
Abstract
Two basic questions about lysozyme activities on the selected microorganisms were investigated, namely whether lysozyme inhibits biofilm production and which concentrations of the enzyme have the ability to change the natural biofilm producing capacity of different strains of Staphylococcus aureus (methicillin sensitive and resistant), Streptococcus pyogenes, Pseudomonas aeruginosa, and Gardnerella vaginalis. The effect of lysozyme on biofilm formation capacities of 16 strains of selected microorganisms was investigated, whereby four testing replicates have been performed in vitro using the Test Tube method, and the potential of lysozyme to change biofilm forming capacities depending on its concentration, species, and strains of microorganisms is demonstrated. A lysozyme concentration of 30 μg/ml indicated to have the highest inhibiting effect on all tested microorganisms. Furthermore, G. vaginalis was the most sensitive of them all, as its biofilm formation was inhibited in the presence of as low as 2.5 μg/ml of lysozyme. At enzyme concentrations of 7.5–50 μg/ml (with the exception of 30 μg/ml) the biofilm forming capacities of P. aeruginosa were enhanced. Depending on the strain of P. aeruginosa, the total biofilm quantity was either reduced or unaffected at lysozyme concentrations of 2.5, 5, 7.5, and 30 μg/ml. In contrast, lysozyme concentrations below 15 or 20 μg/ml did not affect or increase the volume of biofilm formation, while higher concentrations (15, 20, 25 μg/ml) reduced biofilm formation by 50% (3/6) and 30 μg/ml of biofilm reduced biofilm forming capacity of S. aureus by 100% (6/6). The results of this study are a strong foundation for further research on lysozyme as a modulator of the biofilm forming capacity of different species with the potential to aid in the development of new drugs for the treatment of oral and vaginal infections.
Introduction
C
Recent studies have shown that cell–cell communication and quorum sensing are crucial for understanding bacterial behavior as it relates to biofilm formation and infection. Quorum sensing, also known as quorum quenching, is the regulation of gene expression in response to fluctuations in cell-population density, in which quorum sensing bacteria produce and release signal molecules, or autoinducers, whose concentration increases as a function of cell density.3,4 Furthermore, new paradigms in signal-mediated cell–cell communication have been initiated through recent studies that demonstrated that signal-mediated communication is not restricted to interactions between the same, but also between different bacterial species, as well as between bacteria and host cells.
A promising strategy to combat infections is the inhibition of quorum quenching. Many molecules that regulate a number of traits, including virulence and biofilm formation, have been identified as signals in the bacterial cell–cell communication.9,10 Considering the significance of quorum quenching in the potential prevention of infection, especially at a time of increasing antibiotic resistance, efforts in the search for novel quorum quenching signaling molecules are ongoing.
Human lysozyme (muramidase or N-acetylmuramic hydrolase) is a secretory ubiquitous enzyme that can be found in tears, milk, saliva, liver, and plasma as part of the innate defense system of the human body.11–13 A well known antimicrobial mechanism of lysozyme is its enzymatic activity in the bacterial peptidoglycan where it hydrolyses of β-1,4 glycosidic bonds between N-acetylglucosamine and N-acetylmuramic acid. In addition, there is an on-lytic mechanism of the enzyme activity that is related to its cationic and hydrophobic properties, which results in bacterial autolysis. 14 Furthermore, bacteria have developed their own defense mechanisms against lysozyme action. Two major mechanisms are behind the ability of bacteria to resist the antimicrobial action of lysozyme, namely protection of the bacterial outer membrane and the production of chemical variants of peptidoglycan that preclude the binding of lysozyme to peptidoglycan. Staphylococcus aureus, an important human pathogen, is known for its resistance to lysozyme through the modification of the peptidoglycan with O-acetyltransferase. Widely distributed among pathogenic strains, the gene coding for O-acetyltransferase (oatA) is responsible for the O acetylation of the N-acetylmuramic acid C-6 hydroxyl group. 15
Any suppressed effect of a bacterial infection is worth investigating, which is of particular relevance today, at a time when the World is confronted with vast antibiotic resistance and the rise of persistent biofilms.6–8 A new approach to monitoring the microbial community imposed a new strategy of studying the effects of antimicrobial and antibiofilm forming substances. To evaluate the possibility of its therapeutic use, alone or in combination with other antimicrobial substances, the need to study effects of lysozyme on biofilm forming capacities of microbial communities is highlighted. Two basic questions about lysozyme activities on the selected microorganisms were investigated, namely whether lysozyme is an inhibitor of biofilm production and which concentrations of lysozyme change the natural ability of different strains of S. aureus (S. aureus, including methicillin-sensitive [MSSA]), methicillin-resistant S. aureus (MRSA), Streptococcus pyogenes, Pseudomonas aeruginosa, and Gardnerella vaginalis to produce biofilm.
Materials and Methods
The study was conducted during the period from May 1st until June 15th, 2016 at the Institute for Biomedical Diagnostic and Research NALAZ (Sarajevo, Bosnia, and Herzegovina).
Materials
A total number of four bacterial species were used, namely MSSA, MRSA, G. vaginalis, P. aeruginosa, and S. pyogenes. A total of 16 bacterial strains of microorganisms and 5 quality control strains purchased from the American Type Culture Collection (ATCC) and 11 hospital strains (isolates from the patient library of the Institute for Biomedical Diagnostic and Research NALAZ) were included in the study (Table 1). Four testing replicates have been performed in vitro using the Test Tube Method (TTM). 16
ATCC, American Type Culture Collection; MRSA, methicillin-resistant S. aureus; MSSA, S. aureus, including methicillin-sensitive.
Methods
Cultivation of microorganisms
S. aureus, S. pyogenes, MSSA, and MRSA were cultivated on Columbia agar in combination with 5% sheep blood (bioMèrieux); G. vaginalis was grown on selective Gardnerella agar medium (bioMèrieux); P. aeruginosa was grown on chromID CPS selective agar (bioMèrieux). Organisms were incubated at 37°C for 24 hours except G. vaginalis and S. pyogenes, both of which were incubated in a carbon dioxide (5%) enriched atmosphere for 48 hours.
Preparation of inoculums for experiments
Inoculums were prepared as previously described. 17 Briefly, overnight cultures of 16 selected strains were used to prepare a cell suspension in Trypticase Soy Broth (TSB) with 1% glucose medium (BD) to an optical density at 600 nm (OD600) of 0.05. Suspensions were incubated at 37°C with 5% CO2 until the culture reached an OD600 of ∼0.2. Glycerol was added to a final 10% (v/v) and stored at −80°C until use. Aliquot amounts of 7 × 105 colony-forming unit (CFU)/ml were used as inoculums for the TTM.
Investigation of the ability of microorganisms to form biofilms
We detected biofilm formation using the TTM. The strains were tested for biofilm production by modification of a previously described standard method. 16 Briefly, an aliquot of each bacterial strain (7 × 105 CFU/ml) was inoculated in four replicates into plastic tubes containing 5 ml of TSB culture with 1% glucose medium and incubated for 16 hours with shaking at 37°C with 5% CO2. Four test tubes that contained TSB without bacterial inoculums were used as negative controls. After the incubation period, the content was decanted, and tubes were washed with sodium phosphate buffer (0.2 M, pH 7.3). Tubes were then filled with 2 ml of 0.1% crystal violet stain and left at room temperature for 5 minutes. After staining, tubes were washed with deionized water thrice and left to dry overnight in an inverted position. Total biofilm amounts that were formed either at the bottom of the tube or along the tube walls were subsequently scored as weak (1), moderate (2), or strong (3). 18
The biofilm forming capacity was further confirmed spectrophotometrically at OD600. The cutoff OD for the spectrophotometric determination of biofilm formation was calculated as three standard deviations above the mean OD of the negative control. Classification of bacterial adherence was calculated as per given formulas for all tested bacteria as one of three possible categories: weak (ODc < OD ≤2OD), moderate (2ODc < OD ≤4OD), and strong (4ODc < OD) biofilm formation.
Analysis of lysozyme effect on biofilm formation
Effect of lysozyme on biofilm forming capacities of microorganisms that previously formed biofilm in the absence of the enzyme was analyzed using the aforementioned TTM. Strains were treated as before in the presence of various concentrations of the lysozyme of egg albumen (Lysozyme Manufacturing Company B.V.) reconstituted in sodium phosphate buffer (0.2 M, pH 7.0). The initial concentration of lysozyme in the sodium phosphate buffer was 70 μg/ml and was adjusted to the desired concentration by adding TSB. Tested concentrations were 2.5, 5, 7.5, 10, 15, 20, 25, 30, 40, and 50 μg/ml.
Statistical analysis
After different lysozyme concentrations were applied to the species individually, the result was analyzed by the HCE 3.5 program to find potential correlation among them (www.cs.umd.edu/hcil/hce). The program hierarchically clustered the results by Pearson's correlation method.
Results
According to the results obtained through the TTM, 10 out of the 16 tested microorganisms formed biofilm (Table 2). Various biofilm intensities were observed, ranging from weak (P. aeruginosa 3989) to moderate (G. vaginalis 4251, G. vaginalis 14018, MRSA 4290, MRSA ATCC 43300, and MRSA 7958) to strong (P. aeruginosa ATCC 10145, MSSA 3129, MSSA 3202, and MSSA 3906) (Table 2). In contrast, no biofilm production by S. pyogenes (hospital strains 3025, 3185, 4317, and ATCC 19615), P. aeruginosa 4228, and MSSA ATCC 25923 was observed. Bacterial strains that produced biofilms were subsequently treated with various lysozyme concentrations, and different biofilm forming capacities were observed (Table 2).
Amount of biofilm estimated: absence = 0, weak = 1, moderate = 2, and strong = 3.
The results also showed that all MRSA strains expressed the same biofilm forming potential when no lysozyme was added; however, fluctuations occurred as we increased the concentrations of lysozyme in the growth medium. Namely, at the concentration of 2.5 μg/ml of lysozyme two MRSA strains expressed higher biofilm forming capacity (which increased from moderate to high), while at the concentration of 7.5 μg/ml all strains expressed high biofilm formation. The biofilm forming capacity started to decrease at a lysozyme concentration of 15 μg/ml for one of the strains and at 30 μg/ml for all the tested strains.
All tested MSSA strains had strong (3) biofilm forming capacities in the absence of lysozyme. At 2.5 μg/ml of the enzyme, two of three strains already demonstrated a decrease [strong (3) to moderate (2)] in the production of biofilm. Furthermore, biofilm formation variation was observed among three strains in the presence of different enzyme concentrations, but the overall trend in production was that biofilm formation decreased with increasing concentrations of lysozyme (Table 2 and Figs. 1 and 2). Finally, all three strains expressed lower biofilm formation in the presence of 30 μg/ml of enzyme.

Detection of biofilm formation capacity in the absence and presence of various concentrations of lysozyme using the Test Tube Method (four testing replicates have been performed for each strain of microorganism, and average capacity values are shown; N = 2 for Pseudomonas aeruginosa, N = 3 for MSSA and N = 3 for MRSA; N = 1 for Gardnerella vaginalis). MRSA, methicillin-resistant Staphylococcus aureus; MSSA, S. aureus, including methicillin-sensitive.
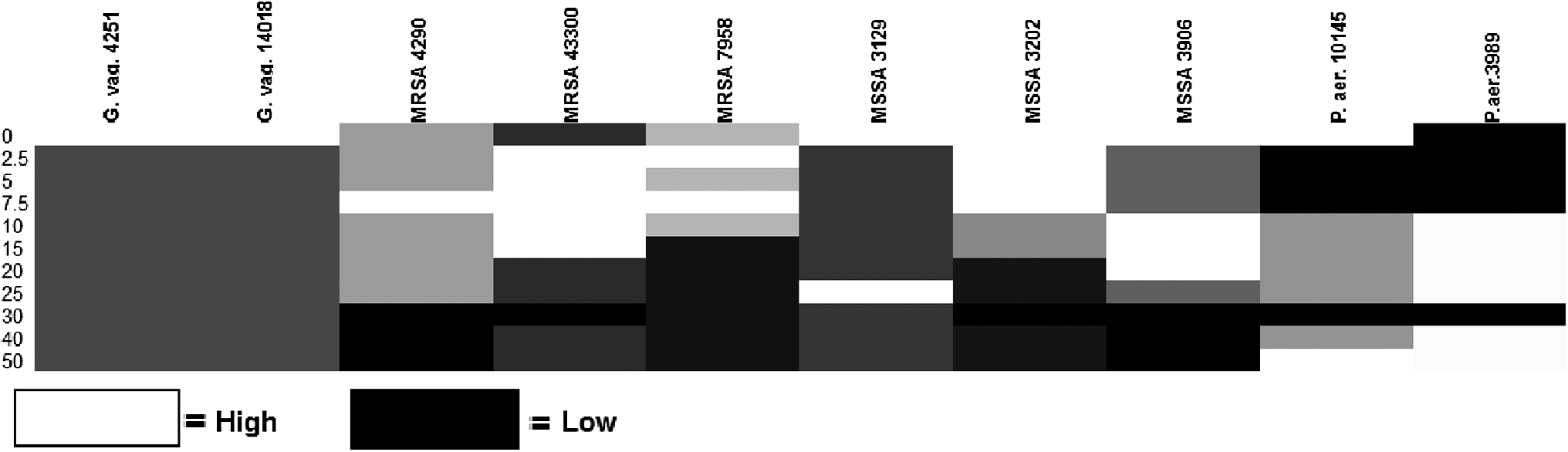
Hierarchical clustering of biofilm formation.
S. aureus had the same biofilm forming capacity when no lysozyme was added. Fluctuations in the biofilm forming capacity started occurring at a concentration of 2.5 μg/ml of lysozyme, and this was observed at concentrations up to 20 μg/ml at which the strains expressed moderate and high biofilm forming capacities.
In the absence of lysozyme, P. aeruginosa 10145 produced strong (3), while P. aeruginosa 3989 produced weak (1) biofilm. The total quantity of P. aeruginosa biofilm was reduced or was not affected, depending on the strain of P. aeruginosa, at lysozyme concentrations of 2.5, 5, 7.5, and 30 μg/ml (Table 1). At enzyme concentrations of 7.5–50 μg/ml the biofilm forming capacities of both strains of P. aeruginosa were enhanced, except at concentration of 30 μg/ml (Fig. 2).
For the two tested P. aeruginosa strains, biofilm forming potential was not the same when no lysozyme was added (one strain expressed high biofilm formation, the other weak). At concentrations of 2.5–7.5 μg/ml both strains expressed weak biofilm forming capacities, while moderate biofilm formation was observed at concentrations of 10–25 μg/ml of lysozyme.
Average values of biofilm forming capacities of MRSA (N = 3), MSSA (N = 3), and P. aeruginosa (N = 2) in the presence of listed lysozyme concentrations through the TTM are illustrated in Fig. 1 (except for G. vaginalis with an N = 1).
There seems to be a strain-specific lysozyme concentration threshold after which the biofilm formation potential decreases. For example, the lysozyme concentration thresholds for MRSA 4290, MRSA 7958, MSSA 3202, and MSSA 3906 are 25, 10, 15, and 25 μg/ml, respectively (Table 2).
Variation of the biofilm formation potential for all tested strains can be seen in Fig. 2. In the absence of lysozyme, the two tested strains of G. vaginalis produced moderate biofilm (2), while in the presence of all tested concentration of lysozyme, they did not produce biofilm. In the presence of 10 μg/ml of lysozyme, P. aeruginosa 10145 demonstrated an increase in biofilm production. Unlike P. aeruginosa, all strains of S. aureus (MSSA and MRSA) showed a decrease in biofilm formation with increasing concentration of lysozyme. Finally, S. aureus demonstrated lysozyme concentration dependent variations in the production of biofilm.
Discussion
Several studies confirm the protective function of the innate immune system against biofilm infections as they point out the antibiofilm activity of lactoferrin, which, together with lysozyme and peroxidase, is a ubiquitous and abundant constituent of human external secretions.19–22 A well-known effect of lysozyme is that it cleaves specific bonds in the peptidoglycan layer, but other possible ways of its antimicrobial activity, such as its signaling molecule capabilities, are not sufficiently explored.11–14,23
Using the TTM, we found that lysozyme affects biofilm forming capacities of each of the bacterial strains that formed any type of biofilm in the absence of it. G. vaginalis 4251 lost its capacity to form biofilm, and it can therefore be speculated that it is the most sensitive of all strains to lysozyme. Furthermore, the biofilm formed by G. vaginalis was reduced even with additions of the smallest concentration of lysozyme. It is of relevance to mention that a disturbed vaginal balance can lead to adverse outcomes for women and their neonates.24,25 G. vaginalis is predominant in bacterial vaginosis, the most common vaginal infection worldwide. It can cause an increased shift in vaginal pH, and hence, lysozyme can function properly in this environment. In addition, synergetic effects of lysozyme and antibiotics against infections were observed in vitro on G. vaginalis biofilm models. 26 Our findings highlight the mediating effect of lysozyme in vaginal infections, which is a strong starting point for the development of new treatments. 25
P. aeruginosa is an opportunistic pathogen which rarely causes disease in healthy individuals, but it can cause life-threatening infections in immunocompromised patients. Under high treat of infections with P. aeruginosa are those who, among other conditions, suffer from burn injuries, cystic fibrosis, respiratory diseases, and HIV infections. P. aeruginosa developed many antibiotic resistance mechanisms, one of which is conferred onto them due to their protective biofilm.27,28
P. aeruginosa has the ability to cause lethal infections that are difficult to eradicate due to their high levels of antibiotic resistance mechanisms, one of which is conferred onto them due to their protective biofilm.27,28 This makes it one of the leading causes of inhospital infections. According to the TTM, the previously strongly forming P. aeruginosa 10145 has a decreased capacity to form biofilms in the presence of 2.5–7.5 μg/ml of lysozyme. In contrast, those lysozyme concentrations had no effect on the initially weakly forming hospital strain. For both strains, the biofilm forming capacity began to increase at concentrations of 10–25 μg/ml and they decreased at 30 μg/ml. Biofilm forming capacity of P. aeruginosa increased again in the presence of 40 and 50 μg/ml of lysozyme, and it remained unchanged for P. aeruginosa 3989. Interestingly, however, the intensity of the biofilm of the former is the same as the starting intensity, whereas that of the clinical strain doubled at the highest enzyme concentration. It is important to note that the increase in biofilm formation capacity in the presence of small concentrations of lysozyme can be alarming, since lysozyme is a well known natural preservative used in food processing industries and accumulation of its biofilms is surely not a positive outcome in this case.13,29
S. aureus, including MSSA and MRSA, is an important human pathogen that forms biofilms in the nasopharynx, anterior nares, and the skin of healthy individuals, but can also promote disease in almost any tissue in the human body.30,31 The successful invasion of the system by S. aureus is regulated by peptide quorum sensing which is crucial for proper biofilm formation. 32 However, a recent investigation demonstrated that S. aureus biofilm induces the lysozyme expressions to a higher level than planktonic cells, indicating that the S. aureus biofilm affects the innate immune system. 33 Biofilm-associated MSSA and MRSA have a special clinical relevance because they show an innate resistance to antimicrobials and clearance by host defenses and they likely contribute to the persistence and resistance to treatment of staphylococcal infections. 32 According to another study, 5 μg/ml lysozyme resulted in no effect on suppression of biofilm formed by S. aureus and no inhibition of growth of the selected strains. 21 This is in contrast with our results, as all hospital strains of MSSA demonstrated a strong ability to form biofilms while MSSA ATCC did not have this capability.
MRSA is the most prominent biofilm-forming human pathogen causing both healthcare-related and community-acquired infections. 32 In this study, all MRSA strains produced biofilm at weak or moderate levels. All of the previously moderately forming MRSA strains lost the capacity to form biofilms at 30 μg/ml of lysozyme, whereas in the presence of lower enzyme concentrations, their biofilm forming capacity was either preserved or enhanced.
Previous work has demonstrated the differential effect of lysozyme against various bacterial species, whereby it was found that lysozyme has a mean inhibitory concentration of <500 μg/ml on the growth of Escherichia coli. 22 In our work, the concentration of 30 μg/ml was the most efficient inhibitor of biofilm formation for all tested strains of microorganisms (G. vaginalis, MSSA, MRSA, and P. aeruginosa). The differences between our results and those found previously 22 could be due to the difference in species of tested microorganisms, as well as in experimental design.
To our knowledge, this is the first study that summarizes the effect of lysozyme on a wide spectrum of hospital and reference strains of microorganisms. Lysozyme has seemingly been neglected in the past, and enormous needs for antimicrobial agents are currently highlighted.
Results of this investigation of lysozyme, a natural prominent antimicrobial agent, point to the need for further research to exploit its lytic and biofilm forming properties, either alone or in combination with other active substances, to contribute to prevention or treatment of bacterial infections.
Footnotes
Acknowledgment
The authors gratefully thank Bosnalijek d.d. Sarajevo for their technical and professional help and expertise.
Disclosure Statement
No competing financial interests exist.