
Abstract
Strong evidence of high potency of rhodomyrtone as a promising antibacterial agent against pathogenic gram-positive bacteria has been clearly demonstrated in our previous work. The aim of this study was to provide insight into early action of rhodomyrtone, an acylphloroglucinol, on membrane damage in multidrug-resistant methicillin-resistant Staphylococcus aureus (MRSA). Early effects of rhodomyrtone on the bacterial membrane integrity were detected in a time-course study. Flow cytometry revealed a reduction in green fluorescent emission and increase in uptake of propidium iodide in rhodomyrtone-treated bacterial cells in a concentration- and time-dependent manner. Disruption of cytoplasmic membrane was further monitored by measuring cellular adenosine triphosphate (ATP) and potassium ion (K+). Leakage of both ATP and K+ and significant decrease in intracellular ATP in MRSA were observed following treatment. Pronounced changes in the bacterial ultrastructure and morphology were confirmed by transmission electron microscopy and scanning electron microscopy. Bacterial cell disruption, holes in cell surface, and bulge formations were noted in rhodomyrtone-treated cells. In this study, we provided relevant data to clarify that rhodomyrtone is a bacterial cell membrane-damaging agent. A possible early effect of this novel compound involves bacterial membrane disruption.
Introduction
M
Treating resistant bacterial infections has become extremely challenging for the infectious disease physician. Therefore, there is a continuous need to discover and develop novel therapeutic approaches and prophylaxis strategies to combat these emerging multidrug-resistant strains and potentially life-threatening infectious diseases. 8
Plant extracts from leaves of Rhodomyrtus tomentosa (Aiton) Hassk. (Myrtaceae), an evergreen shrub originating from Southeast Asia, exhibit potent antibacterial efficacy against S. aureus and Escherichia coli. 9 A bioactive compound of the extracts is rhodomyrtone, an acylphloroglucinol, 10 which showed a remarkable antimicrobial activity against a number of gram-positive bacteria such as Enterococcus faecalis, S. aureus, S. epidermidis, Streptococcus spp., S. pyogenes, Propionibacterium acnes, and Bacillus subtilis, including multidrug-resistant organisms.9–13 Subsequent studies revealed that rhodomyrtone was effective in preventing biofilm formation and kills the bacterial cells inside the biofilm of S. aureus 14 and P. acnes.
Recently, Srisuwan et al. 15 have reported that rhodomyrtone could act as immunomodulatory regimens, which enhance innate immune responses against MRSA infections. Chorachoo et al. 16 proposed the potent antibacterial activity of liposomal encapsulated rhodomyrtone for treatment of acne vulgaris caused by S. aureus, S. epidermidis, and P. acnes. A brief report has been published describing the use of rhodomyrtone cream as a tropical agent for S. aureus skin infections in rabbits. 17 The previous results showed that rhodomyrtone has gained high interest as a lead candidate for potential novel antibacterial agents. Elucidating fundamental antibacterial mechanisms of rhodomyrtone is necessary for clinical applications.
We have recently undertaken proteome analysis of MRSA upon rhodomyrtone treatment, which indicated that the purified compound showed pronounced effects on the expression of several proteins relating with some crucial metabolic and biosynthetic pathways of the bacteria. 11 Meanwhile, comparison of gene expression profiling between MRSA cultures with or without rhodomyrtone provides us with a better understanding of MRSA response to the compound. The most involved genes encode essential proteins in metabolic pathways, amino acid metabolism, membrane function, adenosine triphosphate (ATP)-binding cassette transportation, lipoprotein and nucleotide metabolism, and bacterial cell envelope. 18 However, detailed studies on the mode of action of the compound on bacterial cells has not yet been well defined.
To provide a better understanding of the specific and early action of rhodomyrtone and precise time sequence of the action induced on MRSA cytoplasmic membrane, we investigated the role of membrane permeabilization in the mechanism of action of rhodomyrtone using fluorescent dyes, green- and red-fluorescent nucleic acid stain. The leakage of the intracellular components of rhodomyrtone-treated MRSA, including potassium ion (K+) and ATP, was determined. Morphological and ultrastructural studies of the treated cells were further confirmed by electron microscopy.
Materials and Methods
Bacterial strains
MRSA NPRC R001, a common endemic isolate with mecA gene obtained from Hat Yai Hospital, Thailand,5,11 was used throughout this study. MRSA NPRC R001 was used as a representative clinical isolate that has been identified to be resistant to several classes of antibiotics, including β-lactams (oxacillin and penicillin), gentamicin, tetracycline, erythromycin, ciprofloxacin, clindamycin, and trimethoprim–sulfamethoxazole. 5 Bacteria were precultured overnight from glycerol stock stored at −80°C on trypticase soy agar (Hi-Media, India).
Antibacterial susceptibility tests
Rhodomyrtone was isolated and characterized as described earlier.10,19 Minimum inhibitory concentrations (MICs) were examined by broth microdilution method according to Clinical and Laboratory Standards Institute procedures. 20 Two-fold serial dilutions of rhodomyrtone and nisin (Sigma-Aldrich, St. Louis, MO) in a 96-well microtiter plate were prepared in Mueller-Hinton broth (BD Biosciences) to obtain a concentration range of 0.0625–16 μg/ml and 0.5–512 μg/ml, respectively. Bacteria were added to each well to give a final inoculum of 5 × 105 colony-forming unit (cfu)/ml. After incubation at 35°C for 16–18 hr, the extent of inhibition of bacterial growth was determined.
MIC was defined as the lowest concentration of the agent that completely inhibited the bacterial growth. Viable cell counts were obtained using drop plate method. Experiments were performed in triplicate and repeated at least thrice for each assay.
Flow cytometric measurements
To evaluate the membrane integrity of MRSA after treatment, the BacLight™ RedoxSensor™ Green Vitality Kit (Invitrogen, Carlsbad, CA) was used following the manufacturer's guidelines. Bacterial cells were prepared in phosphate-buffered saline (PBS) to the cell density of ∼1 × 108 cfu/ml. The cultures were exposed to rhodomyrtone at 0.5, 1, 2, and 4 MIC or control condition for 30-, 60-, and 240-min incubation. Following treatment, the bacterial cells were washed twice, diluted in PBS to 1 × 106 cfu/ml, and then stained with fluorescence in the dark. The green and red fluorescence intensities of 50,000 stained bacteria per milliliter were recorded and analyzed using flow cytometric analysis. A Cytomics FC 500 flow cytometer (Beckman Coulter, Inc., Brea, CA) filtered with a 488-nm excitation laser was used for membrane permeability analyses.
Green fluorescence was detected on channel FL1 with a 525 nm bandpass filter. Red fluorescence was detected on channel FL3 with a 620 bandpass filter. Nisin was used as a positive control agent. Ethanol-fixed bacteria (70% ethanol for 5 min) were used as a positive control population of death cells, which were permeable to propidium iodide (>80% propidium iodide-stained cells). The fluorescence values were corrected by subtracting the fluorescence of the control samples. Data were confirmed in three independent experiments. Bacterial cell viability after treatment was monitored by viable cell count.
Measurement of ATP levels
Exponential-phase MRSA NPRC R001 at 1 × 108 cfu/ml was treated with 0.5, 1, and 2 MIC rhodomyrtone or 1% dimethyl sulfoxide (DMSO) (untreated control) at 35°C for 30, 60, and 120 min. To assess the amount of intracellular ATP and ATP release, the bacterial cells were harvested by centrifugation (8,000 g at 4°C for 10 min) and, in parallel, supernatants were filter sterilized and transferred into new tubes to determine extracellular ATP as ATP leakage out of the bacterial cells. Bacterial pellets were washed thrice with ice-cold PBS, suspended in PBS, and kept on ice until use. Intracellular ATP was extracted by DMSO.
ATP was measured using an Enliten ATP assay system bioluminescence detection kit (Promega). Luminescence was measured using luminometer PE Applied Biosystems TR717. The assays were calibrated using a standard curve obtained by measuring the bioluminescence of known concentrations of ATP standard solutions.
Potassium (K+) leakage assay
Log-phase growth bacteria were harvested by centrifugation and washed thrice in ice-cold 10 mM HEPES (Sigma, Germany) buffer, pH 7.4. The bacterial cells were then resuspended in the same buffer to obtain a cell initial inoculum of 1 × 108 cfu/ml. The bacterial cultures were treated with rhodomyrtone at 0.5, 1, and 2 MIC and incubated for 30, 60, 90, 120, and 240 min. High concentration of the bacterial cells was to ensure that the leakage of K+ was determined from the viable cells at each assay time. The bacterial cells were harvested by centrifugation. Experiments were conducted in three independent biological replicates. The K+ concentration in the supernatant was measured using an inductively coupled plasma optical emission spectrometer (ICP-OES; Spectro Analytical Instruments, Inc.)
Transmission electron microscopy
Following exposure to rhodomyrtone at 4 μg/ml for 1 or 4 hr, the bacterial cells were washed twice, resuspended in PBS, and fixed with 2.5% glutaraldehyde in PBS (pH 7.3). The samples were incubated overnight at 4°C, centrifuged, and washed twice with PBS. The pellets were fixed with 1% osmium tetroxide for 2 hr and were then left at room temperature for 1 hr. The samples were dehydrated in a series of graded ethanol solutions (70%, 80%, 90%, and 100%), infiltrated in Epon 812 resin, embedded, and left to polymerize overnight at 80°C in an oven. Each sample was cut into thin sections, ∼80–90 nm, using an ultramicrotome, and double stained with 5% uranyl acetate, and lead citrate. Morphology and ultrastructure of bacteria were observed and photographed by JEOL JEM-2010 transmission electron microscopy (TEM) at 80 kV.
Scanning electron microscopy
The bacteria cultures with or without treatment with 16 μg/ml rhodomyrtone were incubated for 1 hr. The bacterial cells were washed twice with PBS and fixed in 2.5% glutaraldehyde in 0.2 M PBS (pH 7.3) for at least 2 hr at room temperature. Samples were dehydrated by washing with increasing ethanol concentrations (50%, 70%, 80%, 90%, and 100%) and then air-dried. The samples were then sputtered using gold in a JEOL FC-110 Ion Sputtering Device and viewed with an FEI Quanta 400 and an accelerating voltage of 5 kV.
Statistical analysis
Statistical analysis of the bacterial counts between test and control groups was performed using one-way analysis of variance. Significance was evaluated at p-value of 0.05.
Results
Antimicrobial activity of antimicrobial agents
Anti-MRSA activity of rhodomyrtone and nisin was measured by broth microdilution method. The MIC and minimum bactericidal concentration (MBC) of rhodomyrtone against MRSA NPRC R001 were 1.0 and 4.0 μg/ml, respectively, which were similar to previously published data. 11 The MIC value of nisin against MRSA NPRC R001 was 256 μg/ml.
Membrane permeabilization of MRSA by rhodomyrtone treatment—fluorescent dye studies
The time course of antimicrobial action of rhodomyrtone at different concentrations against MRSA was further characterized. BacLight Redox Sensor Green reagent (Invitrogen) was utilized to monitor the effect of rhodomyrtone on MRSA membrane permeability and the bacterial vitality. Nisin, a pore-forming antibiotic, was used as a positive control agent. By flow cytometric analysis, the partition of two-color fluorescence among the bacterial cells and assay media was proportional to determine the permeability of the bacterial membrane after exposure to rhodomyrtone. A reduction of the green fluorescent emission and an increase in the degree of propidium iodide in rhodomyrtone-treated MRSA were detected, indicating bacterial membrane damage, which occurred in a concentration-dependent manner (Fig. 1A).

The percentage of membrane-permeabilized cells of MRSA determined by flow cytometry
The compound caused a loss of green fluorescence within 30-min treatment at MIC and higher than MIC. Meanwhile, the bacterial cells treated with 0.5 MIC of rhodomyrtone demonstrated a loss in membrane integrity after 60 min. Nisin presented a potent effect on the bacterial membrane at all time points. Viable cell count was assessed by enumeration on agar plates to confirm the killing of MRSA cells under conditions performed in the membrane integrity assays (Fig. 1B). The correlation between membrane permeability and bacterial cell death was observed. A decrease in viability induced by both rhodomyrtone and nisin was accompanied by complete membrane damage detected by fluorescent dye analysis. The results clearly showed that rhodomyrtone at all tested concentrations permeabilized MRSA cytoplasmic membrane in the population.
Rhodomyrtone causes ATP leakage
To further monitor the physiological consequences of membrane disintegration and the bacterial membrane functions, intracellular ATP and ATP leakage from the MRSA cells were measured at time intervals following the addition of rhodomyrtone by luminescence-based assay. Rhodomyrtone-treated cells showed a remarkable reduction of intracellular ATP when compared with untreated control. The action was accompanied by both concentration- and time-dependent manner after 30- and 60 min-treatment. No significant difference of intracellular ATP was detected in the 120 min-treatment among all rhodomyrtone concentrations (Fig. 2A). Meanwhile, release of extracellular ATP after treatment was investigated. Increases in extracellular ATP of the bacteria in a concentration-dependent manner were observed after exposure to the compound for 30 and 60 min (Fig. 2B).
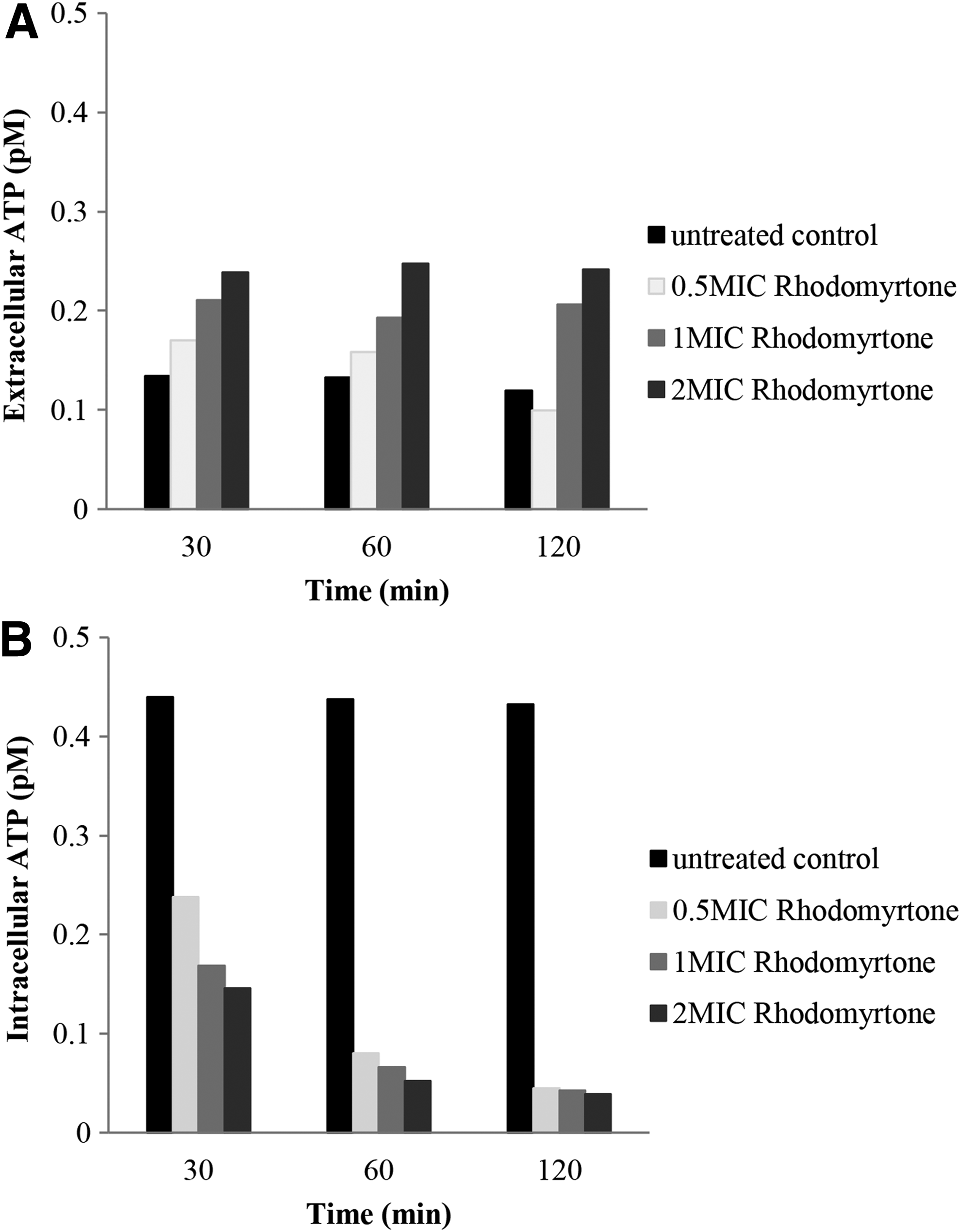
Intracellular
Release of cellular potassium after treatment
K+ leakage from rhodomyrtone-treated MRSA cells compared with untreated cultures was examined over 240 min at different exposure times. K+ started to leach out from the bacterial cells within the first 30 min after treatment at all concentrations of rhodomyrtone and remained constant until 240 min. The leakage amount was not in a rhodomyrtone concentration- and time-dependent manner. However, more release of K+ was observed in the treated cultures at all concentrations when compared with the control (Fig. 3).
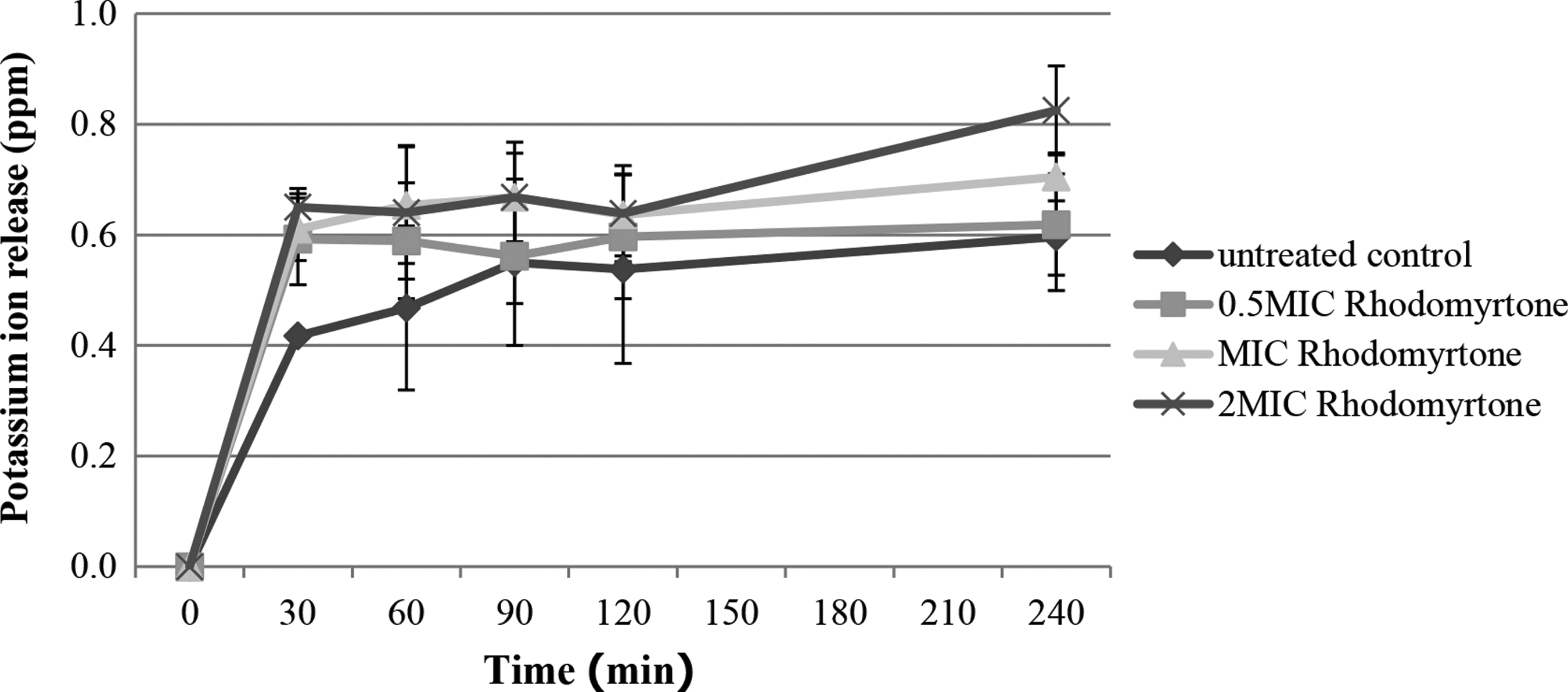
Potassium leakage of MRSA cultures after exposure to rhodomyrtone. Values were SD of the means (SD) from three independent determinations; p-value <0.05, rhodomyrtone-treated cultures compared with untreated control. The amount of extracellular potassium of rhodomyrtone-treated cells was subtracted at each time point from that of untreated cells.
Transmission electron microscopy
Untreated MRSA cells displayed a normal cell shape with well-defined membranes, undamaged structure, and morphology (Fig. 4A). The cultures with 4 μg/ml of rhodomyrtone showed loss of cell wall and cell wall shreds after a 1-hr incubation (Fig. 4B), and enlargement of cells with abnormal initiation of septum formation (Fig. 4C). The bacterial cytoplasmic membrane mostly lost its integrity and showed irregularity cytoplasmic components (Fig. 4B–D). Some cell lysis and an undefined septum were revealed (Fig. 4B). Detachment of the cell wall from the cytoplasmic membrane and shrinking cells were clearly visible following the challenging of rhodomyrtone for 4 hr, as demonstrated in Fig. 4E and G. Damage of cell walls and septum was occasionally found (Fig. 4F). The cytoplasmic region appeared to be a nonhomogeneous appearance of mesosome structure (Fig. 4F, G) when compared with the control. However, no notable lysis cell was observed after treatment for 1 and 4 hr.

Transmission electron micrographs of rhodomyrtone-treated cells. MRSA NPRC 001R cultures with addition of 4 μg/ml of rhodomyrtone were incubated for 1 hr
Scanning electron microscopy
Untreated S. aureus displayed typical round to oval cocci with smooth and intact surface as a sign of morphological integrity (Fig. 5A). Broken cells were observed in cultures after exposure to 16 μg/ml of rhodomyrtone for 1 hr of incubation (Fig. 5C). This culture condition induced protrusion in the bacterial cell surface and septum (Fig. 5B, D), and showed a smaller cell size (Fig. 5D). No obvious difference in the bacterial cell surface architecture between rhodomyrtone-treated cells and rhodomyrtone-untreated control was observed (Fig. 5A–E). Certain treated cells showed holes (Fig. 5E), bust in their cell envelope (Fig. 5C), and complete lysis cells (Fig. 5E).
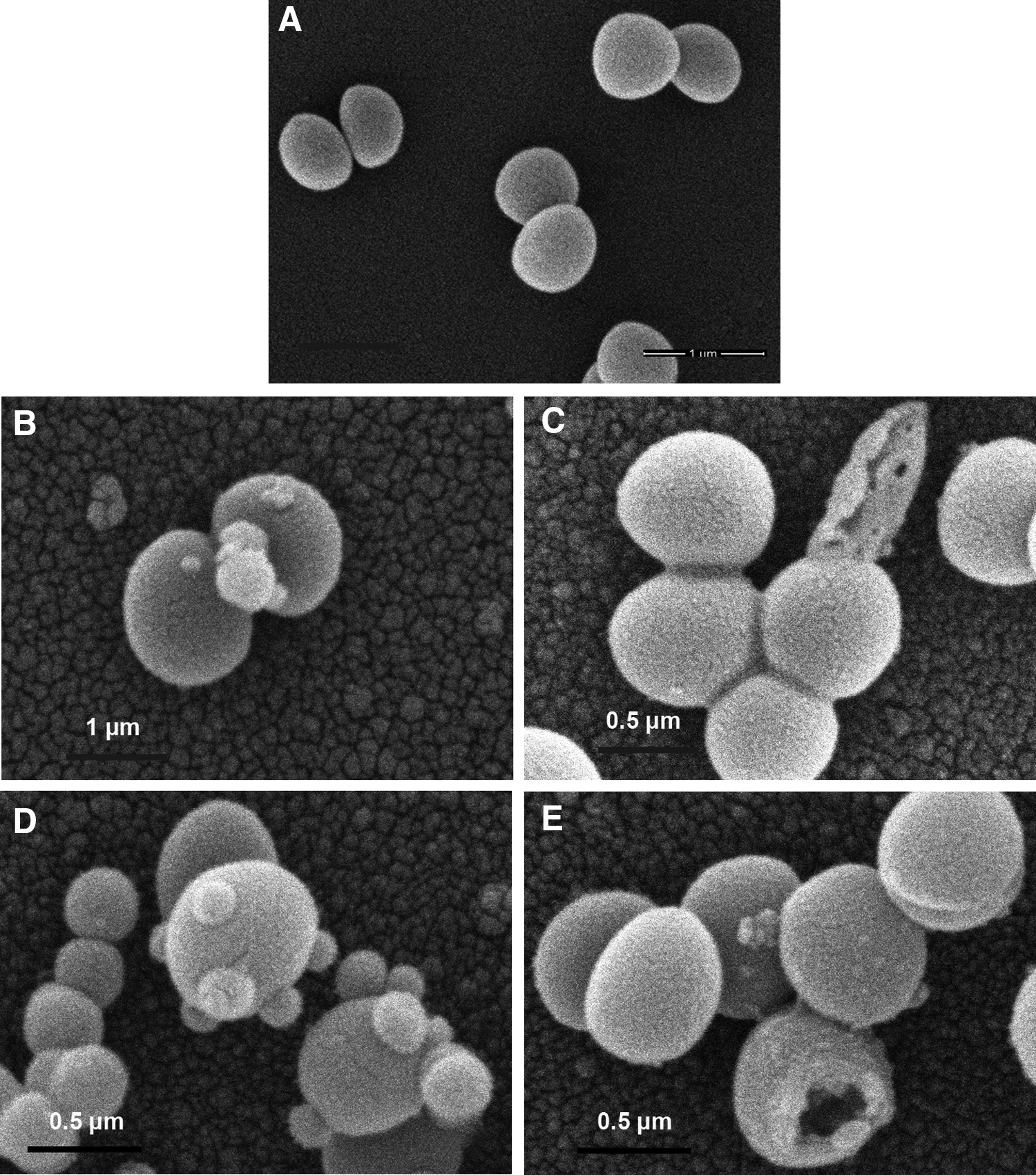
Scanning electron micrographs of rhodomyrtone-treated cells. MRSA NPRC 001R was grown in Mueller-Hinton broth containing 16 μg/ml rhodomyrtone for 1 hr
Discussion
Rhodomyrtone has been defined as an acylphloroglucinol derivative with an excellent in vitro antibacterial activity against several important antimicrobial-resistant organisms.10,12 Moreover, our research group reported the attempt and application of the plant crude extract and rhodomyrtone as a new alternative compound to reduce bacterial infections, such as ex vivo study in a bovine udder epidermal tissue model, 21 and an antiproliferative and apoptosis-inducing agent in HaCaT cells. 22
The investigation of the effect of the compound at early time points could provide useful information to support the use of rhodomyrtone as an alternative antimicrobial product. In some recent studies by our group,10–12,18,23 antimicrobial activity testing by broth microdilution and time-kill analysis of rhodomyrtone against several pathogenic gram-positive bacteria (clinical and reference strains) showed similar results with MIC values ranging from 0.5 to 2 μg/ml. In this study, MRSA NPRC 001R was used as a representative clinical isolate of multidrug-resistant MRSA obtained from Hat Yai Hospital, Thailand.
To evaluate the early effects of rhodomyrtone on the bacterial cytoplasmic membrane, double-color fluorescence using flow cytometric approach was used. In the bacterial cultures treated with or without rhodomyrtone, redox sensor green dye was used to evaluate metabolically active cells, while red fluorescence, propidium iodide, a marker for membrane damage, was utilized to monitor whether the bacterial cells were alive or dead. It is postulated that rhodomyrtone may exert its inhibitory effects by increasing the membrane permeability causing leakage of the bacterial cellular contents within 30 min of treatment.
Membrane potential of microorganisms is generated and maintained by concentration gradients of ions such as sodium, potassium, chloride, magnesium, and hydrogen. 24 If the bacterial membrane becomes compromised, the small ions tend to leach out. Therefore, the determinations of the intracellular components released such as ATP and K+ 25 were also applied to monitor membrane damage caused by rhodomyrtone. Measurement of ATP content in microorganisms can be related to viability, metabolic viability, or by utilizing an average concentration or amount of ATP inside the cells.
Potassium is a significant intracellular ion in many bacteria and is actively imported into cells. 26 Potassium leakage from microorganisms that were exposed to agents was used to estimate the membrane-damaging effects. Upon addition of agents, the alterations of intracellular and extracellular ATP balance in the bacteria were observed such that the ATP is lost through the disrupted cell membrane. The study demonstrated that the intracellular ATP decreases along with a minor ATP release. In contrast, Leejae et al. 12 reported no significant change in the leakage of S. aureus cellular components, which was measured by the change of A260nm after treatment for 4–24 hr. After 18 hr of treatment, the major localization of rhodomyrtone is on S. aureus cytoplasmic substance, while a small amount of rhodomyrtone was detected at cell wall and cell membrane. 12
Electron microscopy is important as a complementary technique to confirm the early action of rhodomyrtone, by demonstrating both cell surface effects and intracellular alterations after treatment. There was some information available in scanning electron microscopy (SEM) analysis of MRSA exposed to rhodomyrtone with no profound change in bacterial morphology and structure between treated cells and untreated control due to the treatment at sub-MIC. 18 Meanwhile, late effect of rhodomyrtone at 0.174 μg/ml against Staphylococci after overnight incubation was observed on the bacterial cell envelope. Phase-contrast microscopic analysis showed that the compound at 5 MIC demonstrated only insignificant morphological effects on Streptococcus mutans; indistinct enlarged cells have been detected after treatment. 10
Ethanolic extract of R. tomentosa at MBC value caused some disorders during cell division step with irregular shape and different size on S. pyogenes cells after treatment for 14 hr. 27 In this study, early effects of rhodomyrtone at 4 and 16 μg/ml after 1 and 4 hr of incubation on MRSA morphology and ultrastructure were clearly visualized using TEM and SEM. Several distinct appearances of the bacterial cellular damage following treatment were observed. Untreated cells appeared round and intact under microscopes. TEM studies on rhodomyrtone-treated MRSA showed mesosome-like structures and detached cell walls. These observations were also reported on S. aureus after exposure to defensin, 28 a cationic microbicidal peptide. 29 Mode of action of defensins is a consequence of the disruption of microbial membrane integrity and function.
The formation of mesosome, membrane-bound organelle, inside microorganism cells also occurred after exposure to many antimicrobial agents and compounds such as trimethoprim,
30
rifampicin, monohydrochloride of
S. aureus treated with supra-MIC of rhodomyrtone developed mesosome-like structures, but no remarkable changes were observed in the bacterial cell walls. Changes in the cell walls were not as prominent as those found in Staphylococci exposed to some antibiotics such as beta-lactam antibiotics that inhibit cell wall synthesis and cause marked thickening of staphylococcal cross walls. 32 Furthermore, many distinct signs of damage of cell envelope were clearly demonstrated by SEM microscopy, such as blisters, numerous small spherical features, some holes and deep craters, and burst cells. Bulge formation was noted on rhodomyrtone-treated MRSA, the influences similar to those of some membrane-disrupting antibiotics at bactericidal concentration. 33
In conclusion, the results improve our understanding of the novel additional action of rhodomyrtone on multidrug-resistant MRSA membrane. Rhodomyrtone induces the disruption of the bacterial membrane associated with a leakage of K+ and ATP out of the cells. However, previous reports suggested that the compound might kill bacteria by plural effects. 18 Rhodomyrtone, an anti-MRSA agent, exhibited bactericidal activity caused by a reduction in the bacterial viability with substantial membrane-damaging effects. Therefore, this study presents experimental evidence that supports further development of rhodomyrtone as a novel therapeutic agent against MRSA infections.
Footnotes
Acknowledgments
The research was supported by a grant from Prince of Songkla University, Contract No. SCI550154S, and partly supported by TRF Senior Research Scholar (Grant No. RTA5880005), the Thailand Research Fund.
Disclosure Statement
No competing financial interests exist.