
Abstract
Methicillin-resistant Staphylococcus aureus (MRSA) has an ability to invade nonprofessional phagocytic cells, resulting in persistent infections and most likely host cell death. Series of our studies have claimed pronounced antibacterial efficacy of Rhodomyrtus tomentosa leaf extract. This study was to further investigate potency of the extract in intracellular killing of human HaCaT keratinocytes. Pretreatment of MRSA with the extract resulted in a remarkable reduction in the bacterial adhesion to HaCaT keratinocytes, compared with untreated control (p < 0.001). In addition, at least 60% inhibition of the bacterial invasion into HaCaT cells was observed. Intracellular killing assay demonstrated that the extract exhibited strong antibacterial activity against intracellular MRSA at nontoxic concentrations (128 mg/L), which may have resulted from the increase in bactericidal activity under phagolysosomal pH. Transmission electron microscopy displayed the effects of the extract on alterations in the bacterial cell morphology with cell lysis. Fluorescence microscopy revealed that the extract decreased MRSA-induced apoptosis in HaCaT cells. In addition, cytotoxicity of HaCaT cells caused by MRSA supernatant was reduced at least 50% by the extract. The potential activities of R. tomentosa extract may be useful in an alternative treatment of MRSA infections in slight acidic compartments, particularly skin infections.
Introduction
S
Recently, natural compounds have been of interest to control staphylococcal infections by inhibiting the virulence factors with due attention to the increasing resistance to chemically synthesized antibiotics. Secondary metabolites produced by plants, for example, phenolic and flavonoid compounds, can be used as antimicrobial agents to control infectious diseases. 9 They are able to interfere with the bacterial pathogenesis by inhibition of quorum sensing, 10 biofilm formation, 11 colonization, 12 and inflammation. 13 In addition, a bioactive phenolic compound, phloroglucinol derivative, showed several biological properties, for example, antimicrobial, antiprotozoal, antioxidant, and antiviral activities.14,15 However, according to ethnomedical use in a primary healthcare system, traditional plant extracts have been focused on since they are commonly employed due to low toxicity and contain several bioactive compounds that may have synergistic effects. In addition, plants can be grown locally, resulting in the increase in accessibility to treatment for the inaccessible patients.
Rhodomyrtus tomentosa is an evergreen shrub plant distributed throughout tropical Asia and has been used as an antiseptic wash to clean wounds,16,17 as well as in skin beautifying agents. 18 The antiseptic solution from a decoction of the leaves has been applied to treat infectious wounds, impetigo, and abscesses. 17 Phytochemical characterization of R. tomentosa ethanol leaf extract showed that the major chemical constituents include primary metabolites such as oils, proteins, and carbohydrates, as well as secondary metabolites such as alkaloids, flavonoids, saponins, steroids, and terpenoids. 16 Recently, an acyphloroglucinol derivative, rhodomyrtone, isolated by column chromatography has been claimed as a new antimicrobial agent against gram-positive bacteria.19,20
We focused on R. tomentosa leaf extract based on our previous reports, which demonstrated its efficacy on the killing of gram-positive bacteria 20 and the bacteria growing in the biofilm. 21 Recently, it has been reported that the extract and rhodomyrtone could reduce adhesion of pathogens to plastic surfaces. 22 Moreover, both agents showed antistaphylococcal adherence to bovine epidermal cells. 23 Based on the traditional use for treatment purposes, we further investigated effects of R. tomentosa leaf extract against MRSA colonization of human HaCaT keratinocyte cells and their activity to kill the intracellular pathogens.
Materials and Methods
Preparation of R. tomentosa ethanol extract
R. tomentosa leaves were dried in an oven at 45°C for 7 days, then ground, and the powder was soaked with 95% ethanol at room temperature for 7 days. The extract was filtered through a Whatman filter paper No. 1 and concentrated using a rotary evaporator (BUCHI Rotavapor R-114). The concentrated extract was kept in an oven at 37°C for 10 days to completely dry and dissolved in dimethyl sulfoxide (DMSO; Merck) before use. The percentage yield of the extract was found to be 7.8%.
Bacterial strains and growth conditions
Seventeen clinical isolates of MRSA were kindly received from the Natural Product Research Center of Excellence (NPRCoE), Faculty of Science, Prince of Songkla University, Thailand. MRSA NPRC 001R, a multidrug-resistant strain, was used as a representative isolate for further studies. S. aureus ATCC 25923 and ATCC 29213 were included as controls. The bacteria were precultured at 37°C for 24 hr on trypticase soy agar (TSA). The single colony was transferred into 3 ml TSB and grown at 37°C with shaking overnight. The overnight cultures were diluted 1:100 with fresh TSB and continually shaken for 4, 18, and 24 hr to allow the cells to reach mid-exponential, stationary, and late stationary phases, respectively. After incubation, the cultures were centrifuged at 7,000 rpm for 10 min. The cell pellet was collected, washed thrice with phosphate-buffered saline (PBS), pH 7.4, and resuspended in Dulbecco's modified Eagle's medium (DMEM, low glucose; Life Technologies) supplemented with 10% fetal bovine serum (FBS; HyClone). The bacterial suspension was adjusted to ∼1 × 109 CFU/ml (OD600nm of 1) and confirmed by counting colonies on TSA. For some studies, the bacterial cells were killed at 55°C for 1 hr in a water bath.
Cell culture
Human HaCaT keratinocyte cells were grown in DMEM supplemented with 10% FBS and 50 mg/L gentamicin at 37°C in a humidified incubator with 5% CO2.The keratinocytes were passaged every 4–5 days at a ratio of 1:8 by incubating with EDTA in PBS (0.2 g/L; Merck) for 10 min and trypsin-EDTA (0.25%; Sigma) for 10 min, respectively. The medium was changed every 2 days.
Determination of the antibacterial activity of R. tomentosa extract against extracellular S. aureus
Minimal inhibitory concentration (MIC) and minimal bactericidal concentrations (MBC) of the crude extract, H2O2, and lysozyme against S. aureus ATCC 29213 and MRSA were performed by a modified broth microdilution method recommended by Clinical Laboratory Standardization Institute guideline. 24 Mueller-Hinton broth (MHB) was adjusted to pH 7.3 and pH 5.0 to mimic the pH conditions of extracellular medium and phagolysosomes, respectively. The extract, H2O2, and lysozyme were serially diluted 2-fold in MHB (both pH 7.3 and pH 5.0) to give final concentrations ranging from 4 to 1,024 mg/L, 2 to 2,048 mg/L, and 60 to 62,500 mg/L, respectively. An exponential phase of S. aureus was adjusted to an OD600nm of 0.01with MHB (both pH 7.3 and pH 5.0) to achieve a concentration of ∼5 × 106 CFU/ml. One hundred microliters of the suspension was added to each well of a 96-well plate containing 100 μl of the test agents and incubated at 37°C for 16–18 hr. Ten microliters of resazurin (Molecular Probes) was added to each well and incubated for 1 hr. The MIC was defined as the lowest concentration of antimicrobial agent giving the resazurin color equal to that of uninoculated broth. Aliquots of the broth were dropped onto MHA plates and incubated at 37°C overnight to determine MBC value. The antibacterial concentration that completely killed the bacterial growth was defined as the MBC value.
Influence of intracellular conditions on antibacterial activity
The MICs and MBCs of H2O2 and lysozyme in the presence of the extract at subinhibitory concentrations under phagolysosomal pH (pH 5.0) were performed as described above. This assay was carried out in a 96-well plate containing 50 μl of H2O2 or lysozyme, 50 μl of the extract (8 mg/L), and 100 μl of the bacterial suspension.
Bacterial adhesion and invasion assay
To investigate the effect of the extract on bacterial adhesion to human keratinocytes, MRSA NPRC 001R was treated with the extract at 4 to 16 mg/L for 18 hr. The treated bacteria (1 × 107 CFU/ml) were coincubated with HaCaT keratinocytes (1 × 106 cells/ml) in a 24-well plate containing Thermanox glass coverslips and incubated for 30 min in 5% CO2. The monolayers on coverslips were washed twice with PBS, detached with trypsin-EDTA (0.25%), and then lysed with 0.025% Trixon X-100. The lysed suspension was plated onto TSA and incubated overnight to determine the bacterial attachment to HaCaT cells.
For invasion assay, the bacteria were pretreated with the extract for 4, 18, and 24 hr, followed by incubation with HaCaT keratinocytes. After 60 min of internalization, the coverslips were washed twice with PBS and transferred to a new 24-well plate containing gentamicin (100 × MIC) to kill any remaining extracellular bacteria for 60 min. Subsequently, the monolayers were washed twice with PBS, treated with 0.25% trypsin for 10 min in 5% CO2, and lysed by addition of 0.025% Triton X-100. The cell lysates were serially diluted 10-fold and plated on TSA to quantify the internalized bacteria. The inhibition of adhesion and invasion was calculated according to the following formula:
Inhibition (%) = [1 − (CFU/ml of the treated sample)/(Mean CFU/ml of the untreated control)] × 100.
Determination of the antibacterial activity of R. tomentosa extract against intracellular S. aureus
Intracellular infection assays were modified from a published study. 25 Briefly, HaCaT keratinocytes (1 × 106 cells/ml) were infected with MRSA at different concentrations (1 × 105, 1 × 106, and 1 × 107 CFU/ml) in a 24-well plate and incubated for 60 min in 5% CO2. The infected keratinocytes were washed twice with prewarmed PBS. After bacterial removal, extracellular MRSA was killed by gentamicin at a final concentration of 500 mg/L for 1 hr. The infected keratinocytes were washed twice with prewarmed PBS, transferred to a new 24-well plate containing the extract at concentrations of 16 to 128 mg/L (16 mg/L, which is equivalent to 1 × MIC), and then incubated for 24 hr. The infected cells without the extract were used as a control. The infected cells were washed four times with PBS, detached with 0.25% trypsin at 37°C for 10 min, and resuspended in 0.025% Triton X-100 to lyse HaCaT cells. The viable bacteria in the cell lysates were plated on TSA and incubated overnight. The killing of MRSA by HaCaT keratinocytes in the presence or absence of the extract was investigated by examining the reduction in the number of viable intracellular bacteria.
Morphological studies
Transmission electron microscopy (TEM) analysis was performed to observe the morphological change in intracellular MRSA. Therefore, this assay was carried out as previously described. Briefly, HaCaT cells were grown in a six-well plate until confluent growth and then infected with MRSA (1 × 106 CFU/ml). After 60 min of incubation, the infected cells were treated with the extract at 64 mg/L as a treatment group and with vancomycin at 1 × MIC as a control group for 24 hr. The cells were washed thrice with PBS, fixed in 3.7% glutaraldehyde at room temperature for 60 min, and then washed twice with PBS. The sample was processed for TEM analysis by fixation with 1% osmium tetraoxide, followed by ethanol dehydration, and was embedded in Epon 812 resin. The morphology was imaged using a JEOL JEM-2010.
Assessment of apoptosis
HaCaT keratinocytes were plated in a six-well plate containing a glass coverslip until confluent growth. The monolayers (1 × 106 cells/ml) were washed and incubated with MRSA (1 × 106 CFU/ml) for 60 min. The infected cells were washed thrice with prewarmed DMEM and treated with the extract at 64 mg/L as a treatment group and with vancomycin at 1 × MIC as a control group for 24 hr. The cells were washed twice with prewarmed PBS and fixed with 3.7% formaldehyde at room temperature for 10 min. The fixed cells were washed with PBS, stained with 5 mg/L Hoechst 33342 (Life Technologies), and incubated for 15 min in the dark. The samples were washed thrice with PBS and the nuclear morphology of MRSA-infected HaCaT cells in the presence or absence of the extract was observed under a fluorescent microscope.
Human skin epithelial toxicity assay
The human skin epithelial toxicity assay was adapted from a published procedure. 26 In this study, the effect of R. tomentosa extract as a quorum-sensing inhibitor was investigated by determining the survival of HaCaT keratinocytes exposed to MRSA supernatant. MRSA was grown overnight in TSB, followed by dilution (1:500) with fresh TSB containing the extract at 4 to 16 mg/L and incubation for 18 hr with shaking to reach stationary phase. The bacterial suspension was centrifuged and then filtered through a syringe filter (0.22 μm; Corning). The sterile supernatant was diluted 1:2 in DMEM supplemented with 10% FBS. Heat-inactivated supernatant (56°C for 60 min) was included as a nontoxic supernatant. Two hundred microliters of the supernatant was added to each well of a 96-well plate containing HaCaT monolayers and incubated for 24 hr in 5% CO2. The supernatant was discarded and replaced by the addition of fresh DMEM supplemented with 10% FBS. To determine cytotoxicity, 10 μl resazurin (final concentration 50 mg/L) was added to HaCaT cells and incubated for 4 hr. The absorbance of viable cells was measured at 570 and 600 nm using a microplate reader (Biotex PowerWaveX-I).
Cell viability assay
HaCaT cells were seeded into a 96-well plate at a density of 2 × 104 cells/ml and incubated at 37°C for 24 hr in a CO2 incubator. The monolayers were incubated with the extract at concentrations ranging from 4 to 512 mg/L for 6 and 48 hr. DMSO at 1% was used as a negative control. At specific time points, HaCaT cells were incubated with 10 μl resazurin (final concentration 50 mg/L) for 4 hr and the absorbance was read at 570 and 600 nm. The percentage of viable cells or reduction of resazurin to resorufin by viable cells was calculated according to the following formula:
Where, O1 = molar extinction coefficient (E) of oxidized resazurin at 570 nm (O1 = 80,586), O2 = E of oxidized resazurin at 600 nm (O2 = 117,216), A1 = absorbance of test wells at 570 nm (HaCaT cells incubated with the extract), A2 = absorbance of test wells at 600 nm (HaCaT cells incubated with the extract), P1 = absorbance of positive growth control well (HaCaT cells plus resazurin, but without the extract) at 570 nm, and P2 = absorbance of positive growth control well (HaCaT cells plus resazurin, but without the extract) at 600 nm.
Statistical analysis
Data are presented as the mean ± SEM of the results of at least three independent experiments performed in triplicate samples. Paired two-tailed Student's t-test was used to compare the test and control groups. p < 0.05 were taken to indicate statistical significance.
Results
R. tomentosa extract inhibits MRSA adhesion and invasion into human keratinocyte cells
Colonization and invasion are important phenomena of S. aureus that contribute to the pathogenesis of bacterial infections. Therefore, we evaluated the effects of R. tomentosa extract on MRSA adhesion to human HaCaT keratinocytes as well as their invasion. The bacterial cells were pretreated with the extract for 18 hr, followed by exposure to HaCaT cells, and the results are given in Figure 1. The extract at 4, 8, and 16 mg/L significantly inhibited the bacterial adhesion to keratinocyte cells to 37.5%, 62.5%, and 76.5%, respectively, compared with the control (p < 0.001), indicating that the inhibitory effect of the extract was dose dependent.

Inhibitory effects of Rhodomyrtus tomentosa extract on attachment of methicillin-resistant Staphylococcus aureus (MRSA) to HaCaT keratinocytes. MRSA was treated with the extract at 4, 8, and 16 mg/L (MIC is equivalent to 16 mg/L) for 18 hr. HaCaT cells (1 × 106 cells/ml) were incubated with the extract-treated MRSA (1 × 107 CFU/ml) for 30 min. The bacterial cells without treatment were used as control. Adhering bacteria were counted on agar plates. Data are the mean ± SEM from two independent experiments with four technical replicates. ***p < 0.001, compared with the control. MIC, minimum inhibitory concentration.
The effects of the extract on MRSA invasion in relation to different growth phases were investigated. In this study, the bacterial cells were treated with the extract for 4, 18, and 24 hr according to the bacterial growth phases, including exponential, stationary, and late stationary phases, respectively. No effects on the bacterial invasion were observed after 4 hr of exposure (data not shown). In contrast, pretreatment of MRSA with the extract at sub-MIC and MIC for 18 hr significantly decreased the bacterial invasion into HaCaT keratinocytes when compared with the untreated bacteria (p < 0.01, Fig. 2). Moreover, MRSA treated with the extract for 24 hr was able to reduce the bacterial internalization to a significantly greater extent than the controls (p < 0.0001). The results demonstrated that the inhibition of bacterial invasion is dependent on their growth phases.
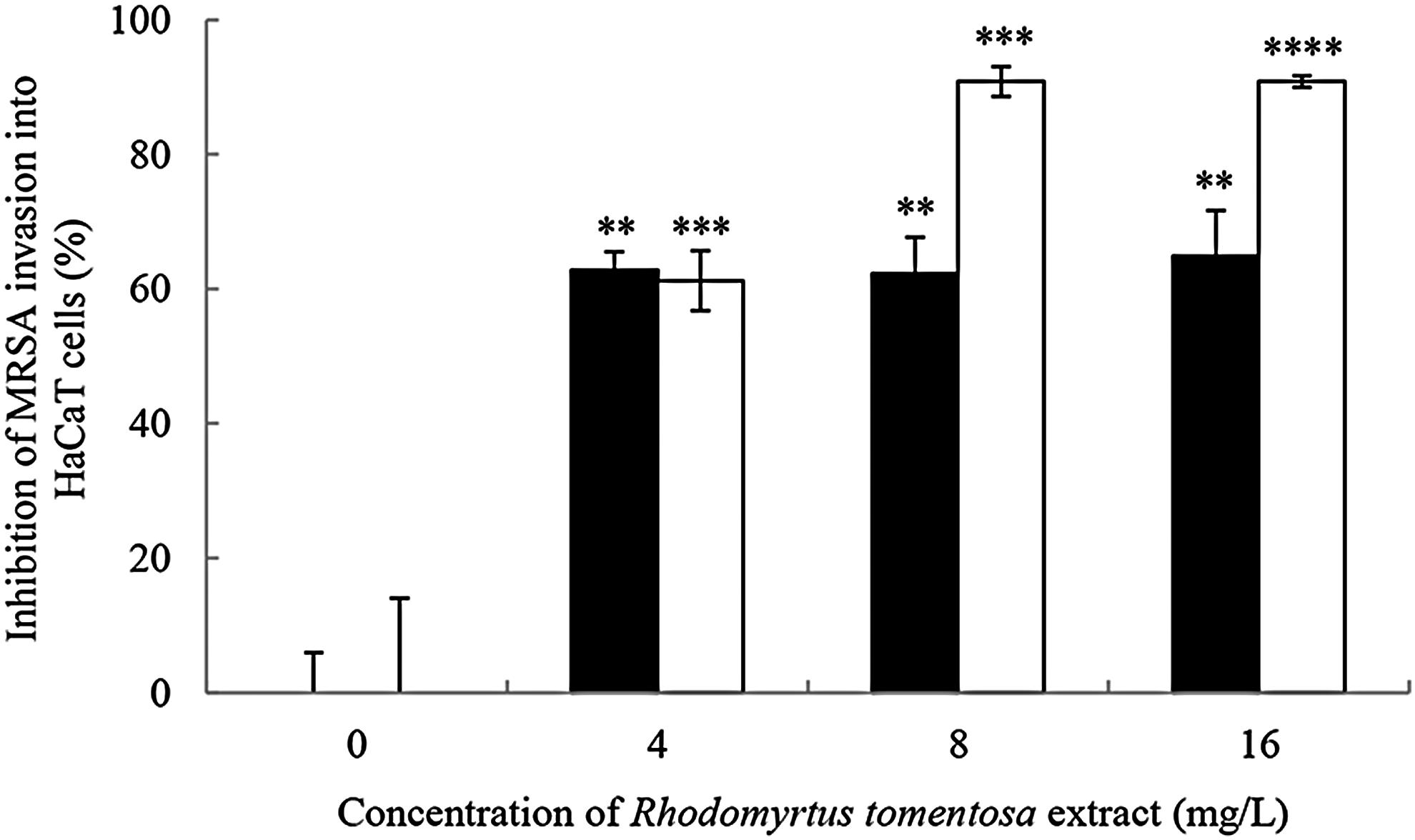
Effects of R. tomentosa extract on invasion of MRSA into HaCaT keratinocyte cells. The bacterial cells were treated with the extract at 4, 8, and 16 mg/L (MIC is equivalent to 16 mg/L) and then incubated to reach stationary phase (18 hr, ■) and late stationary phase (24 hr, □). HaCaT cells (1 × 106 cells/ml) were incubated with the extract-treated MRSA (1 × 107 CFU/ml) for 60 min. A control was treated with the medium alone. The inhibitory effects of invasion are determined by counting the number of invading bacteria. Data are the mean ± SEM from two experiments with four technical repeats. **p < 0.01, ***p < 0.001, and ****p < 0.0001, compared with the control.
Intracellular killing activity of R. tomentosa extract against MRSA infection of HaCaT keratinocytes
Bacterial entry into their host cells may result in persistent infections, which is dependent on bacterial density at the site of infection. We investigated whether R. tomentosa extract could kill intracellular MRSA within HaCaT keratinocytes. The cells (1 × 106 cells/ml) were infected with MRSA at different densities (105–107 CFU/ml) and further incubated with increasing concentrations of the extract. As presented in Figure 3, the extract at 16–128 mg/L could lower the intracellular pathogens up to 61–98%, respectively, when the bacterial load at 105 CFU/ml was used (p < 0.01). When infected with 106 CFU/ml, the decrease in the numbers of internalized bacteria was detected significantly at 40–83% by treatment with the extract at the same concentrations (p < 0.001). However, no considerable inhibitory activity of the extract against the intracellular pathogens was observed at the high bacterial density (107 CFU/ml) (data not shown). It can be concluded that R. tomentosa extract is able to kill MRSA within HaCaT cells, which is dependent on both the extract concentrations and infectious doses.
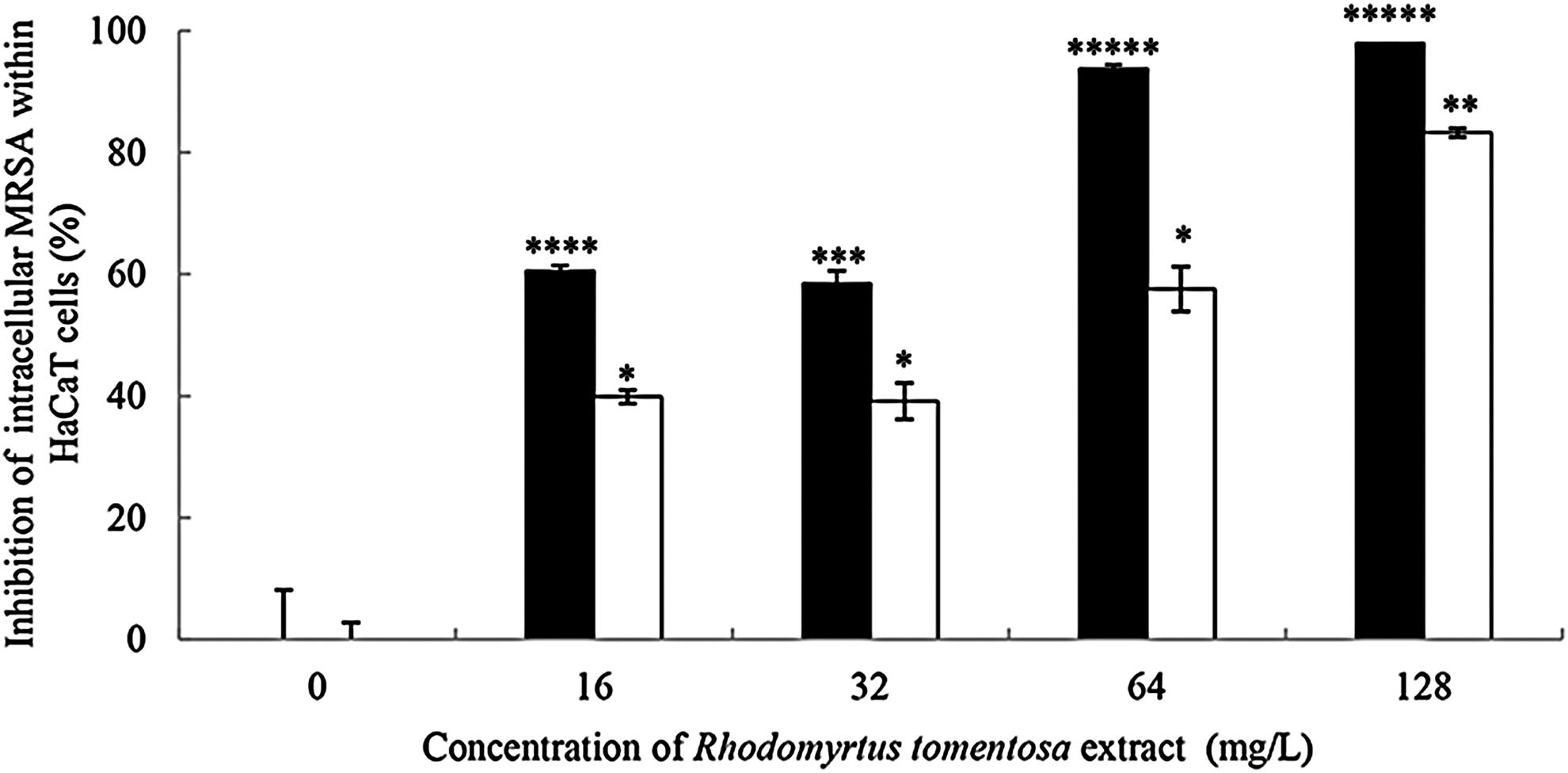
Antibacterial activities of R. tomentosa extract against intracellular MRSA. HaCaT keratinocytes (1 × 106 cells/ml) were seeded into a 24-well plate for 24 hr, followed by MRSA infection at concentrations of 1 × 105 CFU/ml (■) and 1 × 106 CFU/ml (□) for 1 hr. The infected cells were washed and then treated with the extract at concentrations from 16 to 128 mg/L (MIC is equivalent to 16 mg/L) for 24 hr. Control cells were treated with vancomycin at 1 × MIC to prevent the extracellular bacterial growth. Data are the mean ± SEM from two experiments with four technical replications. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, and *****p < 0.00001, compared with the control.
TEM analysis was further carried out to demonstrate morphological changes of intracellular MRSA within the keratinocytes (Fig. 4). Normal cell division and intact cell wall of the pathogens were observed in the controls (Fig. 4A). Interestingly, bacterial morphologies of intracellular MRSA treated with the extract at 64 mg/L for 24 hr exhibited alterations in cell shape, atypical sizes such as oval shaped (Fig. 4B) as well as swollen cells (Fig. 4C, F), and completely lysed cells with the broken walls and the release of their cytoplasmic materials (Fig. 4D). In addition, they displayed impaired cell separation with irregular septa (Fig. 4E–G).

Transmission electron microscopic micrographs of intracellular MRSA after exposure to R. tomentosa extract. HaCaT keratinocytes (1 × 106 cells/ml) were infected with MRSA (1 × 106 CFU/ml) for 1 hr and then media were removed to discard the extracellular bacteria. The intracellular bacterial cells were incubated in the absence
As intracellular MRSA can cause damages to the host cells, we further examined whether the extract could decrease apoptosis of HaCaT keratinocytes. As shown in Figure 5B, HaCaT cells infected with the pathogens demonstrated an obvious increase in the numbers of apoptotic cells, which is characterized by chromatin condensation and fragmentation when compared with the control cells (Fig. 5A). In contrast, the apoptotic HaCaT cells caused by MRSA infections were considerably reduced by treatment with the extract (Fig. 5C).

Effects of R. tomentosa extract on nuclear morphology of MRSA-infected HaCaT keratinocytes. HaCaT cells were seeded for 24 hr in a 6-well plate at a density of 1 × 106 cells/ml
Protective effects of R. tomentosa extract against MRSA-secreted protein-induced damage to HaCaT keratinocytes
Based on previous work that quorum sensing has been strongly inhibited by R. tomentosa extract, 21 damages to HaCaT cells by MRSA-secreted proteins controlled by quorum-sensing system was investigated. The bacterial cells were grown in the presence of the extract at 4, 8, and 16 mg/L and the supernatant was exposed to HaCaT keratinocytes. Up to 94.47% toxicity was observed in HaCaT cells treated with MRSA supernatant. In contrast, the cytotoxicity of the keratinocytes was significantly reduced by at least 50% by the extract at 8 and 16 mg/L (p < 0.001). Similar results were also found in the heat-inactivated supernatant as a negative control (Fig. 6).

Effects of R. tomentosa extract on cytotoxicity of HaCaT keratinocytes caused by MRSA supernatant. Overnight culture of MRSA was grown in the presence of the extract at concentrations from 4 to 16 mg/L for 18 hr. The bacterial supernatant was harvested and filtered (0.22 μm). Sterile supernatant was exposed to confluent HaCaT cells (1 × 106 cells/ml) for 24 hr, followed by adding resazurin to determine the cytotoxicity. HaCaT cells with the medium alone were used as an untreated control. Heat-inactivated MRSA supernatant was used as a negative control. Data are the mean ± SEM from three biological experiments. ***p < 0.01, ****p < 0.001, compared with no extract control.
Antibacterial activity of R. tomentosa extract at acidic pH condition mimicking phagolysosomal environment
S. aureus has the ability to resist phagolysosome killing by modifying their mechanisms to prevent lysozyme and respiratory burst activities. Therefore, in the following work, susceptibility of S. aureus to the extract in the presence of acidic pH, H2O2, or lysozyme was determined (Table 1). The MICs of the extract against both S. aureus ATCC 29213 and MRSA at pH 5.0 modestly diminished by 1-fold dilution, compared with those at neutral pH. In addition, the decrease in MBCs of the extract for both strains at pH 5.0 by 4–5-fold dilutions was clearly observed, compared with the values at pH 7.3. In contrast, there were no changes in MIC and MBC values of the extract in the presence of H2O2 and lysozyme. To confirm that the antibacterial activities of R. tomentosa extract were increased by the acidic pH, 16 clinical strains of MRSA and reference strains of S. aureus ATCC 29213 were additionally studied. As presented in Figure 7, the MIC and MBC values at neutral pH of the extract against the tested bacteria ranged from 16 to 128 mg/L and 512 to 1,024 mg/L, respectively. In contrast, the acidic pH caused a slight decrease in the MICs from 8 to 32 mg/L and a marked reduction in the MBCs from 32 to 128 mg/L. It was revealed that the extract exhibited a bacteriostatic activity in all the tested isolates at neutral pH, while the activity at pH 5.0 displayed bactericidal effects. The data confirmed that phagolysosomal pH enhanced the pronounced bactericidal activity of the extract against S. aureus.
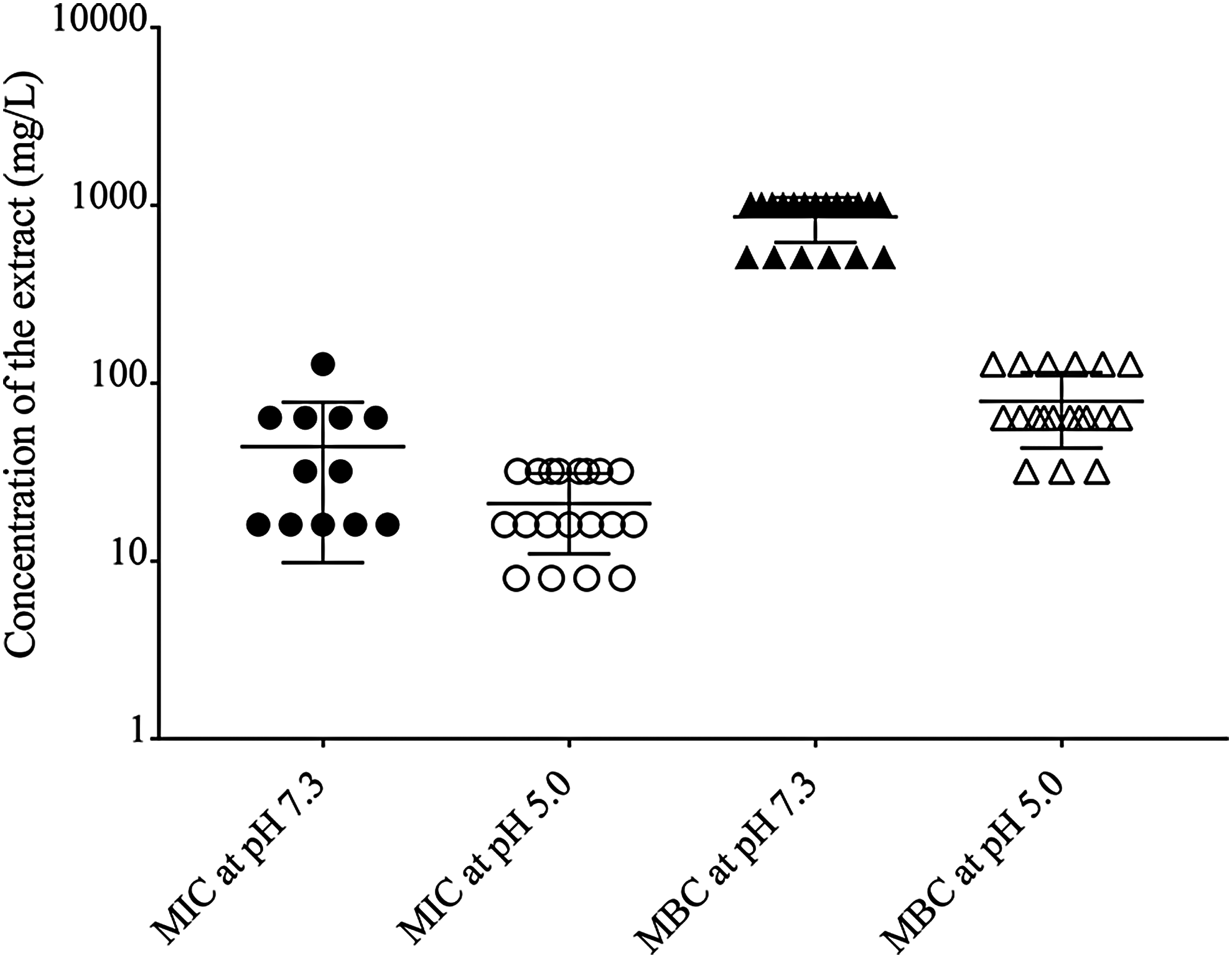
Influence of pH on susceptibility of S. aureus to R. tomentosa extract. The bacterial cells were treated with the extract at concentrations ranging from 4 to 1,024 mg/L in broth at pH 7.3 or 5.0. The diagram presents the MIC and minimum bactericidal concentration (MBC) of the extract against S. aureus ATCC 25923 and ATCC 29213 as well as methicillin-resistant S. aureus (n = 19). The bars indicate medium value in the range of MICs at pH 7.3 (●) and pH 5.0 (◯) or MBCs at pH 7.3 (▲) or pH 5.0 (△). Data were collected from two independent experiments and three technical replicates.
pH 7.3, the conditions to mimic extracellular environment.
pH 5.0, the conditions to mimic phagolysosomal environment.
NA, not applicable.
Effects of R. tomentosa extract on cytotoxicity in HaCaT keratinocytes
To confirm that R. tomentosa extract at the concentration used was not related to cellular toxicity, HaCaT keratinocytes were incubated with increasing concentrations of the extract. The cytotoxicity was assessed by the resazurin reduction assay and the results are presented in Figure 8. The extract at concentrations up to 256 mg/L did not significantly decrease the viability of HaCaT cells at 6 and 48 hr of incubation (≥89% viability). Therefore, it is suggested that the extract at the concentrations up to 16 × MIC had no cytotoxicity toward human HaCaT keratinocytes.

Cytotoxicity of R. tomentosa extract to HaCaT keratinocyte cells. HaCaT cells (2 × 104 cells/ml) were exposed to the extract at concentrations ranging from 4 to 512 mg/L for 6 hr (■) or 48 hr (). The cell viability was determined by resazurin assay and shown as the percentage of viable cells. Data are the mean ± SEM of three independent experiments. *p < 0.05, compared with no extract control. DMSO, dimethyl sulfoxide.
Discussion
Staphylococcal skin infections are initiated by bacterial attachment to epithelial cells and subsequently followed by the invasion. During the bacterial invasion into epithelial cells, inflammatory molecules, including interleukin (IL)-6, IL-8, and antimicrobial peptides, are produced to control the infections.27,28 However, the pathogen can modify itself by producing bacterial biofilm, pigments, and enzymes, as well as by modifying their surfactants, leading to resistance to the antimicrobial molecules, and resulting in persistent and deep infections.2,12,29,30
World Health Organization has reported that 85% of the people in developing countries used plant extracts as a primary traditional medicine in a healthcare setting, as well as the herbs were used in complementary and alternative medicine in many developed countries. 31 Therefore, several studies have been focused on natural products as alternative treatments to enhance host immunity32–34 and inhibit bacterial virulence factors resulting in the infection control.35,36 In this study, R. tomentosa extract was effective in reducing MRSA attachment and invasion into HaCaT keratinocytes. According to our previous study, antistaphylococcal adhesion was observed after exposure to the extract and its purified compound, rhodomyrtone. 22 Certain studies on evergreen shrubs showed that phloroglucinol derivatives were the main active ingredients in plant extracts from the aerial parts.37,38 Hypericum hircinum extract displayed antibacterial activity against S. aureus, but had no activity in its purified components due to the loss of phloroglucinol during the isolation. 37 Hypericum perforatum ethanol extract containing a phloroglucinol derivative, hyperforin, had a good anti-gram-positive activity, whereas the activity was independent of flavonoids. 38 Our previous studies showed that R. tomentosa leaf extract exhibited a strong antibacterial activity against gram-positive bacteria.19,20 The active fractions were further fractionated and confirmed by nuclear magnetic resonance study, which revealed that the phloroglucinol derivative rhodomyrtone was the active component.19,20
Treatment of staphylococcal biofilm infections is very difficult since it can resist antibiotics and host immunity resulting in persistent infections.2,3 Therefore, it requires small molecules to penetrate through biofilm matrix to kill the bacterial cells. Rhodomyrtone, a small molecule (442.55 g/mol), inhibited virulence factors associated with the ability of staphylococcal infections.35,39,40 For instance, the decrease in biofilm formation involving the bacterial adherence was detected. 35 Similarly, autolysin expression associated with the enhanced invasion was reduced after exposure to rhodomytone. 39 Therefore, the reduction of bacterial adhesion and invasion after treatment with R. tomentosa extract may be due to the inhibition of the virulence factors by rhodomyrtone. Moreover, we earlier reported that rhodomyrtone could induce IL-6 expression in response to MRSA infection, which led to the enhanced intracellular killing by human monocytes. 34 The results suggested that the extract might affect the interactions between the bacteria and their host cells, resulting not only in deficient attachment and invasion of the pathogen but also in IL-6 production, which in turn increase antimicrobial peptide production and enhance intracellular killing of S. aureus in keratinocyte cells.27,28
Our previous studies demonstrated that rhodomyrtone could inhibit immunodominant antigen A, 39 superoxide dismutase, 39 and staphyloxanthin 40 produced by S. aureus. In addition, the compound completely inhibited DnaK protein 39 that contributes to the survival of S. aureus, the production of bacterial pigment and autolysin, as well as the increased killing activity of β-lactam drug in MRSA. 41 These virulence factors lead to the increasing intracellular bacterial persistence in host cells since they can resist to lysozyme and oxidative stress such as superoxide as well as hydrogen peroxide.42,43
R. tomentosa leaf extract with several active ingredients possesses antibacterial, 20 antibiofilm,21,35 antioxidant, 44 anti-inflammatory, 45 and antimicrobial adhesion activities,22,23 which may display synergistic effects on treatment of skin infections. In this study, susceptibility of S. aureus to R. tomentosa extract in the presence of acidic pH, hydrogen peroxide, and lysozyme to mimic intracellular environment of host phagocytes was tested. The results indicated that both S. aureus ATCC 29213 and MRSA could overcome the killing activity of hydrogen peroxide and lysozyme. Probably, it is due to the keratinocyte nature, which is nonprofessional phagocytes and has poor intracellular killing activity. On the other hand, an increase in the susceptibility of pathogens to the extract in phagolysosomal pH condition was observed. To confirm an enhanced anti-MRSA activity of the extract under phagolysosomal pH, the intracellular killing assay was performed. The results showed that intracellular MRSA within HaCaT keratinocytes was reduced by treatment with the extract, while in the absence of the extract, the bacterial cells could survive and replicate. Together with our finding, it can be suggested that the extract enhanced the intracellular killing activity of HaCaT cells in correlation with the phagolysosomal pH condition, but not the host oxidative stress. We further investigated effects of R. tomentosa extract on bacterial nitric-oxide synthase produced by S. aureus that plays the role in resistance to host oxidative stress as well as antimicrobial peptides. 30 The results showed that no effect of the extract on the production of the bacterial nitric-oxide synthase was demonstrated (data not shown). This may be a result of the effect of phagosomal acidification on reducing the bacterial surface hydrophobicity, 46 leading to an enhanced antimicrobial activity against the intracellular pathogens by R. tomentosa extract. Similarly, there was evidence that acidic pH within phagocytosis vacuole is required to destroy S. aureus by activating host immune responses. 47 However, the intracellular conditions have been previously reported to interfere with activities of certain antibiotics such as vancomycin, gentamicin, macrolides, and quinolones.25,48 Our results further confirmed that the extract could enhance the intracellular killing activity in phagosomal pH condition by altering cell morphology and inducing cell lysis.
Accessory gene regulator (agr) locus is a global regulator that controls multiple virulence factors of S. aureus, including induction of host cell death. 49 In this study, R. tomentosa extract decreased apoptotic HaCaT cell death caused by MRSA. Moreover, reduction in the keratinocyte death caused by the bacterial supernatant might result from the inhibition of quorum sensing through the agr system by R. tomentosa extract. 21 In addition, clp proteolytic complexes play a role in controlling the agr system and the absence of the molecules displayed a low pathogenicity in mouse skin abscess model. 50 Interestingly, clp proteins have been previously reported to disappear after MRSA exposure to rhodomyrone. 39 Possibly, R. tomentosa extract could inhibit the bacterial cell to cell communication through the agr system leading to the reduction in MRSA colonization and host cell death. Ideally, a deletion of clp protease in S. aureus should inhibit the production of bacterial virulence and stress survival. Virulence factors such as protein A and secreted proteins are controlled through quorum-sensing agr locus involving the expression of clp chaperones during the late bacterial growth phase. 51 We observed reduction in MRSA invasion after exposure to R. tomentosa extract at 18 and 24 hr, as well as decrease in MRSA-secreted protein-induced damage to HaCaT keratinocytes. Together with our findings, the data suggested that R. tomentosa extract might involve in the inhibition of clp protease, resulting in decrease in bacterial cell wall-anchored proteins, which play a role in bacterial invasion and secreted proteins associated with host cytotoxicity. In addition, R. tomentosa at effective concentrations (4–128 mg/L) was not found to be toxic to HaCaT keratinocytes. According to our previous in vivo study using Swiss albino mice, oral administration of R. tomentosa extract at concentration of 2 g/kg body weight/day was safe until 14 days of observation. 44 Moreover, the ethanol leaf extract displayed an IC50 value of 476 mg/L against human dermal fibroblasts. 52
Our results indicated that an acidic pH could enhance the antibacterial activity of R. tomentosa extract against intracellular MRSA. Therefore, the extract may be suitable for the treatment of bacterial infections in acidic compartments, particularly skin, and for the therapeutic use as an economically viable alternative medicine.
Footnotes
Acknowledgments
This work was supported by the Thailand Research Fund (Grant number RTA5880005) and the Royal Golden Jubilee Ph.D. Program (Grant number PHD/0293/2552).
Disclosure Statement
No competing financial interests exist.