
Abstract
The aim of the present study was to screen for the presence of carbapenemase-producing Enterobacteriaceae (CPE) isolates from wild boars and Barbary macaques in Algeria. Fecal samples were collected from wild boars (n = 168) and Barbary macaques (n = 212), in Bejaia, Algeria, between September 2014 and April 2016. The isolates were identified and antimicrobial susceptibility was determined. Carbapenem resistance determinants were studied using PCR and sequencing, while clonal relatedness was performed using multilocus sequence typing (MLST). PCR was used to investigate certain virulence genes. Three CPE isolates from three different samples (1.8%) recovered from wild boars were identified as Escherichia coli (two isolates) and Klebsiella pneumoniae (one isolate). These isolates were resistant to amoxicillin, amoxicillin–clavulanate, tobramycin, ertapenem, and meropenem. The results of PCR and sequencing analysis showed that all three isolates produced the OXA-48 enzyme. The MLST showed that the two E. coli isolates were assigned to the same sequence type, ST635, and belonged to phylogroup A, whereas K. pneumoniae strain belonged to ST13. The K. pneumoniae strain was positive for multiple virulence factors, whereas no virulence determinants were found in E. coli isolates. This is the first report of OXA-48-producing Enterobacteriaceae in wild animals from Algeria and Africa.
Introduction
I
The blaOXA-48 gene was firstly identified in a K. pneumoniae isolate from Turkey. 6 Since then, several other OXA-48-producing isolates of various Enterobacteriaceae have been reported in North African countries and many European countries.5,7,8 Reports on carbapenemase-producing bacteria in animals are rare, but are on the increase.9–13 The carbapenemase producers were isolated from companion animals.14–17 However, much effort has been made to improve our knowledge on the resistance in human populations and domestic animals, but less attention was devoted to wildlife. Nevertheless, the occurrences of multidrug-resistant pathogens in wild animals, which have not been directly exposed to antibiotics, have been reported through different geographical areas. 3 This indicates the possible transmission between humans, domestic animals, the natural environment, and wildlife. 3
Fischer et al. speculated that wildlife can be a reservoir for carbapenemase-producing bacteria. 9 Since then, some studies have taken place.18–20 In terms of wild animals in Algeria, only one publication describing OXA-23-producing Acinetobacter baumannii from wild fish has been reported. 21 Reports on drug resistance in wild boars (Sus scrofa) and Barbary macaques (Macaca sylvanus) are limited: a review of existing databases resulted in only three reports that were published on extended spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from wild boars in Czech Republic and in Portugal22,23 and one report about ESBL-producing E. coli and K. pneumoniae isolated from both wild boars and Barbary macaques in Algeria. 24 In contrast to ESBL-producing Enterobacteriaceae, no carbapenemase-resistant bacteria were isolated from wild boars and Barbary macaques. Wild boars (S. scrofa) are a species found worldwide, which can thrive in areas that are influenced by human activity. 25 The Maghreb race of wild boar in this study is known as S. scrofa barbarus, and is a native to North Africa. 26 These groups of wild boars drink from various fountains, feed, and defecate in private gardens and public parks. The Barbary macaque is a species on the way to disappearance. 27 Although they can facilitate ecotourism, their contact with human and pets can negatively influence the Barbary macaque. 28
In this work, we aimed to investigate the occurrence of carbapenemase-producing Enterobacteriaceae (CPE) isolates from wild boars and Barbary macaques in Bejaïa (Algeria) and to evaluate the potential of virulence of these strains. We also analyzed the phylogenetic grouping and determined the clonal relatedness of these isolates by multilocus sequence typing (MLST). Interestingly, in this study, we report the first detection of carbapenemase-producing bacteria isolated from wild boars in Algeria. This document represents one of the few reports of OXA-48-producing Enterobacteriaceae in wildlife.
Materials and Methods
Sample collection and processing
Fecal samples were collected from wild boar and Barbary macaque from September 2014 to April 2016. Thirteen individual rectal swabs were collected from wild boars that were shot near the Kabylie forests located in the north of Algeria (36°45′N 5°04′E) (Fig. 1). During this period, hunts were organized by local hunting associations, and samples from each hunt were independently transported to the laboratory. In addition, samples of fresh stools found along forest roads and foot paths were also collected using sterile swabs (wild boars n = 155, Barbary macaques n = 212) (Fig. 1). All samples were transported under refrigeration (4°C) to the laboratory and analyzed within a maximum of 4 hr. The swabs were cultured overnight at 37°C in 10 ml of nutrient broth (Fluka, St Louis). For CPE screening, 200 μl of the culture was streaked onto MacConkey agar plates (Fluka) containing ertapenem (0.5 mg/L) and was then incubated overnight at 37°C. Up to three colonies (per fecal sample) with a presumptive Enterobacteriaceae morphology growing on the MacConkey agar plates were selected and cultured individually on Trypticase soy agar (Fluka). The isolates were identified by matrix-assisted laser desorption and ionization time-of-flight mass spectrometry (Microflex, Bruker Daltonics, Bremen, Germany). 29
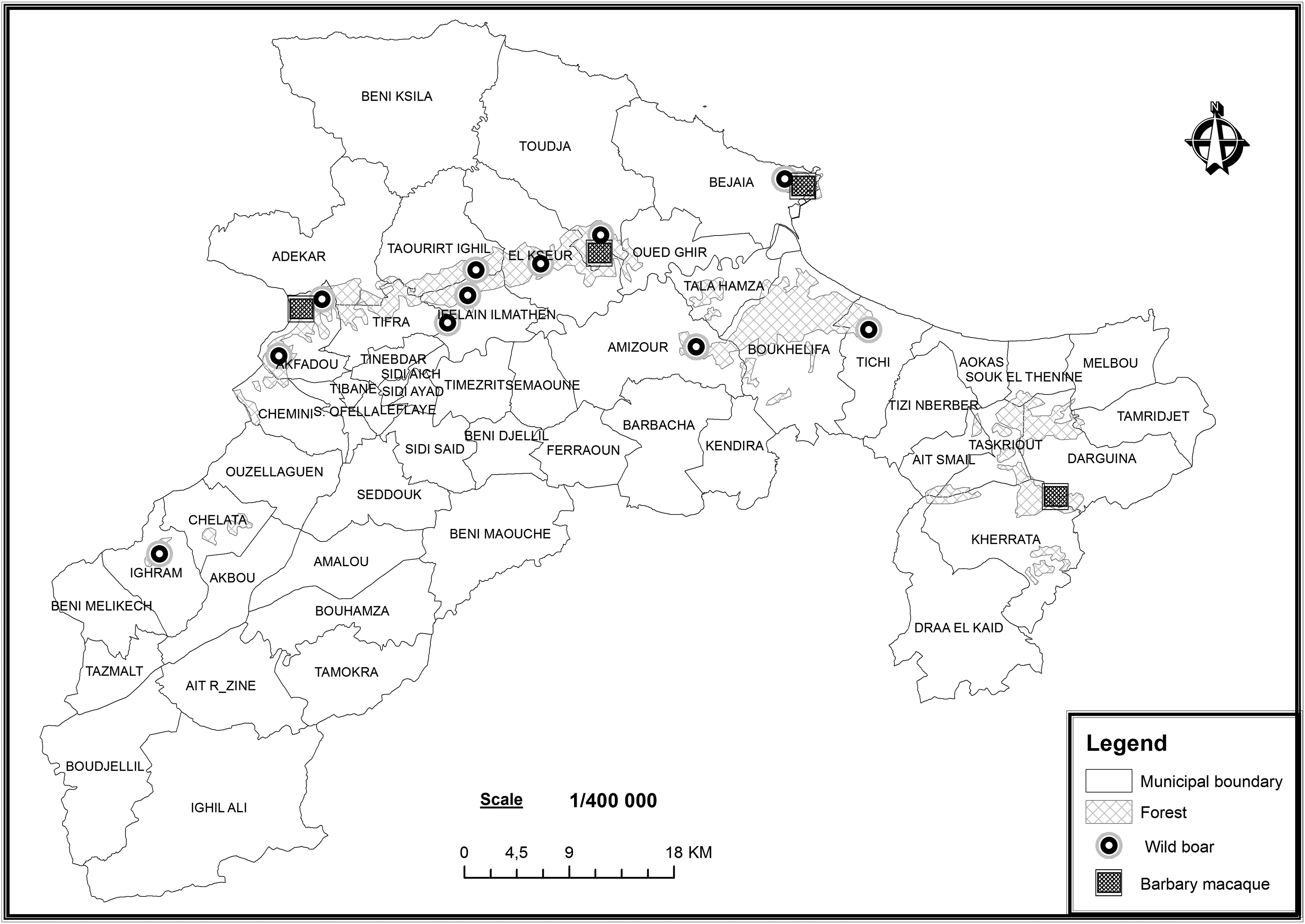
Map showing the locations of fecal samples collected from Barbary macaque and wild boar in different study sites in Bejaia, Algeria.
Antimicrobial susceptibility testing
Antibiotic susceptibility was tested on Mueller Hinton agar (Fluka) using the standard disk diffusion procedure as described by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (www.eucast.org). 30 The following antibiotics were tested: amoxicillin (AMX, 20 μg), amoxicillin–clavulanate (AMC, 20–10 μg), cefepime (FEP, 30 μg), cefotaxime (CTX, 5 μg), ceftazidime (CAZ, 10 μg), aztreonam (ATM, 30 μg), imipenem (IMP, 10 μg), ertapenem (ETP, 10 μg), meropenem (MEM, 10 μg), trimethoprim–sulfamethoxazole (SXT, 25 μg), amikacin (AK, 30 μg), kanamycin (KAN, 30 μg), tobramycin (TOB, 10 μg), gentamicin (CN, 15 μg), nalidixic acid (NA, 30 μg), pefloxacin (PEF, 5 μg), ciprofloxacin (CIP, 5 μg), ofloxacin (OFX, 5 μg), tigecycline (TGC, 15 μg), and colistin (CT, 50 μg) (Bio-Rad, Hercules, CA). Interpretation was based on the EUCAST breakpoint. The reference strain E. coli ATCC®25922 was included as control.
Phenotypic carbapenemase detection
The modified Carba NP test was used to confirm carbapenemase production, as described by Bakour et al. 31 The microbiological testing of MBL activity was determined using ethylenediaminetetraacetic acid inhibition as previously described. 32
Molecular analysis of beta-lactamase determinants
Standard PCR was performed to screen for the presence of blaKPC, blaOXA-48, and blaNDM carbapenemase genes, 33 as well as ESBL genes (blaTEM, blaSHV, and blaCTX-M 33 ) and pAmpC genes (blaCMY-1, blaCMY-2, and blaDHA-1). 34 Negative and positive controls were used in all PCR reactions.
Phylogenetic grouping and screening for virulence determinants
E. coli isolates were assigned to the phylogenetic groups (A, B1, B2, C, D, E, and F) by using the new quadruplex PCR based on the method described by Clermont et al. 35
E. coli strain isolates were also examined for the presence of genes encoding certain putative virulence determinants (Table 1), including papC (C adhesin classes of P fimbriae), sfa/foc (S and F1C fimbriae), afa/dra (binding, adhesions), iutA (siderophores included aerobactin), kpsM II (group II capsule synthesis), fyuA (genes of yersiniabactin), ibeA (invasion of brain endothelium), hlyA (alpha-hemolysin A), iucD (aerobactin siderophore synthesis), and papG I,II, III alleles (G adhesin classes of P fimbriae). The amplification was carried out with the following thermal cycling conditions: 95°C for 12 min. This was followed by 25 cycles of denaturation (94°C, 30 sec), annealing (63°C, 30 sec), and extension (68°C, 3 min), and a final extension (72°C, 10 min). 36
For K. pneumoniae strain, PCR was used to detect genes encoding the adhesins (fimH-1, mrkD), FimH-like adhesin (kpn), aerobactin receptor (iutA), yersiniabactin biosynthesis (irp-1), yersiniabactin receptor (fyuA), catecholate siderophores receptor (iroN), enterobactin biosynthesis (entB), mucoviscosity-associated gene A (magA), regulator of mucoid phenotype A (rmpA), and cytotoxic necrotizing factor-1 (cnf-1). (Table 1). PCR conditions were: 94°C for 4 min, followed by 30 cycles of 94°C for 30 sec, appropriate annealing temperature for 40 sec, 72°C for 1 min, and 72°C for 10 min. 37
Molecular epidemiology
MLST was performed using seven conserved housekeeping genes (pgiB, gap, tonB, mdh, phoE, infb, and rpoB) for K. pneumoniae, according to Pasteur schemes available on the Institute Pasteur MLST website (www.pasteur.fr/mlst) and (purA, icd, adk, fumC, recA, mdh, and gyrB) for E. coli, as described at the E. coli MLST database (http://mlst.warwick.ac.uk/mlst/dbs/E. coli).
DNA sequencing
PCR products were purified and sequenced using Big Dye terminator chemistry on an ABI 3730 automated sequencer (Applied Biosystems, Foster City, CA). The sequences obtained were analyzed using BlastN and BlastP against the NCBI database (www.ncbi.nlm.nih.gov).
Conjugation and transformation assays
Conjugation experiments were performed by the broth mating assay using a sodium azide-resistant J53 E. coli strain as recipient. 38 Transconjugants were selected on Luria Broth containing 200 mg/L sodium azide and 0.5 mg/L of ertapenem. The assay has been redone three times. When no transconjugants were obtained, transformation assays were performed. Plasmids were extracted by using the High Purity Plasmid Miniprep Kit (Neobiotech, Los Angeles, CA) according to the manufacturer's protocol. Then, transformation assays of extracted plasmids into E. coli DH10B ElectroMAX competent cells (Invitrogen, Carlsbad, CA) were carried out by electroporation as previously described. 39 Transformants were selected on LB agar plates containing 0.5 mg/L of ertapenem. 39
In addition, PCR amplifications targeting blaOXA-48 gene were also performed on the extracted plasmids.
Results
Of the 380 fecal samples collected from wild boars (n = 168) and Barbary macaques (n = 212), three carbapenem-resistant isolates suspected to produce carbapenemases were obtained from individual wild boars. The modified Hodge test and carba NP test were positive for these strains. The isolates were identified as E. coli (two isolates) and K. pneumoniae (one isolate) using MALDI-TOF MS. No carbapenem-resistant isolates were obtained from Barbary macaques.
The antibiotic susceptibility profiles of the three isolates showed that these isolates are resistant or intermediate to amoxicillin, amoxicillin–clavulanate, tobramycin, ertapenem, and meropenem. They remained susceptible to most of the antibiotics tested, including, cefepime, cefotaxime, ceftazidime, aztreonam, imipenem, trimethoprim-sulfamethoxazole, amikacin, kanamycin, gentamicin, nalidixic acid, pefloxacin, ciprofloxacin, ofloxacin, tigecycline, and colistin (Table 2).
AK, amikacin; AMC, amoxicillin–clavulanate; ATM, aztreonam; AX, amoxicillin; CAZ, ceftazidime; CIP, ciprofloxacin; CN, gentamicin; CT, colistin; CTX, cefotaxime; ETP, ertapenem; FEP, cefepime; IMP, imipenem; KAN, kanamycin; MEM, meropenem; NA, nalidixic acid; OFX, ofloxacin; PEF, pefloxacin; SXT, trimethoprim–sulfamethoxazole; TGC, tigecycline; TOB, tobramycin.
Molecular characterization by PCR and DNA sequencing showed that all carbapenem-resistant isolates harbored the blaOXA-48 gene (Table 3). None of the isolates was found to carry the blaNDM, blaKPC, blaCTX-M, blaTEM, blaSHV, blaCMY-1, blaCMY-2, and blaDHA genes.
MLST, multilocus sequence typing; NT, not tested.
Analysis of E. coli phylogroups showed that these isolates belong to phylogroup A (B043 and B044) and theses isolates were negative for all virulence genes tested. The K. pneumoniae isolate (B063) was positive for multiple virulence factors, including: fimH-1, mrkD, entB, irp-1, and fyuA (Table 3).
The clonal relationship of isolates analyzed by MLST showed that the two OXA-48-producing E. coli isolates were assigned to a unique sequence type, ST635. These strains were isolated from two rectal swabs from two different wild boars hunted in the same day. The OXA-48-producing K. pneumoniae strain recovered from the fresh wild boar stools belonged to ST13 (Table 3).
No transconjugants or transformants were obtained for the three strains tested. However, the PCR amplifications performed on extracted plasmids of the two E. coli strains showed positive results for blaOXA-48 gene suggesting a plasmidic location of blaOXA-48. On the other side, PCR amplification targeting blaOXA-48-producing K. pneumoniae showed that the isolate harbored a unique plasmid that did not contain the blaOXA-48 gene, suggesting a chromosomal insertion of this gene.
Discussion
The global spread of acquired carbapenemase-producing microorganisms has become a serious problem for public health. 40 In addition to humans and pets, antimicrobial resistance has also been reported in wildlife that has not been exposed to antibiotics.24,41,42 In this respect, Fischer et al. reported for the first time the isolation of NDM-1-producing Salmonella from a black kite (Milvus migrans) in Germany. 9 Also, Dolejska et al. have isolated IMP-4-producing Enterobacteriaceae in the silver gull on Five Islands in Australia. 19 Furthermore, Bhardwaj et al. reported the isolation of NDM-5-producing E. coli strains from a tiger in India. 18
In our study, the three isolates recovered from three different wild boar samples, were found resistant to ertapenem, meropenem, and penicillins and remain susceptible to broad-spectrum cephalosporins. This phenotype is evocative of OXA-48 carbapenemase, 43 which was confirmed by PCR amplification. To the best of our knowledge, this is the first report of CPE strains in wild animals in Algeria and in Africa and the first report on wild boars in the world. To date, there is one report from Algeria concerning the presence of OXA-48-producing E. coli in animals. 17 However, a limited number of studies on OXA-48-producing strains in Algeria has been published, involving clinical strains recovered from hospital or community settings,7,44,45 from fresh vegetables. 46 Concerning the environment, one report about the presence of OXA-48-like-producing Enterobacteriaceae isolated from river water in Algeria was published recently. 47
In our study, the blaOXA-48 genes identified among E. coli strains isolated from two captured wild boars living in the El Kseur forest (Bejaia) are located on plasmids. The blaOXA-48 gene is mostly linked to the dissemination of a single IncL/M-type self-transferable plasmid, 8 but chromosomal insertions were also reported, 48 as probably observed for OXA-48 producing K. pneumoniae strain isolated in our study.
Our MLST results showed that the two E. coli strains belong to ST635, and K. pneumoniae was assigned to ST13. The two sequence type (STs) identified in our study were different to those reported in Algeria. In these studies, the authors have reported that blaOXA-48 genes producing Enterobacteriaceae were associated with different STs identified in several Algerian hospitals,7,45,49–51 in fresh vegetables, 46 and in the Soummam River. 47
Nevertheless, the two STs found in our study were already identified in the world. The ST13 sequence type of K. pneumoniae is an internationally disseminated clone in clinical isolates, which in Ireland and Finland has been found to carry OXA-48.52,53 Concerning ST635, it was identified in E. coli isolates, which carried the blaNDM-12 gene recovered from a urine sample in Nepal 54 and the blaSHV-12 gene isolated from hospitalized patients in The Netherlands. 55
These findings indicate that there is a potential transmission of multidrug-resistant pathogens from humans to wild boars and vice versa. First, these groups of wild boars are endemic animals living in African nature reserves, where warm and humid climate in the continent makes favorable conditions for the persistence of bacterial contamination in the environment. 56 In addition, the anthropogenic impact on the environment, for example, generating waste water from human settlements, industrial and agricultural discharges containing resistant bacteria can influence the epidemiology and spread of these bacteria and their transmission either directly or indirectly to wild boar. The obvious explanation for this transmission is their feeding habits, involving garbage and waste water contaminated not only by bacteria of human origin but also by rodent feces, domestic animals, or animal manure. 24 Another risk factor for the spread of carbapenemase-producing bacteria throughout the wild environment can be the wild birds; they can easily transmit theses strains to waterways and other environment sources, including wildlife, through their fecal deposits.9,19,20,57,58 As previously described; wild birds are considered to be a reservoir of carbapenemase-encoding genes. 9
On the other hand, the acquisition of multidrug-resistant zoonotic pathogens might be by the colonization of wild boars with resistant strains harbored by their prey, 59 and since the wild boars can live in close contact with humans 24 they can become a vector of carbapenemase-encoding genes in the environment, which could increase their transmission to domestic animals and thereby its public health impact.
Studies realized on antimicrobial-resistant bacteria in wild animals can extend our knowledge on hosts’ specific resistance patterns, and thus to intervene more easily in case of an epidemic. 56
The commensal E. coli strain was found generally in the gastrointestinal tracts of humans and animals. 60 The pathogenic potential of E. coli is determined by several virulence factors. 36 In this study, no virulence genes were identified for the two E. coli isolates. According to clinical data, pathogenic K. pneumoniae strains have caused many infectious diseases. 61 Our findings demonstrated that K. pneumoniae strain isolated from wild boar carry a set of virulence genes associated with biofilm formation, host colonization, and mucous adherence as previously reported.37,61,62 This situation shows that these virulence factors are important for Klebsiella pathogenicity.
One limitation of our study is the limited size of the sample considered and the reduced number of strains. Therefore, the use of a serial dilution method is very useful to enumerate CPE population in a sample. Another major problem is the absence of routine methods implanted in Algerian laboratories, such as the availability of chromogenic media.
In conclusion, our findings strongly suggest that wild boars may be reservoirs of strains, which carry carbapenemase-encoding genes and virulence genes. However, further research should be done to gain a more detailed understanding of the importance of the impact of microbial contamination in the environment.
Footnotes
Acknowledgments
The authors thank Linda Hadjadj for technical assistance and all those who participated in carrying out this work. They thank TradOnline for English corrections. All authors have read and approved the article.
Disclosure Statement
No competing financial interests exist.