brief, DNA was extracted by the GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich, Spain) from an overnight culture in LB (Luria–Bertani) broth at 37°C. The blaVEB gene was detected by PCR with primers described by Aragón et al. 10 The genomic DNA from strain N2231 was digested with the HindIII (Promega), and the fragments obtained were autoligated using T4 DNA ligase (Promega), following the manufacturers' instructions. The fragment of DNA containing the blaVEB-4 gene was used as a template for an inverse PCR with primers designed in this study from the blaVEB-4 gene sequence (VEB.inv-FW: 5′-GCAGAGTCCAAAGAAACTTCG-3′ and VEB.inv-R: 5′-GTCAGCTTGAGCATTTGAATAC-3′). Further PCR mapping was performed following the genetic environment of P. mirabilis VB1248 (HQ888851). Nucleotide and deduced amino acid sequences were analyzed and compared by means of the BLAST programs from the National Center for Biotechnology Information website (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
The schematic representation of the sequences, including their comparison and visualization, was generated by Easyfig. 18
Nucleotide sequence accession number
The genetic environment of the P. mirabilis N2231 strain has been deposited in the GenBank database under accession number KX859086.
Results
Susceptibility testing and transfer experiments
P. mirabilis N2231 showed resistance to ampicillin, ceftazidime, and cefotaxime (MICs >16, >32, and >16 mg/L, respectively) as well as some aminoglycosides (gentamicin, tobramycin, and amikacin [MICs >8, >32, and >32 mg/L, respectively]), tetracycline, ciprofloxacin, and trimethoprim/sulfamethoxazole (MICs >8, >2, and >4/76 mg/L, respectively), remaining susceptible to carbapenems (imipenem and ertapenem), cefoxitin and β-lactamase-inhibitors (amoxicillin–clavulanate and piperacillin–tazobactam). No transconjugants were obtained in transfer experiments with this strain.
Genetic location of the blaVEB-4 gene
Genomic DNA digestion with S1 nuclease and subsequent PFGE analysis and hybridization with the blaVEB probe were negative, pointing to a chromosomal localization for this gene. The blaVEB-4 chromosomal location was screened by Southern hybridization using the I-CeuI-PFGE profile with blaVEB and 16S rDNA probes. blaVEB and 16S rDNA PCR amplicons hybridized in the same ca. 291 kb I-CeuI fragment, suggesting that blaVEB-4 was located on the chromosome of P. mirabilis N2231 (Fig. 1).
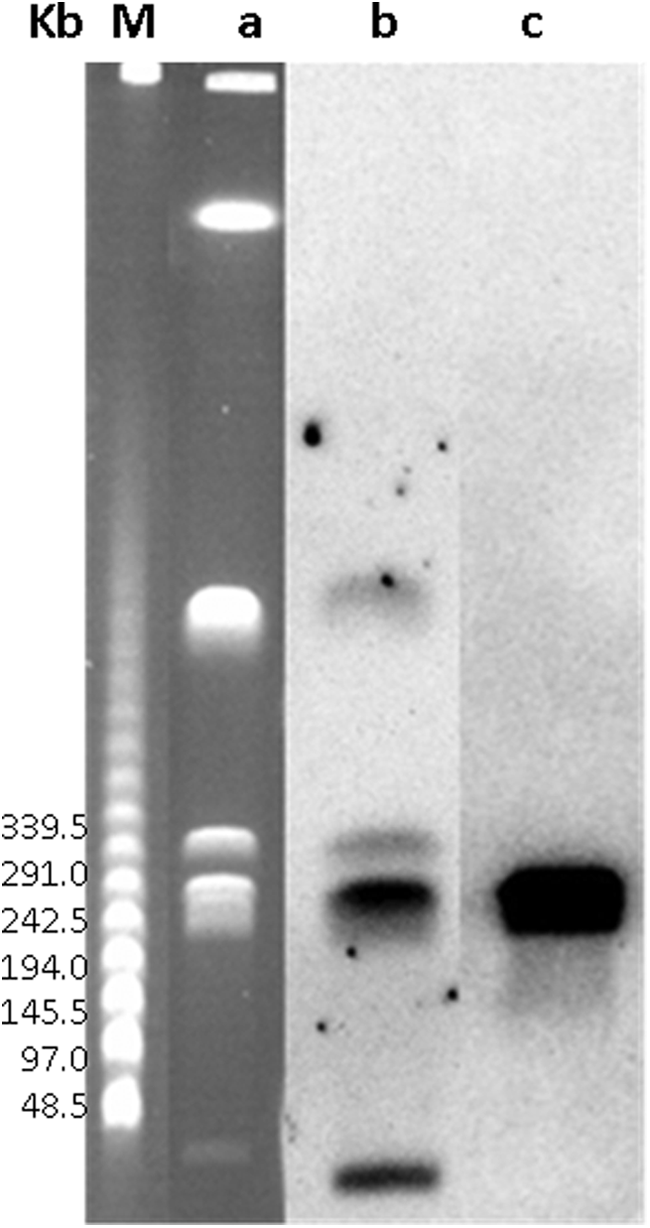
Southern hybridization of I-CeuI-PFGE. a, PFGE-I-CeuI; b, hybridization with 16S rDNA probe; c, hybridization with blaVEB probe, M, lambda ladder PFE marker. PFGE, pulsed-field gel electrophoresis.
Genetic context of the blaVEB-4 gene
Sequence analysis derived from the inverse PCR and PCR mapping in P. mirabilis N2231 revealed that the obtained 15 kb segment corresponded to the multidrug resistance (MDR) region of a Salmonella genomic island 1 (SGI1), and included a previously described complex class 1 integron. 19 The blaVEB-4 described herein was located in a similar integron as part of a truncated gene cassette in opposite orientation (Fig. 2). Interestingly, this 15 kb segment was 98% identical to those described by Siebor and Neuwirth 19 in a blaVEB-6-producing P. mirabilis VB1248 (HQ888851) and 100% identical to the deposited sequence described by Zong et al., 2 who also found blaVEB-6 in the P. mirabilis JIE273 chromosome. The blaVEB-4 gene was bracketed by two 135-bp repeated elements named Re1 and Re2, found in opposite orientation in comparison with the P. mirabilis VB1248. A 28-bp segment found between Re1 and the blaVEB-4 was absent in the same region in the P. mirabilis VB1248. The structure found downstream of Re1 until the insertion sequence IS6100 (transposase) was identical to P. mirabilis VB1248 (Fig. 2).
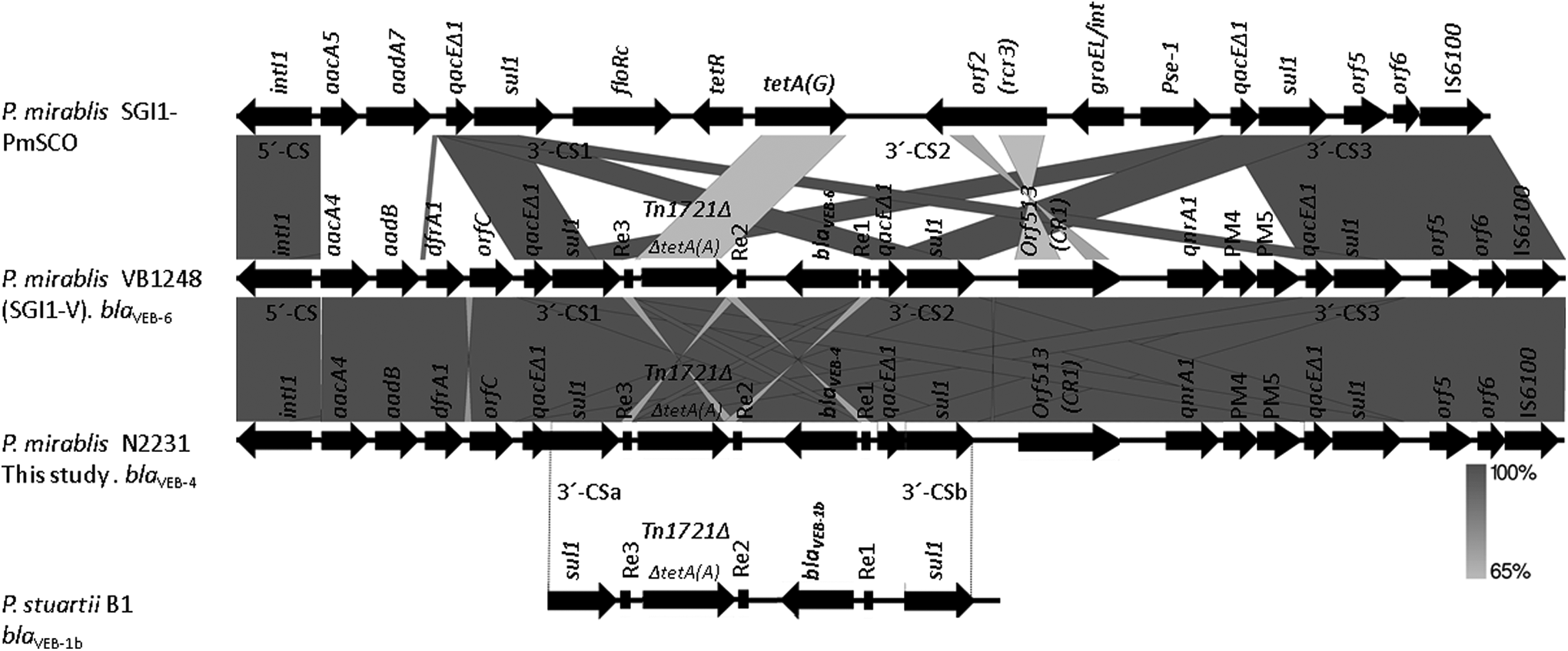
Schematic view of the genetic context of the blaVEB-4 gene in Proteus mirabilis N2231 and comparative structures with the MDR region within the SGI1 in P. mirabilis SGI1-PmSCO (GenBank accession number JX121639), P. mirabilis VB1248 (SGI1-V) (GenBank accession number HQ888851) and P. stuartii B1. Conserved segments (5′-CS and 3′-CS). The repeat elements (Re1, Re2, and Re3). Regions of homology are shaded in gray scale.19,28 MDR, multidrug resistance; SGI1, Salmonella genomic island 1.
In opposite orientation to blaVEB-4, a 432 bp segment corresponding to a hypothetical protein was found, followed by the Tn1721Δ-like truncated tetracycline resistance transposon, including the tet(A) gene (encoding a class A tetracycline efflux protein), 20 which was bracketed by the repeated elements Re2 and Re3. Upstream to Re3 was detected a third copy of a 3′-conserved segment (3′-CS1) and the conservative structure of a class 1 integron containing four gene cassettes: aacA4 and aadB genes (conferring resistance to aminoglycosides), dfrA1 (conferring resistance to trimethoprim), and orfC with unknown function. At the terminal left-hand end, a fragment of a 5′-conserved segment (5′-CS) containing intI1 (integrase) was found.
Discussion
The aim of this study was to characterize the genetic surroundings of the blaVEB-4 gene found in the P. mirabilis N2231 clinical isolate. 10 The well-known VEB β-lactamase has a wide dissemination and an increasing number of novel variants recovered from humans or food indicate a rapid evolution and spread.19,21 P. mirabilis isolates naturally lack bla genes on their chromosome and are therefore generally susceptible to all β-lactam antibiotic agents. However, a progressive increase of multiresistant strains has been occurring in this species, 22 which has demonstrated a great ability to acquire resistance genes, such as inhibitor-resistant TEM β-lactamases, ESBLs, and ampC β-lactamases. 23
Sequence analysis of the genetic context of the blaVEB-4 described herein revealed a 15 kb segment corresponding to an MDR region of a SGI1, including a previously described class 1 integron belonging to the In104 family. 19 SGI1-like variants have been detected among bacteria other than Salmonella, including P. mirabilis.24–26 This MDR region included frequently described resistance genes, such as aacA4, aadB, sul1, dfrA1, and tet(A) in addition to the blaVEB-4 and qnrA genes. Our results were very similar (98% identity) to those reported by Siebor and Neuwirth, 19 who found blaVEB-6 in the MDR region of a SGI1-V from a P. mirabilis clinical isolate, and 100% identical to the reported sequence described by Zong et al. 2 The latter found blaVEB-6 in the P. mirabilis JIE273 chromosome in a truncated gene cassette, similarly flanked by three repeated elements Re1, Re2, and Re3, including the Tn1721Δ-like transposon and duplications of the 3′-CSs of a class 1 integron.
Siebor and Neuwirth 26 have hypothesized that P. mirabilis is involved in the dissemination of SGI1, since they found that the same modifications of the SGI1 backbones occur in Salmonella. A schematic comparison of the MDR regions among P. mirabilis strains, N2231 from this study, SGI1-PmSCO, 26 and VB124819 (Fig. 2), might suggest the presence of an SGI1 element in our strain, considering the similarity of the structures, particularly with P. mirabilis VB1248 (98% identity). Interestingly, the blaVEB-6 variant is essentially identical to blaVEB-4, varying in only one nucleotide (A52G), which predicts only a conservative amino acid substitution (Ile18Val) in the leader peptide.
The blaVEB-1 gene is often part of a gene cassette located in class 1 integrons. 27 Worthy of mention, the immediate genetic environment (3′-CS-Re3-Tn1721Δ-Re2-blaVEB-Re1-3′-CS) of the blaVEB-4 in P. mirabilis N2231 included the 135-bp repeat elements (Re1, Re2, and Re3) bracketing the blaVEB-4 gene and duplications of the 3′-CS of class 1 integrons. Notably, this structure is similar to that observed for blaVEB-1a in P. aeruginosa, 6 blaVEB-1b in P. stuartii B1, 28 and blaVEB-6 in P. mirabilis JIE2732 and P. mirabilis VB124819 (Fig. 2), suggesting that blaVEB is associated with highly conserved genetic structures. In addition, the presence of blaVEB in different bacterial species illustrates how resistance genes may spread via conjugative plasmids and/or integrons, 11 as well as transposable elements. This is of concern in the context of dissemination of resistance genes via horizontal gene transfer.
The high similarity between these genetic contexts, particularly in three P. mirabilis strains with a chromosomally located blaVEB gene, JIE273, 2 VB1248, 19 and N2231 of our study, indicate a common ancestry, possibly mediated by SGI1 elements, taking into account that P. mirabilis is an important host of SGI1. 26 The association of blaVEB variants with MDR regions of SGI1, cassette arrays, repeat elements bracketing the blaVEB genes, duplications of the 3′-CS of a class 1 integron, and components of transposons further support this hypothesis. The presence of these components could also play a role in the movement of the blaVEB gene and suggest possible recombinations and rearrangements of a limited set of different elements, as described by Zong et al. 2
In addition, differences in genes at both ends of the blaVEB structures could be explained by these recombination events, which can mediate large evolutionary jumps in bacterial genomes by rapidly spreading variants associated with increased virulence, fitness, and antibiotic resistance. Interestingly, a recent study reported transmission of blaVEB-6 through the food chain to humans: Seiffert et al. 21 identified blaVEB-6 in a P. mirabilis isolate from poultry meat in Switzerland. The blaVEB-6 from that study had a nucleotide sequence identical to that found in the human VEB-6-positive P. mirabilis VB1248 reported in France.
This is the first report describing the genetic environment of the blaVEB-4 gene, detected in a P. mirabilis N2231 clinical isolate from Spain. The chromosomally located blaVEB-4 gene was inserted in a complex structure of a class 1 integron, which is part of an MDR region of an SGI1, possibly involved in the mobilization of the gene and homologous recombination.
This structure carried several genes whose products confer resistance to aminoglycosides, tetracycline, sulfonamides, and all tested β-lactams, except carbapenems. Therefore, multiresistant strains harboring several resistant genes and complex structures require special attention.
Footnotes
Acknowledgments
The project was supported by the Plan Nacional de I+D+i and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Economía y Competitividad, Spanish Network for Research in Infectious Diseases (REIPI RD12/0015/0017); cofinanced by European Development Regional Fund “A way to achieve Europe” ERDF. P.E. is funded by the Instituto de Salud Carlos III “Sara Borrell” contract CD15/00017.
Disclosure Statement
No competing financial interests exist.