
Abstract
Transmissible plasmids and integrons may play important roles in the persistence and spread of antibiotic-resistant bacteria throughout aquatic environment by accumulating antibiotic resistance genes (ARG). Class 1 and class 2 integron (intI), mobilization (mob), sulfamethoxazole resistance (sul), and trimethoprim resistance (dfr) genes were PCR-amplified and confirmed through DNA sequencing following plasmid extraction from 139 antibiotic-resistant Escherichia coli. E. coli had previously been recovered from wastewater treatment plant effluent and receiving stream water in Northwest Arkansas and isolates had expressed resistance to one to six antibiotics. Almost half of the total isolates (47%) carried putatively transmissible plasmids with mobF12 gene as the most frequently detected mobilization gene. When two or three mob genes were detected per isolate, there was a significant shift in the population toward larger multiple drug resistance (MDR) number. Class 1 and/or 2 integrons were prevalent (46%), and the presence of integron significantly shifted the isolate population toward larger MDR number. More isolates carried single or coexistence of two or three sul genes (99.3%), and single or a combination up to five dfr genes (89.3%) than had exhibited in vitro resistance to the respective antibiotics. These findings indicate not only the role of the wastewater treatment effluent and the stream environment in coaccumulation of ARG with transmissible plasmids and integrons in multiple antibiotic-resistant E. coli populations but also suggest that density of sul and dfr resistance genes within an isolate may serve as a biomarker for mobile MDR in general.
Introduction
T
In aquatic environments, wastewater treatment plants (WWTPs) can serve as input sources to streams of ARGs, bacteria, and antibiotics, including sulfamethoxazole and trimethoprim. 4 The WWTP in Fayetteville, Arkansas received inflows of an average of 42.8 million liters per day originating from various anthropogenic sources, such as hospitals, residences, and industries; and uses UV treatment as tertiary treatment before releasing treated effluent. 5 Yet, Escherichia coli and broad-host-range plasmids, 6 and ARG 7 remained in discharged WWTP effluent, which led to inputs of corresponding ARB in water and sediment of the receiving stream Mud Creek. 5 A number of E. coli exhibiting in vitro resistance from one to six different antibiotics—specifically ampicillin, gentamicin, ofloxacin, tetracycline, sulfamethoxazole, and trimethoprim—and plasmids were recovered from one site upstream, WWTP effluent discharge, and two sites downstream of the pipe discharging treated effluent from the Fayetteville, Arkansas WWTP into Mud Creek.5,6 Genotypically, these multiple drug resistance (MDR) determinants might be associated with mobile genetic elements of transmissible plasmids and integrons carrying sulfamethoxazole (sul) and trimethoprim (dfr) resistance genes.
The presence of ARG in bacterial cells is strongly associated with mobile genetic elements, such as plasmids. 8 In terms of conjugative apparatus and mobility, plasmids are classified into transmissible plasmids (i.e., conjugative or self-transmissible plasmids and mobilizable plasmids) and nonmobilizable plasmids. 9 For transmissible plasmids, the first possesses a complete set of conjugation genes comprising mob genes required for the processing of conjugative DNA and mating pair formation genes required for forming the mating channel; whereas, the latter harbors only mob genes allowing the plasmid to be mobilized by conjugation when it coexists in the same donor cell with a conjugative plasmid.9,10
The ARG dissemination of conjugative and mobilizable plasmids might increase when ARG are present within other genetic elements, such as an integron. Integrons have been highly associated with the occurrence of many different ARG, including the dfr (dihydrofolate reductase) and sul (dihydropteroate synthetase) genes that are related to trimethoprim and sulfamethoxazole resistance, respectively.11,12 The accumulation of multiple sul and/or dfr genes, possibly within integrons of conjugative and/or mobilizable plasmids, may be an important contributor to the persistence and spread of ARB throughout the environment.13,14 Within the integron structure, sulfamethoxazole resistance (sul1) genes are commonly encoded in 3′-conserved segments, whereas trimethoprim resistance (dfr) genes are frequently found on the variable regions (gene cassette array). 15 Although some integrons lack dfr gene cassettes or the typical 3′-conserved segments, the occurrence of integrons might still be useful as an indicator of the occurrence of sul and dfr genes.16,17
The present study focused on detection of two antibiotic resistance (sul and dfr) genes and their associations with the presence of transmissible plasmids (mob genes) and/or integrons (intI genes), since those two ARGs are highly related to the integron structure and aligned with our other studies of persistence 14 and dissemination 13 that specifically used sulfamethoxazole and trimethoprim-resistant E. coli isolates. Because of our other related studies, we were particularly interested in determining if sulfamethoxazole and trimethoprim resistance genes, including redundancy among sul and dfr genes, were related to increased MDR number in isolates from our location (i.e., Arkansas in the mid-southern United States) where this research had not been previously conducted. Other resistance genes (ampC, oxa2, and tetA) had been investigated in a separate study. 7 Hence, the objectives of the research were to determine if the presence of transmissible plasmids, integrons, and/or sul and dfr genes was significantly associated with resistance to greater number of antibiotic classes, or MDR number, of antibiotic-resistant E. coli isolated from treated wastewater effluent and receiving stream water.
Materials and Methods
Initial sources were 139 E. coli harboring plasmids and ARG combined from among four collection sites (WWTP effluent, 20 m upstream, and 640 m and 2,000 m downstream from WWTP effluent discharge) in an effluent-driven stream Mud Creek, a tributary of the Illinois River, located in Fayetteville, Arkansas (36.090° latitude, 94.111° longitude). Mud Creek tributary has a 293 ha of catchment area with 57.7%, 42.3%, and less than 1% of its land use for forests, urban, and pasture, correspondingly. 18 Initial water collection and data and bacterial isolation and characterization procedures are provided in a previous investigation. 5 Isolates were reported to be in vitro-resistant to one of the following antibiotics: 32 μg/ml ampicillin (Am), 16 μg/ml tetracycline (Tc), 8 μg/ml ofloxacin (Of), 4 μg/ml trimethoprim (Tm), or 80 μg/ml sulfamethoxazole (Sx). 5 After initial isolation and in vitro resistance determination, isolates were further screened for resistance against five more antibiotics using E-test® strips (AB BIODISK, Solna, Sweden): Am, 0.016–256 μg/ml; Of, 0.002–32 μg/ml; Tc, 0.016–256 μg/ml; Tm, 0.002–32 μg/ml; Sx, 0.064–1,024 μg/ml; and gentamicin (Gm), 0.016–256 μg/ml with minimum inhibitory concentration of 16 μg/ml, following the protocol and minimum inhibitory concentrations supplied by the manufacturer. 5 Isolates were stored in glycerol solution at −76°C. Of 139 isolates, a total of 17 isolates are resistant to 5 or 6 antibiotics, whereas 30, 62, 11, and 19 isolates are identified to be resistant to 4, 3, 2, and 1 antibiotic, respectively (Table 1).
Ampicillin (Am), gentamicin (Gm), ofloxacin (Ox), tetracycline (Tc), trimethoprim (Tm), and sulfamethoxazole (Sx).
intI gene combinations followed by the same letter are not significantly different using a contrasts procedure (p ≤ 0.05).
The E. coli strains were grown on selective Luria-Bertani (LB) agar plates containing antibiotic(s), to which the isolates were originally determined to be resistant, and incubated at 37°C overnight. Single colonies were inoculated into 10 ml of selective LB broth (Mobio, Carlsbad, CA) containing antibiotic(s) similar to LB agar plates, which were incubated overnight at 37°C with shaking at 150 rpm. Plasmid extraction was performed using the Wizard® Plus SV Minipreps DNA Purification System (Promega, Madison, WI) according to the manufacturer's instructions.
The extracted plasmids of DNA were used as template to determine the occurrences of mobilization genes (mobP11, mobP14, mobP51, mobF11, mobF12, mobQ11, and mobQu); integrons (intI1 and intI2); sulfamethoxazole (sul1, sul2, and sul3) and trimethoprim (dfrA1, dfrA8, dfrA12, dfrA14, dfrA17, and dfrB3) resistance genes using PCR amplification with primer sets outlined in Table 2. Trimethoprim genes were selected based upon their prevalence on E. coli recovered from the clinical isolates of human and animal origin associated with integrons. 17 All PCR amplifications were performed in 20 μL reactions containing 1 × PCR buffer, 2.5 mM MgCl2, 200 μM dNTPs, 400 ng/μL bovine serum albumin (Merck KGaA, Darmstadt, Germany), 0.5 μM of each primer, 1 μL of template DNA, and 0.5 U of GoTaq DNA polymerase (Promega). Diethylpyrocarbonate (DEPC)-treated water (EMD Millipore, Darmstadt, Germany) was used as no template control run in parallel with samples. The PCRs were carried out using a PTC-200 thermocycler (MJ Research, Waltham, MA) under conditions as follows: initial denaturation at 94°C for 5 min, followed by 30 cycles of 94°C for 30 sec, annealing (varied temperature, see Table 2) for 30 sec, and 72°C for 60 sec, with a final extension at 72°C for 8 min. The PCR products were analyzed on 1.5% (w/v) agarose gels with ethidium bromide at 100 V for 50 min in 0.5 × TBE buffer and visualized using gel documentation to assess bands of the expected size. Additional confirmation of the PCR products was performed through DNA sequencing (Eurofin Genomics, Kansas City, Kansas).
See reference list for bibliographic information.
FW, forward; RV, reverse.
A statistical analysis was performed to evaluate the effects of occurrence of integron, mobilization genes, sulfamethoxazole resistance genes, and trimethoprim resistance genes toward the MDR number with 95% confidence intervals using GLIMMIX procedure on SAS 9.4 (Cary, NC). The data were analyzed based on a multinomial logit model with a cumulative logit link function and the results were back-transformed to the proportion scale for presentation of the results. Following the preliminary overall test for treatment effects, contrasts were used to compare individual pairs of treatments (p ≤ 0.05) on the cumulative logit scale.
Results
There was a statistically significant difference in mob gene distribution across MDR number (p = 0.0014, Fig. 1). A total of 65 (46%) isolates carried transmissible plasmids indicated by the presence of mob genes (Fig. 2a). Although the mobF12 gene was most prevalently detected from 54 (39%) of 139 total isolates or 83% of the total transmissible plasmids, the mobF12 gene was not only detected singly (26% isolates) but also in coexistence with another mob gene (9% isolates) and with two other mob genes (4% isolates) across all MDR numbers. The co-occurrence of two or three mob genes, which often was mobF12 in combination with either mobP51 or mobQu (Fig. 2a) resulted in a higher proportion of increased MDR in the resistant E. coli population (Fig. 1).
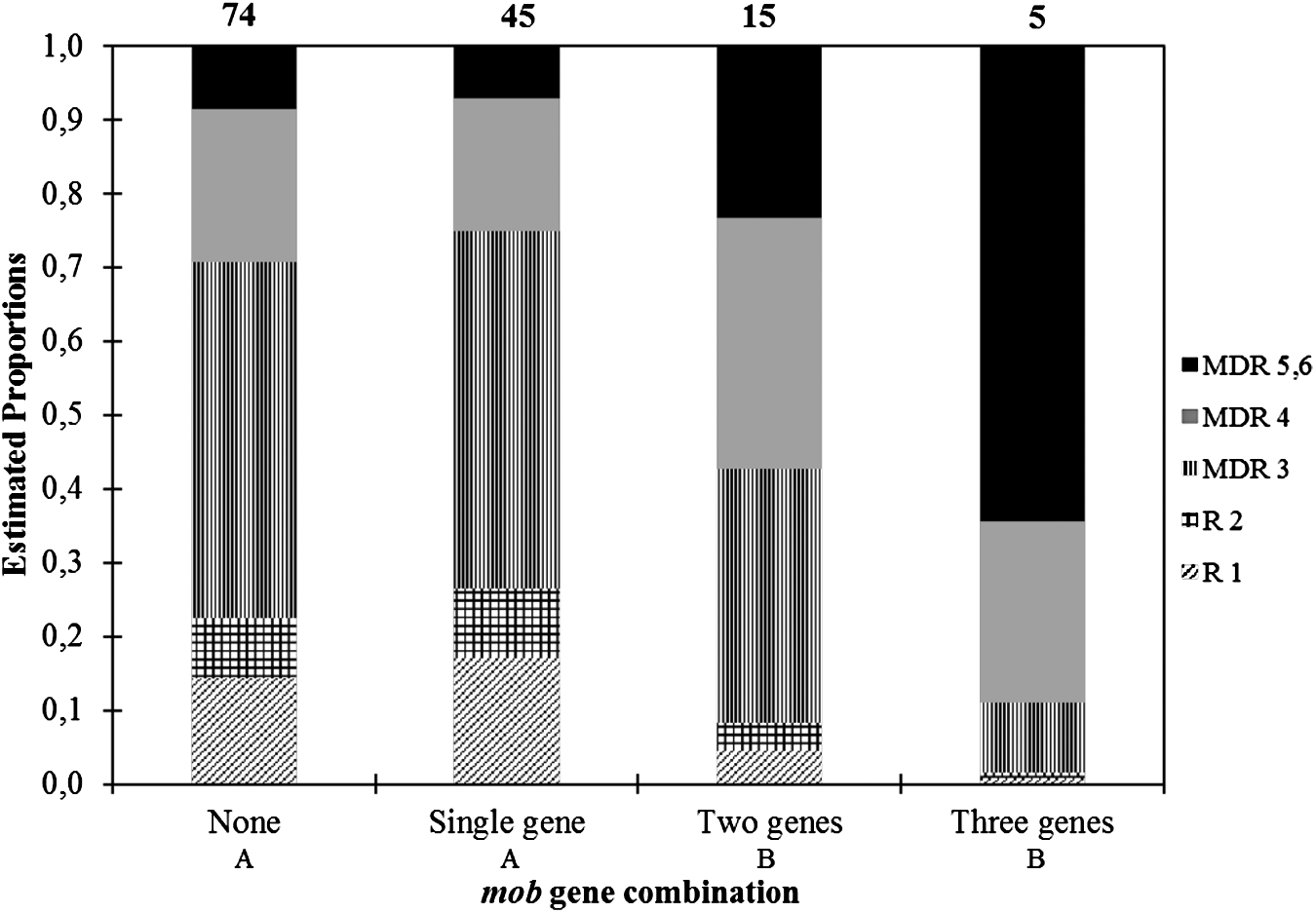
Estimated proportion (estimated using contrasts tested on the cumulative logit scale) of mob gene combination (e.g., number of mob genes) among 139 MDR Escherichia coli isolated from Mud Creek in Fayetteville, Arkansas (R1 n = 19, R2 n = 11, MDR 3 n = 62, MDR 4 n = 30, MDR 5, 6 n = 17). Numbers above each column are the total number of detections for each gene combination. Gene combinations followed by the same capital letter are not significantly different (p ≤ 0.05). MDR, multiple drug resistant.
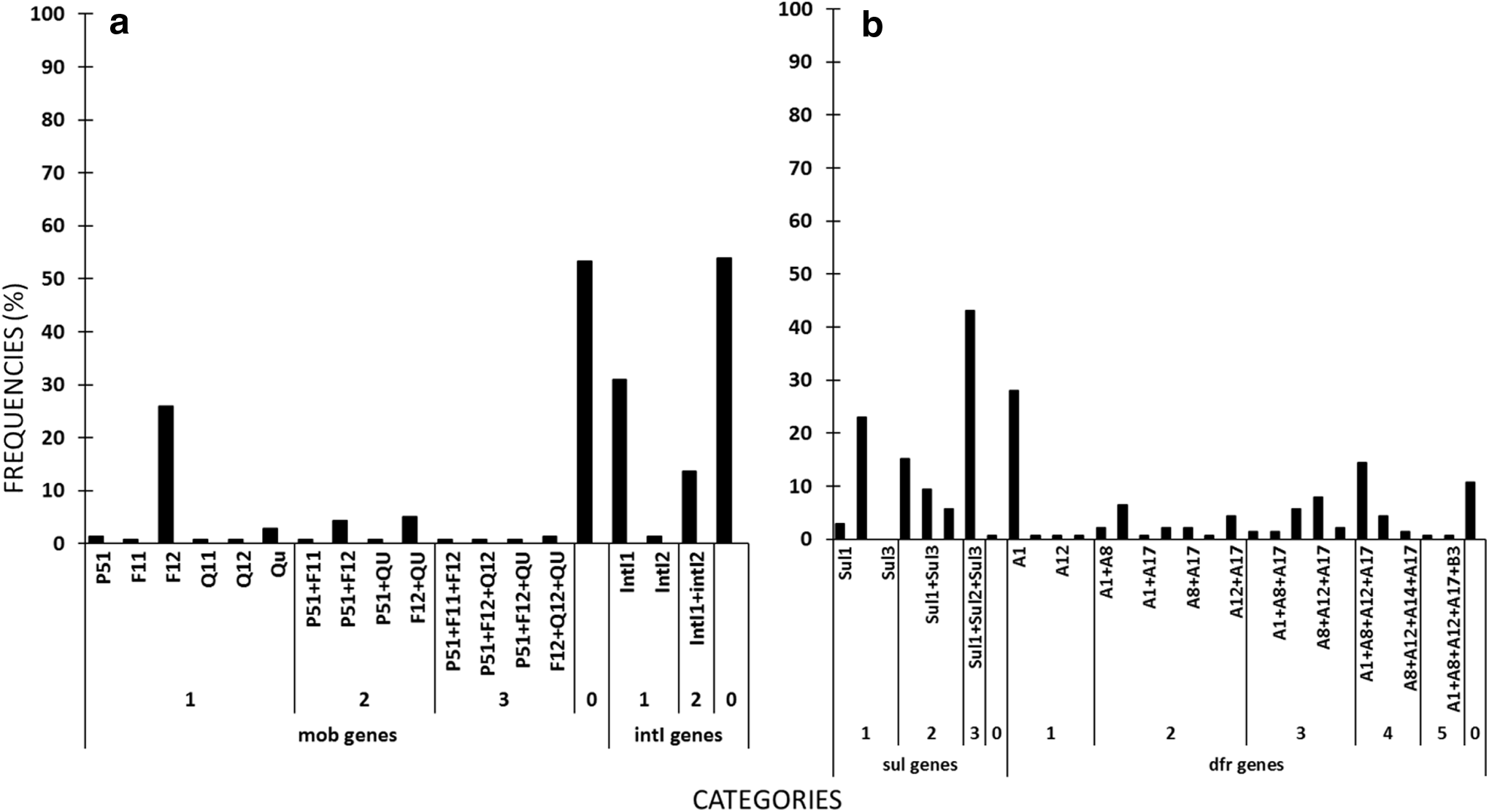
Frequency of occurrence (%) of
Similar to the mob genes, there was also a significant difference (p < 0.0001) in distribution of intI genes among isolates across MDR number (Fig. 3). A total of 64 (46%) of 139 isolates were detected to harbor integron genes, which were subdivided into class 1 integron (31% isolates); class 2 integron (2% isolates); and both class 1 and 2 integron (14% isolates) (Fig. 2a). Isolates displaying in vitro resistance to greater number of antibiotic classes (MDR 4 to MDR 5 or 6) were positively associated with integrons, particularly class 1 integrons (Fig. 3).
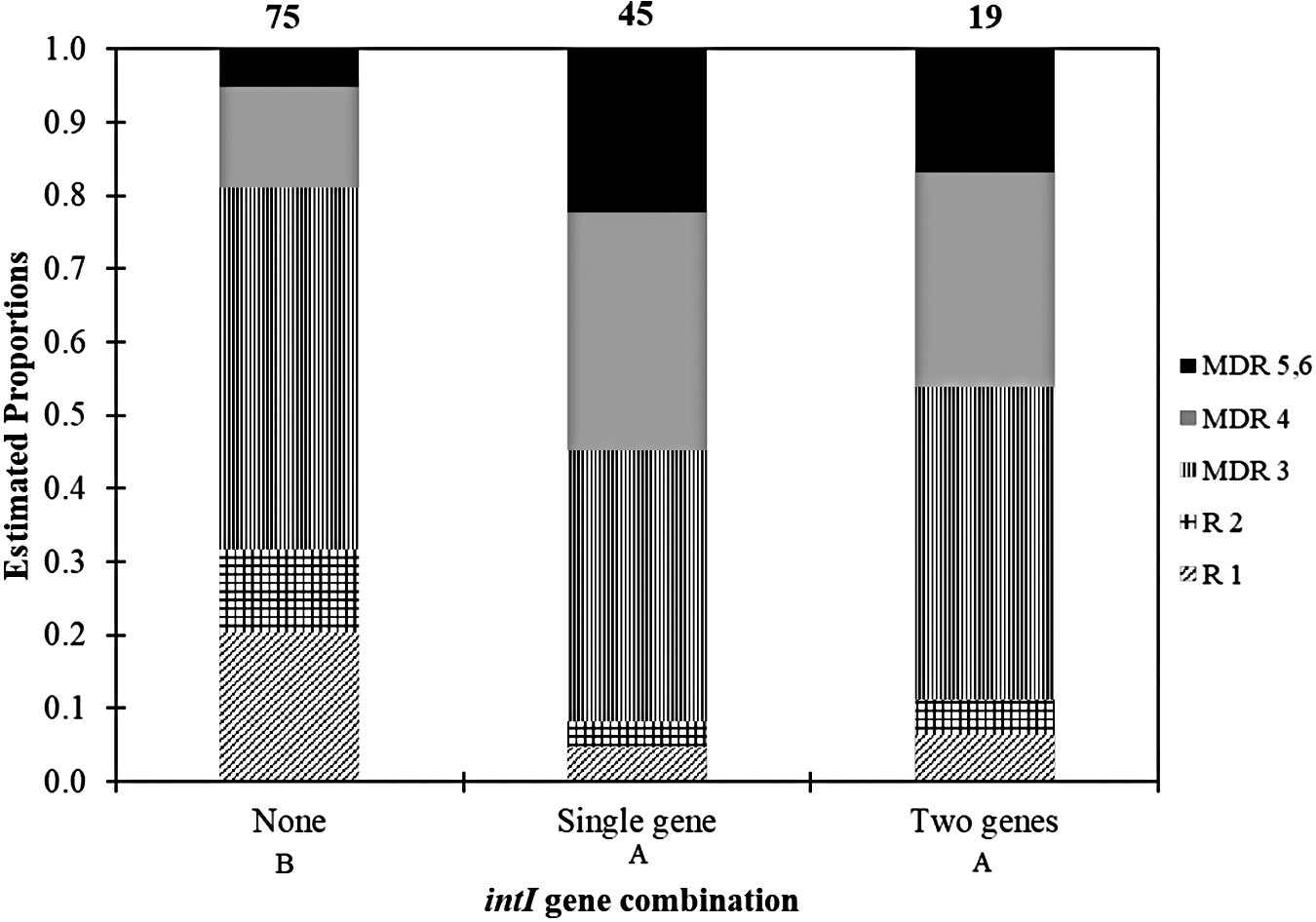
Estimated proportion (estimated using contrasts tested on the cumulative logit scale) of intI gene combination (e.g., number of intI genes: none, intI1 or intI2, or intI1+intI2) among 139 MDR E. coli isolated from Mud Creek in Fayetteville, Arkansas (R1 n = 19, R2 n = 11, MDR 3 n = 62, MDR 4 n = 30, MDR 5, 6 n = 17). Numbers above each column are the total number of detections for each gene combination. Gene combinations followed by the same capital letter are not significantly different (p ≤ 0.05).
A total of 99% of the isolates were positive for at least one sul gene, whereas 105 (75%) isolates had displayed in vitro resistance to sulfamethoxazole leading to a discrepancy between phenotype and genotype, that is, isolates did not show in vitro resistance but harbored sul genes for 34 (24%) isolates (Fig. 2b). A total of 74% isolates were redundant in encoding sulfamethoxazole resistance with coexistence of sul1, sul2, and/or sul3 genes from a single isolate. Coexistence of all three sul genes (sul1, sul2, and sul3) from a single isolate accounted for 60 (43%) of the total 139 isolates. This coexistence of all three sul genes was more likely to occur in E. coli resistant to four, five, or six antibiotics than if there were only one or two sul genes (Fig. 4). The occurrence of single sul gene and coexistence of two sul genes, on the contrary, were distributed similarly across MDR number compared to each other and were less likely to be detected on the isolates resistant to five or six antibiotics.
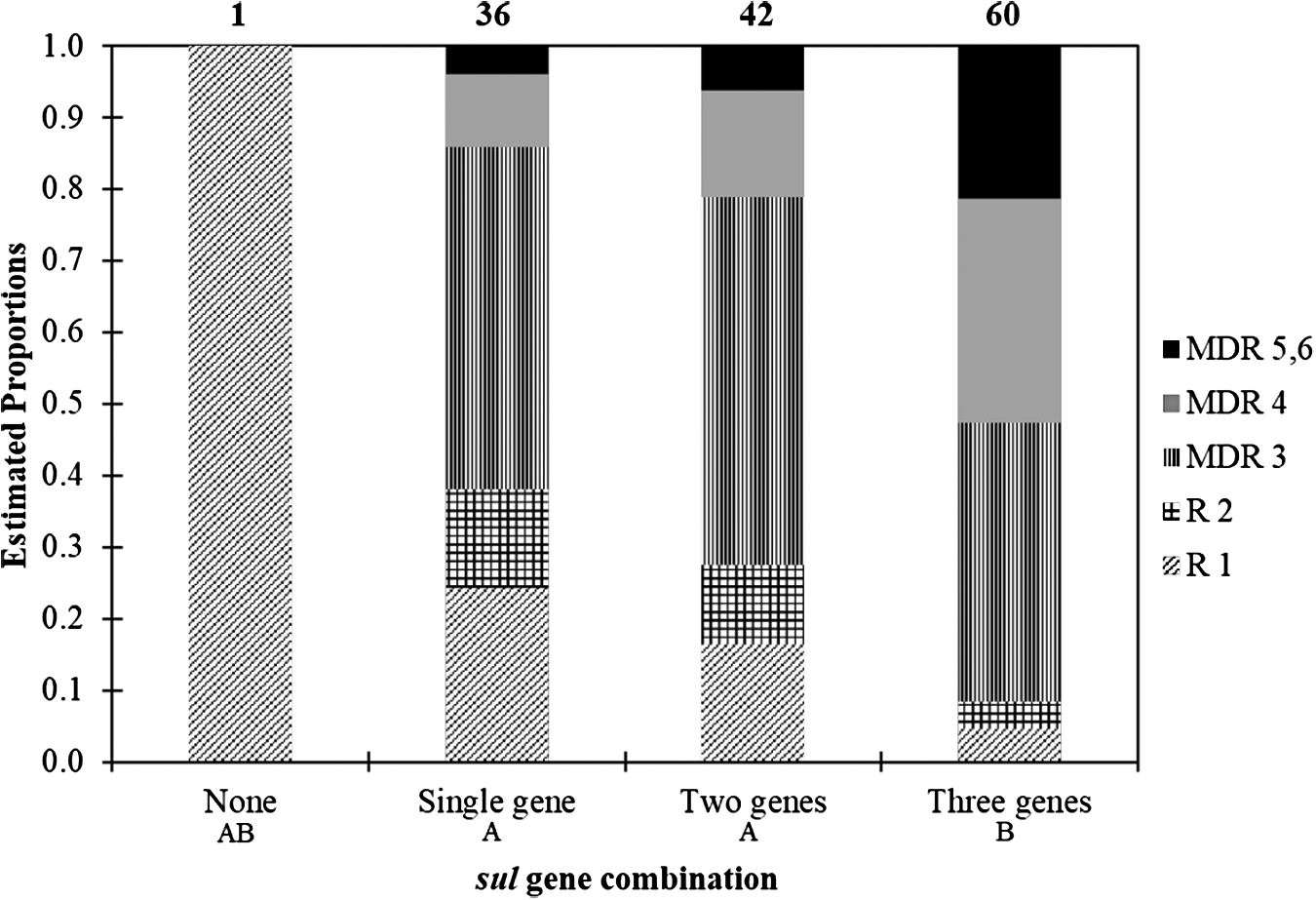
Estimated proportion (estimated using contrasts tested on the cumulative logit scale) of sul gene combination (e.g., number of sul genes) among 139 MDR E. coli isolated from Mud Creek in Fayetteville, Arkansas (R1 n = 19, R2 n = 11, MDR 3 n = 62, MDR 4 n = 30, MDR 5, 6 n = 17). Numbers above each column are the total number of detections for each gene combination. Gene combinations followed by the same capital letter are not significantly different (p ≤ 0.05).
Similar to sul genes, a higher proportion of isolates (89%) harbored one or more genes for resistance to trimethoprim than had exhibited in vitro resistance to the antibiotic (Fig. 5). A total of 59% isolates were redundant in encoding trimethoprim resistance with coexistence of two to five dfr genes from a single isolate. There were 84 (60%) isolates exhibiting in vitro resistance leading to a discrepancy of phenotype and genotype for 39 (28%) isolates. Comparable to the results for sulfamethoxazole, the proportion of isolates exhibiting in vitro resistance (60%) was similar to the proportion of the population exhibiting redundancy in resistance genes against trimethoprim. Three or four dfr genes were commonly detected from a single isolate accounting for 19% or 20% isolates, correspondingly, of the total E. coli. The dfrA1 gene was the most prevalent, followed by dfrA12, dfrA8, and dfrA17, whereas dfrB3 and dfrA14 genes were less often detected accounting for 10 (7%) and 5 (3.5%) of total isolates, respectively (Fig. 2b). While single dfrA1, dfrA8, dfrA12, and dfrA14 were detected in 30% of the total isolates, 17 combinations of two to five different dfr genes were detected in 59% of the total isolate population. Three to five coexisting dfr genes were carried within an isolate on a larger proportion of the population showing in vitro resistance to four, five, or six antibiotics compared to if the E. coli possessed one or no dfr genes (Fig. 5).
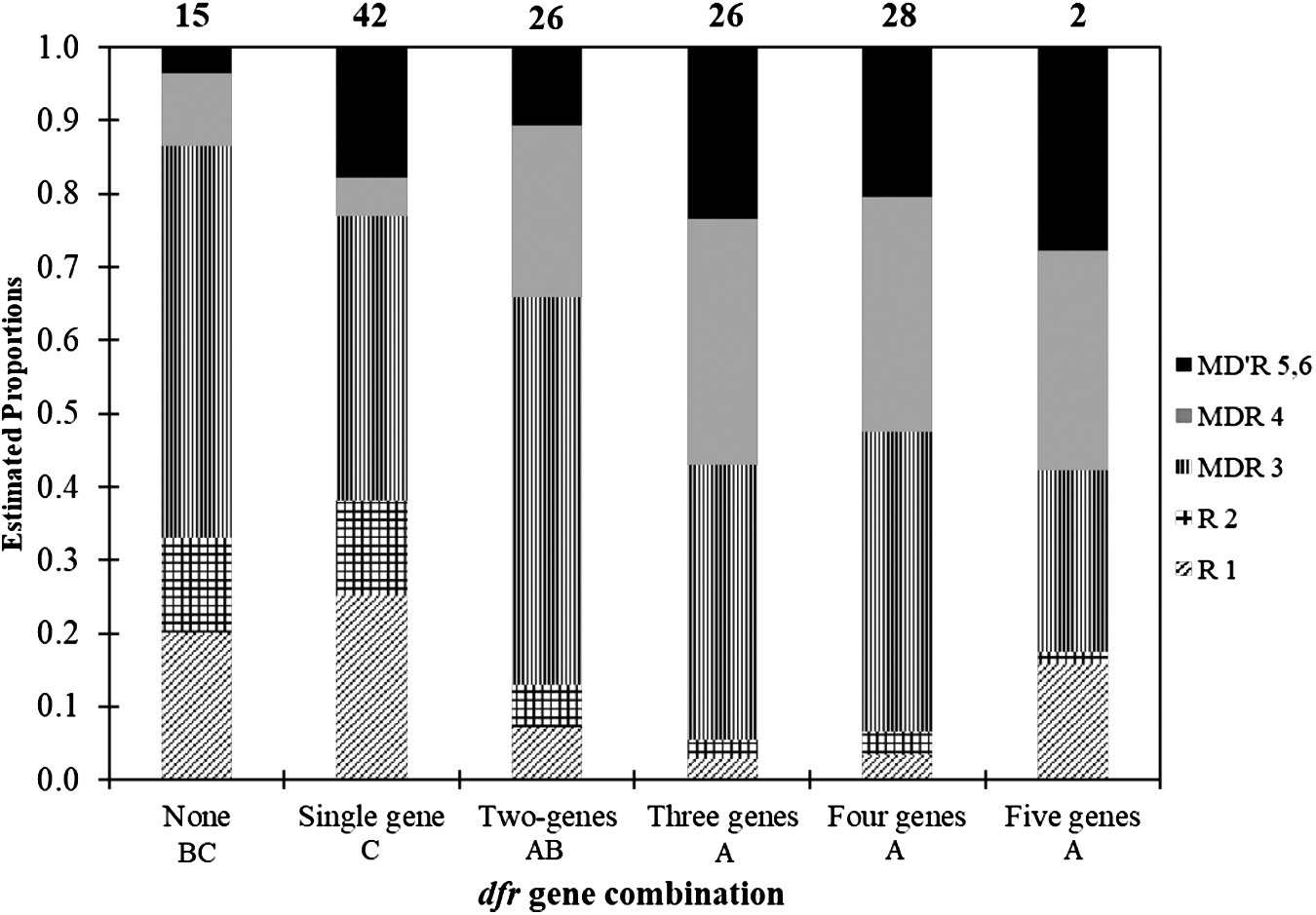
Estimated proportion (estimated using contrasts tested on the cumulative logit scale) of dfr gene combination (e.g., number of dfr genes) among 139 MDR E. coli isolated from Mud Creek in Fayetteville, Arkansas (R1 n = 19, R2 n = 11, MDR 3 n = 62, MDR 4 n = 30, MDR 5, 6 n = 17). Numbers above each column are the total number of detections for each gene combination. Gene combinations followed by the same capital letter are not significantly different (p ≤ 0.05).
Discussion
Prevalence of multiple drug-resistant bacteria, and the association of their ARG with genetic elements such as plasmids and integrons, is increasingly leading to significant implications in both clinical and nonclinical settings. In the present study, the presence of multiple transmissible plasmids or integrons shifted the distribution of ARB toward larger MDR numbers. ARG which are potentially disseminated throughout the environment within transmissible (conjugative or mobilizable) plasmids play a major role in the spread of ARG.13,19 Integrons have also been found to be prevalent in multiple antibiotic-resistant gram-negative and gram-positive bacteria. 20 Furthermore, the results of this study linked increased redundancy of sulfamethoxazole and trimethoprim resistance genes to isolates with increased MDR. 14 Overall, despite the potential limitations of the plasmid DNA extraction approach used, that is, that low copy number, large molecular weight plasmids may not have been extracted and that small amounts of chromosomal DNA could have contaminated our results, the results of this study confirm many of those obtained in other research.
Almost half of the total plasmids in this study were considered as transmissible plasmids indicated by the occurrence of mob genes. Similarly, ∼50% of plasmids recovered from γ-proteobacteria, including E. coli, were found to be potentially transmissible. 20 Conjugative plasmids were considered as a major means of the spread of ARG from multiple antibiotic-resistant E. coli coming from human fecal contamination. 19 Furthermore, the present study also demonstrated that more than half of the positively detected transmissible plasmids were identified as mobF12. These results are in agreement with the previous findings signifying mobF12 gene as the most frequently found mob gene among different genera of Enterobacteriaceae. 21 Moreover, mobF12 was found on more than 90% of plasmids isolated from clinical multiresistant E. coli. 22 In addition, mobF12 was the most abundant mob gene found on plasmid DNA of E. coli isolated from farm animal and human origins that were resistant to third-generation cephalosporin. 23 The mobF12 gene was detected alone or in coexistence with other mob genes in isolates harboring resistance from one to six antibiotics. It is possible that multiple plasmids could account for redundancy in resistance genes in at least some of the E. coli, and may contribute to the persistence and dissemination of antibiotic resistance among bacteria in surface water environments. 14
In addition to mob genes, almost half of the isolates possessed integrons. In terms of MDR number, the present study demonstrated the positive relationship between increasing MDR number and class 1 integron existing singly or in coexistence with class 2 integron. That the presence of class 1 with or without class 2 integrons resulted in a shift toward a greater proportion of the population with larger MDR numbers may indicate the role of integrons in facilitating multiple antibiotic resistance. 24 Despite the fact that class 1 integrons are not necessarily linked to antibiotic resistance, 25 other studies have indicated that bacteria harboring integrons are generally resistant to at least three different antibiotics.26,27 Integrons can potentially be used as a tool to predict the presence of multiple ARB in environmental samples 28 as well as to determine anthropogenic pollution since integrons commonly carry a wide range of resistance genes toward antibiotics, disinfectants, and heavy metals within their structure in a broad variety of pathogenic and nonpathogenic bacteria associated with human waste streams and agriculture. 29 Interestingly, there was no MDR pattern associated with the presence of integron.
The prevalence of class 1 integron (46% of the population) corroborates previous findings. Class 1 and class 2 integron occurred in 40% and 10%, respectively, of multiple antibiotic-resistant E. coli isolated from aquaculture water in Iran 30 and 32% and 3%, correspondingly, of the antibiotic-resistant E. coli isolated from two WWTPs and their receiving waters in Poland. 24 The discrepancy of occurrence between the more predominantly found class 1 integron than class 2 integrons in all of these studies might be related to the association of class 1 integrons with MDR determinants since integrons harbor greater diversity of ARG embedded within the gene cassette compared with the class 2 integrons. 26
Among sul genes, sul2 gene was detected more frequently than sul1 and sul3 genes in E. coli isolates in this study, supporting previous investigations demonstrating a gene-frequency distribution of sul2>sul1>sul3. 31 However, others found distributions of sul1>sul2>sul3.28,32 In the current study, sul3 genes were almost as frequently detected as sul1 genes; sul3 genes were commonly detected in coexistence with another sul gene (detected in 21 plasmids) or with two other sul genes (detected in 60 plasmids), suggesting the emergence of sul3 genes in the aquatic environment. Previously, sul3 genes were the least common gene or unexpressed 33 or were not detected in wastewater and shrimp ponds in North Vietnam. 34 The presence of the sul3 gene was earlier reported in E. coli isolated from animal 35 and human 36 origins.
Regarding the MDR number, the current study signified the redundancy of three sul genes in association toward increasing MDR number. The occurrence of sul1 genes are predominantly found in high levels in sulfonamide-resistant bacteria 37 and are highly associated with class 1 integron as sul1 gene is classically encoded within 3′ conserved regions of class 1 integron. 38 The presence of sul3 gene is emerging due to its association with integron that was recently identified as being encoded within the conserved segment (sul3-CS) of class 1 integron of plasmids recovered from commensal E. coli. 32
More than four-fifths (89%) of the isolates of the present study carried trimethoprim genes with a great diversity of different dfr genes detected in isolates, often within the same isolate, ranging from a single gene to five dfr genes. The four most predominant dfr genes in this study were dfrA1, followed by dfrA12, dfrA8, and dfrA17 genes, which is in accord with previous research demonstrating that dfrA1 was the most predominant dfr gene, followed by dfrA12, in commensal E. coli from pigs in Poland. 39 Both dfrA1 and dfrA12 genes were present in coexistence among two to four trimethoprim resistance genes. Others found that dfrA1 and dfrA17 were found most frequently in clinical isolates, especially in sulfamethoxazole–trimethoprim-resistant isolates from urine specimens.40,41 Similar to sul genes, the occurrence of redundant dfr genes was positively correlated with increasing MDR number. This signified the prevalence of dfr genes among multiple ARB, which might also be a result of association with integrons. Previously, integron-associated trimethoprim-resistant E. coli have been isolated from humans and animals in Lithuania 17 and in Syria. 42
Despite the prevalence and diversity of sul and dfr genes, a discrepancy between phenotype and genotype occurred in the present study accounting for 24% and 28%, respectively. Those isolates that do not show resistance in vitro, but harbor corresponding ARG (also known as silent/cryptic resistance) might act as a reservoir of ARG in antibiotic resistome. 43 These genes do not contribute to in vitro resistance in their original genetic state, yet, their potential determinants might be expressed by mutation, mobilization, or alteration of the expression. 43 In addition, the diversity and the abundance of the potential genes in the resistome might be accelerated along with horizontal gene transfer induced by the selective pressure of antibiotics in the environment. 6
The prevalence and diversity of both sul and dfr genes in the present study demonstrate a redundancy among plasmid-mediated sulfamethoxazole and trimethoprim-resistant genes. Isolates appear to be accumulating ARG, which could be a result of recombination events. 44 Moreover, both sulfamethoxazole and trimethoprim resistance genes are highly associated with other genetic elements, that is, integron and insertion sequences. 41 For example, sulfamethoxazole resistance is acquired due to presence of dihydropteroate synthase (DHPS) genes located on class 1 integrons for sul1 and sul3 genes 16 or genomic islands for sul2 genes. 45 As a part of adaptation to the changing selective pressure of antibiotics, including at subinhibitory level, in the environment, bacteria may be duplicating and amplifying their ARG, leading to increasing/reversing their plasmid size. 46 Subinhibitory concentrations of antibiotics are likely to be present in the stream water environment. A total of 5.8 μg/L of antibiotics have been detected in addition to various pharmaceutical and other organic chemicals in stream water, including Mud Creek in Northwest Arkansas, the stream where the isolates were originally sampled for this study. 47 Trimethoprim was detected at concentrations of 0.029–0.061 μg/L and sulfamethoxazole at concentrations of 0.356–0.564 μg/L in Mud Creek water. 48 A complex mixture of antibiotics and other pollutants plus ARB harboring genetic elements such as transmissible plasmids and integrons in the WWTP is an ideal hotspot and reservoir for ARG to be widely disseminated into aquatic environments. 49 Therefore, further study is needed to determine the effect of antibiotics, particularly when present at subinhibitory concentrations, toward the persistence of bacteria harboring different intI-mob combinations.
Conclusion
Half of the total plasmids of antibiotic-resistant E. coli recovered from WWTP effluent and its receiving stream water in Northwest Arkansas were transmissible as indicated by the existence of mobilization genes, with mobF12 gene as the most frequently detected on the isolates possessing in vitro resistance to one to six antibiotics. While mob and intI genes did not necessarily occur in the same plasmids, about half of antibiotic-resistant E. coli carried at least one integron gene, usually intI1. The current study signified the prevalence of single to coexistence of up to three sul genes and single or a combination up to five dfr genes in resistant E. coli, even in bacterial isolates not exhibiting in vitro resistance to either antibiotic. Having two or more mob, one or two intI, three sul, and/or a combination of two to five dfr genes each contributed to significantly increasing the proportion of the E. coli population exhibiting larger MDR numbers. Overall, these findings indicate the role of transmissible plasmid-mediated and integron-associated antibiotic (sulfamethoxazole and/or trimethoprim) resistance gene diversity among bacteria, which may contribute to persistence and dissemination of antibiotic resistance in the stream environment.
Footnotes
Acknowledgment
This work was supported by the Fulbright Program of the Bureau of Educational and Cultural Affairs (ECA) of the U.S. Department of State; USGS 104B grant administered through the Arkansas Water Resources Center (G11AP20066); Cell and Molecular Biology Interdisciplinary Program of University of Arkansas; and University of Arkansas System Division of Agriculture.
Disclosure Statement
No competing financial interests exist.