
Abstract
Fluconazole resistance among clinical Candida parapsilosis isolates is an emerging problem in many countries, including Kuwait. Resistance to fluconazole is mediated by amino acid substitutions in ERG11 and/or by overexpression of efflux pumps MDR1 and CDR1. Clinical C. parapsilosis sensu stricto isolates (n = 442) were tested for susceptibility to fluconazole by Etest, Vitek II, and broth microdilution methods. ERG11 was analyzed from fluconazole-resistant, fluconazole-susceptible dose-dependent, and selected fluconazole-susceptible isolates. Of 442 C. parapsilosis isolates, 425, 2, and 15 were identified as susceptible, susceptible dose-dependent, and resistant to fluconazole, respectively. PCR sequencing of ERG11 identified Y132F mutation in 5 of 11 fluconazole-resistant isolates available for analysis. This mutation was absent in 46 fluconazole-susceptible and 2 fluconazole-susceptible dose-dependent isolates. A multiplex allele-specific PCR was developed for detection of Y132F mutation in ERG11, and results correlated perfectly with PCR sequencing data for ERG11 codon 132 for all isolates analyzed. Detection of resistance in 15 and reduced susceptibility in 2 among 442 C. parapsilosis isolates highlights emerging resistance to fluconazole in Kuwait. The Y132F mutation in ERG11 was found in 5 of 11 (45%) fluconazole-resistant isolates only. Detection of fluconazole resistance in C. parapsilosis will help in proper management of patients infected with this species.
Introduction
T
Although echinocandins exhibit reduced in vitro activity against clinical isolates belonging to C. parapsilosis complex species, new guidelines from the Infectious Disease Society of America do not endorse fluconazole over echinocandins as primary therapy for bloodstream C. parapsilosis infections as initial use of echinocandins does not negatively influence outcome.8–12 However, fluconazole is still preferred in many countries due to its lower cost and safety profile. Resistance to fluconazole has, however, emerged in C. parapsilosis isolates in recent years with some centers reporting ∼7% isolates as resistant to fluconazole.3,8 The molecular basis of resistance to fluconazole has recently been studied in C. parapsilosis and typically involves overexpression and/or missense mutations in ERG11 encoding 14-α-sterol demethylase participating in ergosterol biosynthesis. Additional mechanisms include overexpression of efflux pumps CDR1 and MDR1 possibly due to activating mutations in their transcriptional regulators, TAC1 and MRR1, respectively.13–15 Unlike C. albicans where several missense mutations have been described in ERG11,16,17 a single missense mutation (Y132F) occurred in 31–100% of azole-resistant clinical C. parapsilosis isolates that also correlated with changes in their sterol composition, and the corresponding mutation in fluconazole-resistant C. albicans is well documented.13–15 On the contrary, missense mutations in transcriptional regulators TAC1 and MRR1 did not always correlate with overexpression of CDR1 and MDR1 and also did not correspond to documented activating mutations in these genes from C. albicans.14,15
Fluconazole resistance was not detected among clinical C. parapsilosis isolates in Kuwait until 2011.18,19 However, several fluconazole-resistant C. parapsilosis strains have been isolated in recent years. This study investigated the susceptibility of clinical C. parapsilosis isolates to fluconazole during 2012 to 2015 by Etest, Vitek II, and broth microdilution method. We also detected mutations in ERG11 gene in fluconazole-susceptible, fluconazole-susceptible dose-dependent, and fluconazole-resistant isolates. A multiplex allele-specific (MAS) PCR was also developed for rapid detection of the common mutation found in fluconazole-resistant C. parapsilosis.
Materials and Methods
Source of clinical C. parapsilosis isolates
A total of 442 C. parapsilosis isolates obtained from blood and other clinical specimens from nine hospitals in Kuwait during January 2012 to December 2015 were analyzed. These included 267 bloodstream isolates from candidemia patients and 175 isolates from other (urine, n = 43; sputum, n = 24; ear swab, n = 29; endotracheal aspirates, n = 22; line tip, n = 9; wound swab, n = 8; nasopharyngeal secretion, n = 7; rectal swab, n = 6; umbilical swab, n = 4; and other miscellaneous specimens, n = 23) anatomical sites (Table 1). The isolates were initially identified by Vitek II (bioMérieux; Marcy-l'Etoile, France) yeast identification system. C. parapsilosis sensu stricto isolates (n = 442) were initially identified by a multiplex PCR described recently by our group. 20 For this purpose, genomic DNA was prepared from each isolate as described previously. 21 PCR amplification of the internally transcribed spacer (ITS) region of ribosomal DNA region was carried out using four forward (mCPF, 5′-TTTGCTTTGGTAGGCCTTCTA-3′; mCOF, 5′-TAAGTCAACTGATTAACTAAT-3′; mCMF, 5′-AACTGCAATCCTTTTCTTTCTA-3′; and mLEF, 5′-TACAGAATTTTGAGAATTGTG-3′) and one reverse (mCPCR, 5′-AATATCTGCAATTCATATTACT-3′) primer. 20 PCR reaction and cycling conditions were same as described previously. 20 PCR amplicons were detected by agarose gel electrophoresis, performed as described previously. 21 The identity of the isolates as C. parapsilosis sensu stricto was further confirmed by PCR amplification of the ITS region of ribosomal DNA using species-specific primers (CPAF, 5′-TTTGCTTTGGTAGGCCTTCTA-3′ and CPAR, 5′-GAGGTCGAATTTGGAAGAAGT-3′), as described previously. 22 The study was approved by the Joint Committee for the Protection of Human Subjects in Research, Health Sciences Center, Kuwait University and Ministry of Health, Kuwait.
MIC, minimum inhibitory concentration; NA, not applicable.
Antifungal susceptibility testing
In vitro antifungal drug susceptibility testing was carried out against fluconazole, voriconazole, amphotericin B, caspofungin, and micafungin by Etest (AB BIODISK, Solna, Sweden) and by Vitek II system (bioMérieux) by following manufacturer's instructions and as described previously. 23 The isolates exhibiting resistance to fluconazole were also tested by broth microdilution method, as described by Clinical Laboratory Standards Institute (CLSI). 24 Revised breakpoints (minimum inhibitory concentration, MIC) of CLSI were used for fluconazole (≤2 mg/L, susceptible; 4 mg/L, susceptible dose dependent; and ≥8 mg/L, resistant), voriconazole (≤0.125 mg/L, susceptible; 0.25–0.5 mg/L, intermediate; and ≥1 mg/L, resistant), caspofungin, and micafungin (≤2 mg/L, susceptible; >2 mg/L, intermediate; and ≥8 mg/L, resistant).24,25 For amphotericin B, isolates with MIC <2.0 mg/L were considered as susceptible. Quality control was ensured using C. parapsilosis ATCC22019 and C. albicans ATCC90028.
Sequencing of the ERG11 gene
Genomic DNA prepared from C. parapsilosis isolates, as described above, was also used for PCR amplification and sequencing of ERG11 gene with specific primers, designed using the sequence of ERG11 gene from C. parapsilosis ATCC22019 (GenBank accession no. GQ302972) as reference. PCR amplification for two separate fragments was performed using CPERG11F1 (5′-CTTAGTAGTGGGATCGGTGGA-3′) and CPERG11R2 (5′-GGGTGAATCCTTTATCTAAATCA-3′) (for N-terminal fragment) or CPERG11F3 (5′-CTGCATCTAGATCCTTATTAGGA-3′) and CPERG11R4 (5′-CATTCTGCATTAAACCCCATTGA-3′) (for C-terminal fragment) primers. PCR amplification was performed in a final volume of 50 μl containing 1× AmpliTaq DNA polymerase buffer I and 1 unit of AmpliTaq DNA polymerase (Applied Biosystems, Branchburg, NJ), 10 pmol of CPERG11F1 and CPERG11R2 or CPERG11F3 and CPERG11R4 primers, 2 μl of template DNA, and 100 μM of each deoxyribonucleotide triphosphate (dNTP). Cycling conditions included an initial denaturation at 95°C for 5 min followed by 30 cycles of 95°C for 1 min, 52°C for 30 sec, and 72°C for 2 min and a final extension at 72°C for 10 min. A portion (5 μl) of PCR product was analyzed on 2% agarose-Tris-borate-EDTA gels and stained with ethidium bromide, as described previously. 21 PCR amplicons were purified using QIAquick PCR Product Purification Kit (Qiagen, Hilden, Germany) using kit instructions supplied by the manufacturer.
Each purified amplicon was sequenced in both directions for maximum coverage and reproducibility of results. N-terminal amplicons were sequenced with CPERG11F1S (5′-TAGTGGGATCGGTGGATCTT-3′), CPERG11F2 (5′-TTAGCCCTTCATGGGTACAACT-3′), CPERG11R1 (5′-GATGTTGTAAATGAAAGGAGCA-3′), and CPERG11R2S (5′-GAATCCTTTATCTAAATCAGCATA-3′) primers. C-terminal amplicons were sequenced with CPERG11F3S (5′-CTAGATCCTTATTAGGAGAAGCA-3′), CPERG11F4 (5′-AACCCTCAGATTACACATGCCA-3′), CPERG11R3 (5′-TACTTTGTGTTTGGCACAACC-3′), and CPERG11R4S (5′-TGCATTAAACCCCATTGATTCTA-3′) primers. The sequencing reactions were performed with BigDye Terminator (version 3.1) Cycle Sequencing Kit (Applied Biosystems, Inc.), as described in detail previously for fungal DNA sequencing except that ERG11 primers described above were used as sequencing primers. 26 Briefly, sequencing reactions in a final volume of 10 μl contained 2 μl purified amplicon, 1× reaction buffer, 2 μl of BigDye terminator (version 3.1) sequencing reagent, and 3.2 pmol of sequencing primer. Temperature cycling parameters included an initial denaturation step at 95°C for 1 min followed by 30 cycles of 1 min at 95°C and 4 min at 60°C. The unincorporated terminators were removed from sequencing reactions by using BigDye XTerminator Kit (Applied Biosystems, Inc.) according to instructions supplied with the kit. The processed samples were loaded on an ABI 3130xl Genetic Analyzer (Applied Biosystems, Inc.) for electrophoresis and data collection according to the manufacturer's instructions. 26
The DNA sequence data generated by the ABI 3130xl Genetic Analyzer were checked for confidence levels with an ABI sequence scanner; reverse compliments were generated and aligned with forward sequences using Clustal Omega (www.ebi.ac.uk/Tools/msa/clustalo). 26 Nucleotide and amino acid sequences were compared with wild-type sequence from C. parapsilosis ATCC22019 (GenBank accession no. GQ302972) using Clustal Omega.
MAS-PCR for detection of Y132F mutation in ERG11
MAS-PCR assay was developed for detecting Y132F mutation in ERG11 using three primers (CPERG11-132F, 5′-TATCTTATGGACAGGATCCATA-3′; CPERG11-132R, 5′-ATAATCCAAAATTTCCCCTCTAA-3′; and CPERG11-132Phe, 5′-GTCTTGCATTCGGACAATCGAA-3′) in PCR. MAS-PCR was performed and amplicons were detected as described previously, 27 except that 10 pmol of CPERG11-132F and CPERG11-132R primers and 1 pmol of CPERG11-132Phe primer were used, and annealing of primers was set at 60°C for 30 sec. MAS-PCR should yield only one (323 bp) fragment from C. parapsilosis containing TAC (Y132) and two DNA fragments of 323 and 231 bp from isolates containing TTC (Y132F mutation) at ERG11 codon 132.
Results
Of 442 individual patient C. parapsilosis sensu stricto isolates collected during 2012 to 2015, 15 were resistant (MIC ≥ 8 mg/L), 2 were susceptible dose dependent (MIC = 4 mg/L), and 425 were susceptible to fluconazole (Table 1). All fluconazole-resistant C. parapsilosis isolates were also confirmed by Vitek II and broth microdilution method. The first fluconazole-resistant C. parapsilosis strain (Etest MIC = 16 mg/L) was isolated from a candidemia patient in 2012. The MIC range by Etest of the remaining blood isolates in 2012 (n = 37) was 0.004–1.5 mg/L. In 2013, none of the C. parapsilosis isolates from blood (n = 59) showed MIC of >3 mg/L (MIC range 0.094–3 mg/L), and none of the nonblood isolate was also resistant to fluconazole. In 2014, 3 of 91 (3.3%) C. parapsilosis bloodstream isolates were resistant to fluconazole (MICs 8–16 mg/L). In 2015, fluconazole resistance (MIC range 12 ≥ 256 mg/L) was detected in 4 of 79 (5.1%) blood isolates, but was also detected among 7 of 50 (14%) nonblood isolates (MIC range 8 ≥ 256 mg/L) for the first time. Of 15 fluconazole-resistant isolates, 4 were lost during storage and subculturing. Thus, only 11 fluconazole-resistant isolates were available for further studies, including molecular characterization of fluconazole resistance-conferring mutations in ERG11. In addition, 2 susceptible dose-dependent and 16 selected fluconazole-susceptible isolates were also tested initially for ERG11 mutations.
The ERG11 coding sequences were 1,569 bp from all 29 isolates. In addition, 5′-untranslated region of 449 bp was also amplified and sequenced. By comparison to C. parapsilosis ATCC22019 sequence, one single-nucleotide polymorphism (SNP) (T591C) resulting in a silent mutation (T197T) was detected in all 29 isolates from Kuwait, while five different SNPs were detected in 12 isolates. All point mutations were homozygous (mutations in both alleles). Two SNPs, C-111T in the 5′-untranslated region and G1193T (R398I), were found together in seven (two fluconazole susceptible and five fluconazole resistant) isolates (Table 2). One SNP, A395T (Y132F), was found together with two other (C-111T and G1193T) SNPs in five resistant isolates only. All five (one each from blood in 2012 and 2014 and three, including two from blood, in 2015) isolates with identical ERG11 sequences were obtained from patients at Farwaniya Hospital in Kuwait (Table 2). Interestingly, 5 of 11 fluconazole-resistant isolates were also resistant (MIC of ≥1 mg/L) and 4 were intermediate (MIC of 0.25–0.5 mg/L) to voriconazole. 25 All 29 isolates were susceptible to amphotericin B (MICs <2.0 mg/L) and to both echinocandins (MICs <2 mg/L) (Table 2).
The MIC values read at 24 and 48 hr were nearly identical for all antifungal drugs.
All isolates contained one SNP (T591C) which resulted in a silent (T197T) mutation in all 29 isolates compared to the wild-type sequence from reference Candida parapsilosis ATCC22019.
AMB, amphotericin B; CFG, caspofungin; ETA, endotracheal aspirate; FLU, fluconazole; MAK, Mubarak Al-Kabeer Hospital; MFG, micafungin; MIC, minimum inhibitory concentration; ND, not done; PF, pleural fluid; SNP, single-nucleotide polymorphism; VRC, voriconazole.
Considering strong association of Y132F mutation in ERG11 with fluconazole resistance in C. parapsilosis,13–15 a MAS-PCR assay was developed as a rapid screen for fluconazole-resistant C. parapsilosis. C. parapsilosis (ATCC22019) containing wild-type ERG11 codon 132 (Y132) was used as reference strain. MAS-PCR accurately detected the presence of TAC in 16 fluconazole-susceptible, 2 susceptible dose-dependent, and 6 of 11 fluconazole-resistant C. parapsilosis isolates containing wild-type sequence at ERG11 codon 132 as they yielded only one (323 bp) amplicon (data from one isolate are shown in Fig. 1, lane Y132). MAS-PCR detected TTC at ERG11 codon 132 in all five fluconazole-resistant isolates containing Y132F mutation as they yielded two amplicons of 323 and 231 bp (data from one isolate are shown in Fig. 1, lane F132). The sensitivity and specificity of MAS-PCR assay were further studied using 30 additional fluconazole-susceptible C. parapsilosis isolates described in this study. MAS-PCR accurately detected wild-type sequence (Y132) at ERG11 codon 132 in all 30 fluconazole-susceptible isolates.
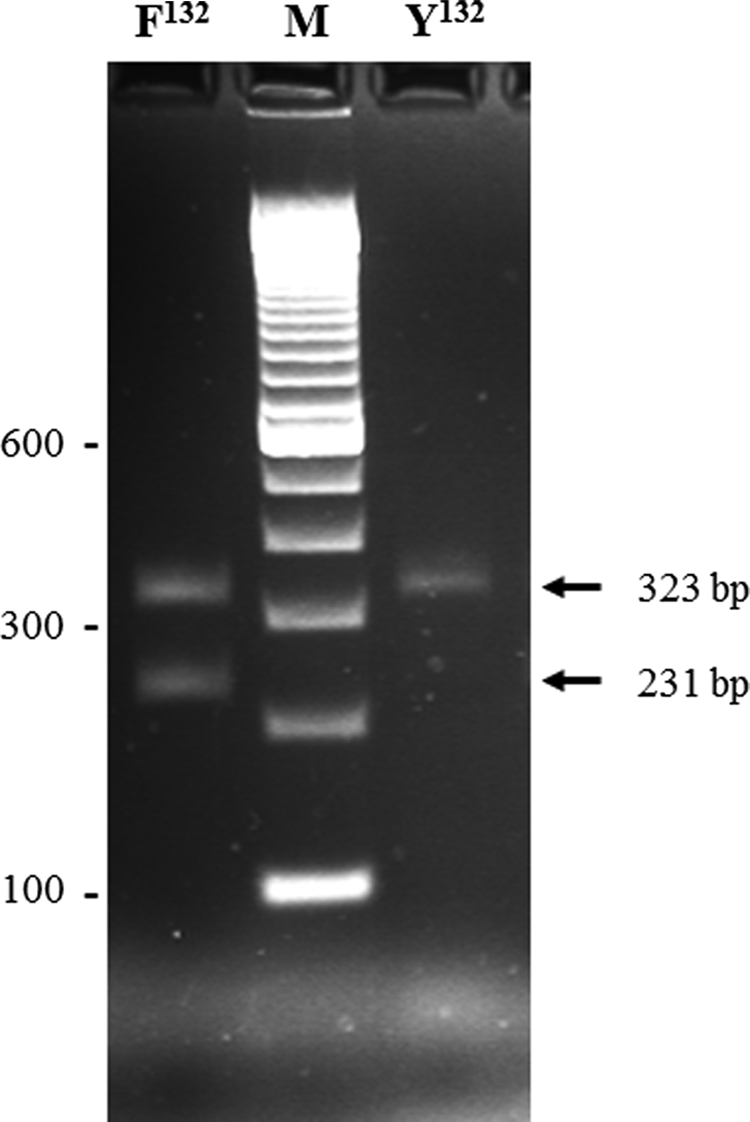
Agarose gel of multiplex allele-specific PCR amplicons obtained from fluconazole-susceptible isolate Kw1912-15 (lane Y132) and fluconazole-resistant Candida parapsilosis isolate Kw2006-15 (lane F132) obtained with CPERG11-132F, CPERG11-132R, and CPERG11-132Phe primers. The position of migration of 323 and 231 bp fragments is indicated by arrows. Lane M is 100 bp DNA ladder, and the positions of migration of 100, 300, and 600 bp fragments are marked.
Discussion
C. parapsilosis is a healthcare-associated pathogen affecting immunocompromised patients of all age groups. Drug-resistant clones of C. parapsilosis have the capacity to persist in nosocomial settings for prolonged periods and have caused outbreaks. 28 Although fluconazole has been widely used in Kuwait for >20 years, resistance in C. parapsilosis against this drug has been observed only since 2012 when the first fluconazole-resistant C. parapsilosis strain was isolated from a female candidemia patient. It could not be ascertained if the patient had ever received fluconazole therapy locally or systemically before resistance developed or whether she had acquired a fluconazole-resistant strain. Surprisingly, none of the resistant bloodstream isolates came from a neonate or a pediatric patient. The long-term use of fluconazole to control C. parapsilosis bloodstream infections in neonatal intensive care unit in Finland also resulted in the emergence of a fluconazole-resistant strain that caused cross-infections over a 12-year period. 29 These findings suggest that acquisition of resistance to fluconazole has been a slowly evolving phenomenon.10,24,30 The analyses of our fluconazole susceptibility data also suggest that the frequency of isolation of fluconazole-resistant C. parapsilosis is increasing in Kuwait as most (11 of 15) fluconazole-resistant isolates were obtained in 2015, including four of eight isolates from blood. Similar findings have also been noted at other geographical locations.31,32 The increasing frequency of fluconazole resistance may become a more challenging therapeutic issue in future considering that echinocandins, the preferred first-line drugs for treatment of candidemia and invasive candidiasis, exhibit higher MICs against C. parapsilosis compared to other Candida species, and instances of “breakthrough” C. parapsilosis infections have been reported in patients who received echinocandin therapy.31–35
PCR sequencing of ERG11 coding and noncoding sequences from 11 fluconazole-resistant, 2 susceptible dose-dependent, and 16 selected fluconazole-susceptible C. parapsilosis isolates showed that only one nonsynonymous mutation (A395T) leading to replacement of tyrosine by phenylalanine at codon 132 (Y132F) was present exclusively in 5 of 11 fluconazole-resistant isolates, while other SNPs were found in both susceptible and resistant isolates. The mutation A395T was also associated with two other SNPs (C-111T and G1193T leading to R398I substitution) in fluconazole-resistant C. parapsilosis isolates in Kuwait. While 9 of 9 (100%) fluconazole-resistant isolates from Brazil contained Y132F mutation alone, 13 Y132F was present either alone or in combination with C-111T and/or R398I in fluconazole-resistant isolates from United States.14,15 In the study from Memphis, Tennessee, 11 of 35 (31.4%) fluconazole-resistant isolates contained Y132F mutation, including 10 isolates with Y132F + R398I mutation (data on C-111T mutation was not reported possibly due to close proximity of this location to PCR amplification primers), while R398I mutation was also detected among fluconazole-susceptible isolates. 15 The Y132F mutation was detected in 17 of 30 (57%) fluconazole-resistant isolates from Atlanta and Baltimore areas in United States. 14 Although not specifically stated, 6 of 17 isolates also likely contained C-111T and R398I mutations. 14 Similar to our study, C-111T and R398I mutations were also detected in 17 of 55 susceptible isolates, as well as in 13 of 37 susceptible dose-dependent isolates. 14 One SNP, T534C (M178T), was found in four (one susceptible, one susceptible dose-dependent, and two resistant) isolates, while another SNP, A847T, leading to N283Y substitution was found in one susceptible isolate only. Previously, these mutations have been described in susceptible dose-dependent isolates only from the United States. 14 Our data show that mutations M178T and N283Y do not confer resistance to fluconazole in C. parapsilosis.
Considering strong association of Y132F mutation in ERG11 with fluconazole resistance in C. parapsilosis,13–15 a MAS-PCR assay was developed as a rapid screen for fluconazole-resistant C. parapsilosis. MAS-PCR accurately detected the presence of TAC in 46 fluconazole-susceptible, 2 susceptible dose-dependent, and 6 of 11 fluconazole-resistant C. parapsilosis isolates containing wild-type sequence at ERG11 codon 132. The MAS-PCR also detected TTC at ERG11 codon 132 in all five fluconazole-resistant isolates containing Y132F mutation. It should be emphasized in this study that MAS-PCR is only designed for detection of Y132F mutation at ERG11 codon 132 and isolates lacking this mutation but containing other ERG11 mutations may yield false-negative results for fluconazole resistance. It is also pertinent to mention in this study that sensitivity of multiplex PCR assay will vary at different geographical locations according to the incidence of Y132F mutation among fluconazole-resistant C. parapsilosis isolates, which has so far been reported to vary (including this study) from 31% to 100%.13–15
Our study has a few limitations. The information on prior therapy with fluconazole for the patients yielding fluconazole-resistant C. parapsilosis isolates was not available. The role of efflux pumps in conferring resistance to fluconazole, particularly in isolates lacking a resistance-linked ERG11 mutation, was not studied. Another limitation of our study is that high-resolution fingerprinting was not carried out for isolates with Y132F mutation, and surveillance cultures to ascertain the source of fluconazole-resistant C. parapsilosis infections at Farwaniya Hospital were not available.
In conclusion, our fluconazole susceptibility data show that the frequency of isolation of fluconazole-resistant C. parapsilosis is increasing in Kuwait. The Y132F mutation in ERG11 was found in 5 of 11 (45%) fluconazole-resistant isolates, but was absent in fluconazole-susceptible and fluconazole-susceptible dose-dependent isolates analyzed simultaneously. MAS-PCR was also developed for rapid detection of Y132F mutation in ERG11 and may help in proper management of patients infected with fluconazole-resistant C. parapsilosis isolates.
The DNA sequencing data reported in this study have been submitted to European Molecular Biology Laboratory/GenBank databases under accession nos. LT596074 to LT596093.
Footnotes
Acknowledgments
Cooperation received from clinical microbiologists working in different hospitals of Kuwait is thankfully acknowledged. The study was supported by Kuwait University Research Sector (Grant No. YM 10/11) and Research Core Facility (Grant No. SRUL02/13).
Disclosure Statement
No competing financial interests exist.