
Abstract
Objective:
The current investigation explores whether extended-spectrum β-lactamase (ESBL) genes exist in clinical non-ESBL–producing Klebsiella pneumoniae isolates.
Methods:
A total of 202 clinical isolates with non-ESBL–producing K. pneumoniae were collected from southern and middle of China. Thirteen β-lactamase genes (blaSHV, CTX-M, TEM, OXA-2, OXA-10, VEB, PER, SFO, GES, CSP, TLA, BEL, and IBC) were screened by PCR and their identity confirmed by sequencing of PCR products. The ESBL-producing phenotype of the isolates that carried ESBL genes was tested and confirmed in 9 of the 18 isolates by a double-disc synergy test. The sequences upstream of ESBL genes of isolates with ESBL-producing genotype (+)/phenotype (−) were also subjected to PCR and sequencing. The ESBL genes and their upstream regions were cloned into Escherichia coli DH5α for functional evaluation.
Results:
A total of 8.9% (18/202) isolates carried ESBL genes. All of them harbored only one ESBL gene, including 33.3% (6/18) blaSHV and 66.7% (12/18) blaCTX-M. Among the isolates carrying ESBL genes, nine isolates were confirmed as ESBL phenotype (−). The ESBL genotype (+)/phenotype (−) isolates had blaSHV-27,38,41,42 (66.7%, 6/9) and blaCTX-M-3,15,24 (33.3%, 3/9). The upstream gene sequences, including promoters of these unexpressed ESBL genes, were intact without any mutations or spacers and effective among eight strains. The ISEcp1 element in the upstream region was not found in one isolate carrying an unexpressed blaCTX-M-15 gene.
Conclusions:
Clinical non-ESBL–producing K. pneumoniae isolates could carry ESBL genes with intact promoter, but without the correlated phenotype. Specific silencing mechanisms may play an important role in regulating ESBL gene expression. This kind of isolates has the potential to transfer their ESBL genes to other bacteria with effective promoters, resulting in ESBL phenotype.
Introduction
K
ESBLs are defined as β-lactamases (bla) belonging to molecular class A and functional group 2be.4,5 ESBLs typically hydrolyze cephalosporins and aztreonam, but neither cephamycins nor carbapenems. However, β-lactamase inhibitors, such as clavulanic acid (CA), are able to inhibit this enzymatic hydrolysis.6,7 The majority of researchers focus on antibiotic-resistant K. pneumoniae rather than antibiotic-sensitive bacteria that are usually ESBL negative in both phenotype and genotype. However, some studies have demonstrated that antibiotic-resistant genes could be present, but are unexpressed in numerous antibiotic-sensitive species.8–12 Our group has also previously reported unexpressed blaSHV ESBL gene in K. pneumoniae. 13
The current investigation explores whether ESBL genes exist in clinical non-ESBL–producing K. pneumoniae isolates. Specifically, we collected clinically isolated antibiotic-sensitive K. pneumoniae isolates and used the VITEK ESBL detection system to screen for ESBL production. Then, non-ESBL–producing K. pneumoniae isolates were screened for a wide range of ESBL genes. Furthermore, the promoter regions of the unexpressed ESBL genes were examined. Our findings have important implications for clinical antimicrobial medication (Fig. 1).
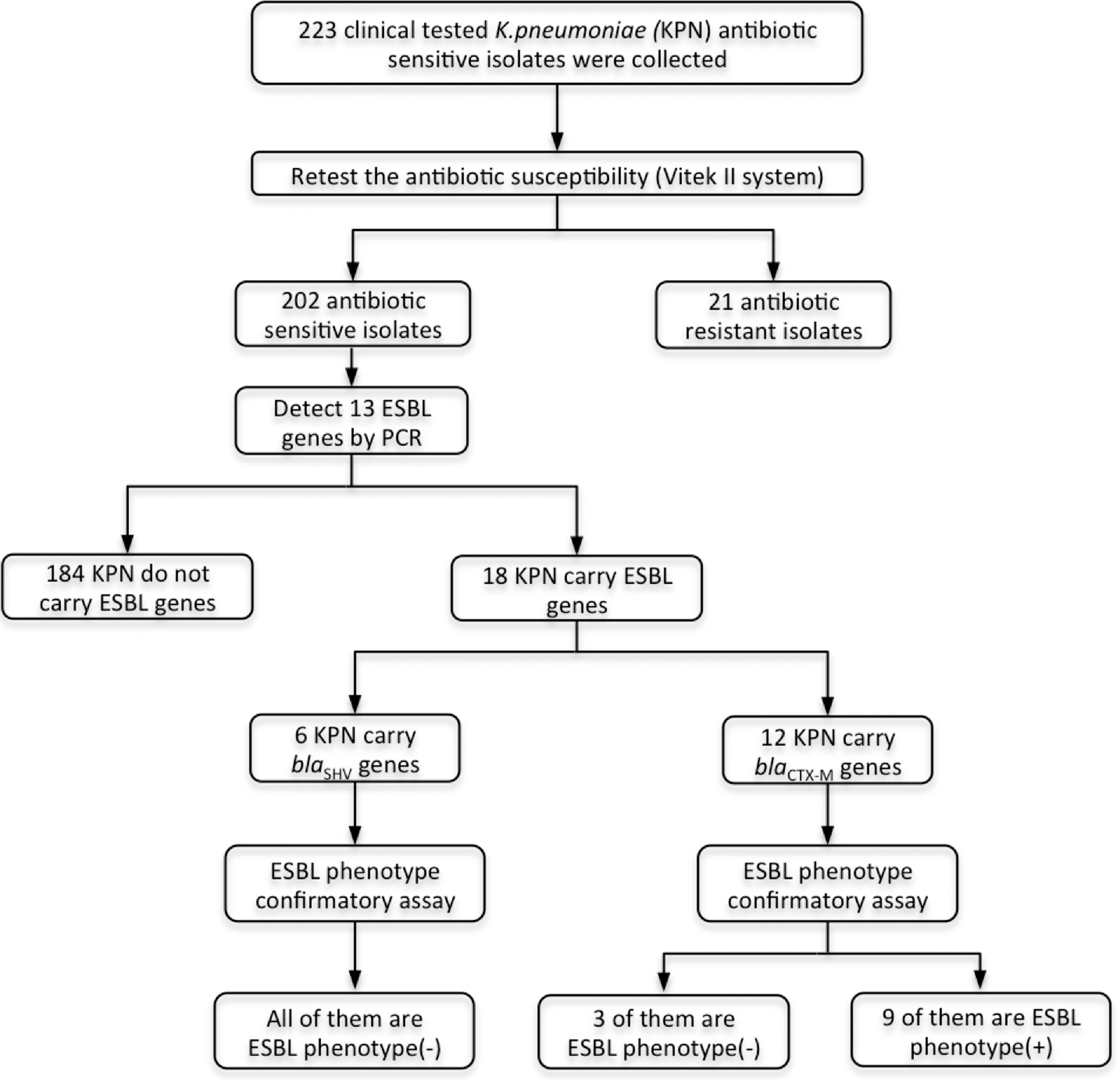
Two hundred twenty-three clinically tested antibiotic sensitive KPN were collected; unexpressed ESBL genes were detectable in 18 KPN finally (+, positive signal; −, negative signal). ESBL, extended-spectrum β-lactamase; KPN, Klebsiella pneumoniae.
Materials and Methods
Bacterial collection and identification
Clinical K. pneumoniae isolates were collected from Fujian Provincial Hospital and the First Affiliated Hospital of Zhengzhou University from 2014 to 2015. The clinical specimens of the patients with infectious diseases were gathered. The isolates were from urine, blood, secretions, sputum, and cerebrospinal fluid with the aid of chromogenic medium initially. Then, the isolates were identified, and the isolates' antimicrobial susceptibilities and ESBL phenotype were tested by the VITEK-2 ESBL detection system. A total of 202 non-ESBL–producing K. pneumoniae isolates were identified; these microorganisms were enrolled in our study and stored at −80°C.
PCR screening to detect ESBL genes in non-ESBL–producing isolates
PCR amplification was used to screen the blaSHV, blaCTX-M, blaTEM, blaVEB, blaGES, blaIBC, blaSFO, blaCSP-1, blaTLA, blaBEL, blaPER, blaOXA-10, and blaOXA-2 genes.14–20 The primers and their sequences are shown in Table 1. The groups within the CTX-M were identified using specific primers for CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9, and CTX-M-25 groups.14,21–23 The primers of these genes were used individually to screen 202 antibiotic sensitive isolates. The PCR products of all ESBL genes were then submitted to Invitrogen (Thermo Fisher Scientific, Inc.) for bidirectional nucleotide sequencing. Analysis of the DNA sequences was performed using the Basic Local Alignment Search Tool (BLAST) (www.ncbi.nlm.nih.gov/BLAST) and the nomenclature website for ESBLs (www.lahey.org/studies). Genotype positive isolates were amplified and sequenced thrice.
Phenotype confirmation of ESBL genotype positive isolates
The genotype positive isolates were tested for ESBL production by double-disc synergy test (DDST) on Mueller–Hinton agar plates in accordance with the guidelines of Clinical Laboratory Standards Institute (CLSI). The antibiotics used were ceftazidime (CAZ), ceftazidime–clavulanic acid (CAZ/CA), cefotaxime (CTX), and cefotaxime–clavulanic acid (CTX/CA). ESBL production was phenotypically confirmed by an increase of ≥5 mm in the zone diameter for CAZ and/or CTX in combination with CA versus its zone diameter when tested alone. All discs were purchased from Beijing Tiantan Biological Products Corporation (China). Escherichia coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality-control strains. Results were interpreted in accordance with the CLSI guidelines.
Detection of the upstream genetic environment of the genotype positive but non-ESBL–producing isolates
The upstream environmental background of non-ESBL–producing K. pneumoniae isolates carrying ESBL genes was further investigated. We amplified the resistance genes and their upstream regions using specific primers. 24 Primers IS26-FCJ and RECF-F for the insertion sequences of IS26 and truncated region of the recF gene, respectively, were used with SHV-R to test whether the promoter regions upstream of the blaSHV ESBL genes were intact. We also used primer ISEcpUP for ISEcp1 insertion sequences paired with CTX-M-1-R and CTX-M-9-R primers to examine the promoter regions upstream of blaCTX-M-3,15,24 genes. PCR products were sequenced and analyzed as described above.
Transformation of the ESBL gene expression system
To investigate the functionality of the expression system of the isolates that were genotype positive but phenotype negative, the ESBL genes were cloned with their promoters into the vector plasmid pUA66 (kindly provided by Uri Alon from the Weizmann Institute of Science in Israel) and transformed into competent E. coli DH5α (Tiangen Biotech Co. Ltd., Beijing, China) (Fig. 2). BlaSHV genes and the promoter sequences were amplified using primers RECF-F (BglII) with SHV-R (XhoI); blaCTX-M genes and the promoter sequences were amplified using primers ISEcpUP (BglII) with CTX-M-1-R (XhoI) and CTX-M-9-R (XhoI), respectively (Table 2). The PCR products were then sequenced, and the sequences were analyzed using sequencing alignment software DNAMAN to confirm their 100% identity with sequences from the parent isolates.

The blaCTX-M genes and their upstream sequence in non-ESBL producer isolates were amplified and sequenced. Then they were inserted into BglII and XhoI site of plasmid pUA66. The plasmids were introduced into Escherichia coli DH5α competent cells; the recombinants were used to test the ESBL phenotype.
After that, ESBL genes and upstream sequences were inserted into the BglII and XhoI sites of the vector plasmid pUA66 that contained 4,260 bp and lacked any promoter regions driving expression of ESBL genes. The fragments and plasmid pUA66 were digested using the restriction endonucleases BglII and XhoI at 37°C for 1 hour, after which they were ligated at 16°C overnight. The constructed plasmids were introduced into competent E. coli DH5α cells by the heat shock method. The ESBL genes in recombinants were tested by PCR and sequenced to confirm that there were not any mutations. The empty vector pUA66 was also transformed into DH5α, and the recombinant was named pUA66. Recombinants with ESBL genes and their promoter sequences were named pUA66-R27, pUA66-R38, pUA66-R41, pUA66-R42, pUA66-PIC3, and pUA66-PIC24. blaCTX-M-3 and 24 were cloned into DH5α alone without upstream regions as described above, and the resulting recombinants were named pUA66-C3 and pUA66-C24.
The ESBL phenotype and antibiotic susceptibilities of transconjugant E. coli isolates were tested by DDST and VITEK. E. coli ATCC 25922 and K. pneumoniae ATCC 700603 were used as quality control strains. Results were interpreted in accordance with the CLSI guidelines.
Results
Bacterial collection and identification
A total of 223 K. pneumoniae clinical isolates were collected, including those from Fujian Provincial Hospital (n = 139) and the First Affiliated Hospital of Zhengzhou University (n = 84). The ESBL phenotypes of these isolates were screened based on their susceptibilities to ceftazidime (CAZ), ceftriaxone, and cefepime tested by VITEK. Among the isolates, 21 were identified as ESBL-producing K. pneumoniae isolates. The ESBL genes were screened in the remaining 202 K. pneumoniae isolates.
Characterization of ESBL genes harbored by antibiotic-sensitive K. pneumoniae isolates
Among the 202 K. pneumoniae isolates, 18 (8.9% 18/202) carried ESBL genes, either blaSHV or blaCTX-M ESBL gene. All of the 18 strains carried only one ESBL gene each, including blaSHV-27,38,41,42 (33.3%, 6/18) and blaCTX-M-3,14,15,24,55,65 (66.7%, 12/18), and genes encoding three novel SHV β-lactamases identified as SHV-190, -191, and -193, which are now included in www.lahey.org/studies/webt.asp
Confirmation of ESBL phenotype
Of the 18 isolates carrying ESBL genes, half (9/18) were non-ESBL–producing isolates by the result of DDST. The details of these nine isolates are listed in Table 3. In these ESBL genotype (+)/phenotype (–) isolates, 66.7% (6/9) carried the blaSHV genes, whereas 33.3% (3/9) contained the blaCTX-M genes. As shown in Table 3, Z42, F13, and F96 carried blaSHV-27 (reference sequences: GenBank accession no. AF293345); F73 carried blaSHV-38 (reference sequences: GenBank accession no. EU979559); Z47 carried blaSHV-41 (reference sequences: GenBank accession no. AF535129); F63 carried blaSHV-42 (reference sequences: GenBank accession no. AF535130); F18 carried blaCTX-M-3 (reference sequences: GenBank accession no. AB059404); F89 carried blaCTX-M-24 (reference sequences: GenBank accession no. AY143430); and F93 carried blaCTX-M-15 (reference sequences: GenBank accession no. JN627489).
ESBL, extended-spectrum β-lactamase.
Upstream promoter regions of non-ESBL–producing isolates
The nine isolates with ESBL genotype (+)/phenotype (−) were further investigated. The promoter regions of blaSHV genes are given by recF gene, while the ISEcp1 insertion sequence is present in upstream region of blaCTX-M genes. (Supplementary Data; Supplementary Data are available online at www.liebertpub.com/mdr). The upstream genetic environments of the blaSHV genes in the six isolates were identified as recF gene containing the promoter sequences (−35 box: 5′–TTGATT–3′; −10 box: 5′–ACAAAT–3′) of these blaSHV genes, but without IS26 insertion sequences. The upstream region of blaCTX-M-3,24 carrying isolates was confirmed as the moving element ISEcp1 containing the promoter sequences (−35 box: 5′–TTGACT–3′; −10 box: 5′–TTTCGT–3′) of these blaCTX-M genes. F18 contained a 127-bp long conserved sequence between the ISEcp1 element and the blaCTX-M-3 gene. By contrast, F89 harbored a 42-bp long conserved sequence between the ISEcp1 element and blaCTX-M-24 gene. However, ISEcp1 was not detected in F93. The genetic environment of these isolates with unexpressed blaCTX-M-15 gene must be further explored.
Transformation of the ESBL gene expression system
Since the upstream region could not be detected in F93, the promoters and ESBL-coding regions from each of the remaining eight isolates (F13, F18, F63, F73, F89, F96, Z42, and Z47) were cloned into a vector (pUA66) and transferred into recipient E. coli isolates. The ESBL-producing phenotype in the recipient isolates was tested as mentioned above. All six blaSHV recombinants were non-ESBL producers. However, when blaCTX-M genes were cloned into DH5α isolates with their promoter sequences, the recombinants exhibited the ESBL-producing phenotype. Then blaCTX-M genes alone were cloned into recipient DH5α isolates as described. The recombinants retained non-ESBL traits (Table 4, Supplementary Table S1).
pUA66-R27, pUA66+promoter+SHV-27; pUA66-R38, pUA66+promoter+SHV-38.
pUA66-R41, pUA66+promoter+SHV-41; pUA66-R42, pUA66+promoter+SHV-42.
pUA66-C3, pUA66+CTX-M-3; pUA66-IC3, pUA66+promoter+CTX-M-3.
pUA66-C24, pUA66+CTX-M-24; pUA66-IC24, pUA66+promoter+CTX-M-24.
CAZ, ceftazidime; CAZ/CA, ceftazidime–clavulanic acid; CTX, cefotaxime (CTX), cefotaxime–clavulanic acid (CTX/CA).
During our investigation, blaCTX-M genes were detected in nine “sensitive” isolates, but six of them were ESBL producers when we retested the ESBL phenotype by the DDST method. The differences between VITEK 2 and DDST should not be ignored. Garrec et al. compared nine phenotypic methods for detection of ESBL production Enterobacteriaceae. They reported that the sensitivities of the VITEK-2 system were 73–79% and that the DDST method provides better results. 25 As we know, VITEK systems are typically used in diagnostic laboratories. The false-negative results given by VITEK can mislead antimicrobial treatment outcomes. A number of studies have been conducted to compare the accuracy of VITEK with those of other methods, and most have found that VITEK is stable and reliable.25–28 In our opinion, VITEK results may be considerably influenced by the standardization of the operating procedure. Thus, doctors must remain cautious when interpreting its results.
Our group previously reported a silent blaSHV-41 gene in an antibiotic-sensitive K. pneumoniae isolate. 13 The upstream region of that K. pneumoniae isolate was the same as that obtained in this study, namely recF. Rice et al., found that when A in the second position of the −10 box was changed to C, the blaSHV downstream of the altered −10 box produced less β-lactamase enzymes. 29 Recently, our group reported that different kind of promoters may lead blaSHV-ESBL phenotypic transformation in K. pneumoniae and blaSHV-28,36,38, and 42 may be not ESBL genes. 30 However, no information has been published to date on unexpressed blaCTX-M genes in K. pneumoniae.
Two unexpressed blaCTX-M genes became ESBL producers by associating with E. coli DH5α recombinants after they were cloned into pUA66 plasmids. However, the mechanism by which the blaCTX-M genes keep silencing in K. pneumoniae remains uncertain. Some drug-resistant genes were silenced because of missing promoter regions. Examples include the methicillin-resistant mecA gene of Staphylococcus sciuri 31 and the carbapenem-resistant cfiA gene of Bacteroides fragilis. 32 Isolates with resistance genes are sensitive to the antibiotic because of their distinct lack of promoters necessary for gene expression. We found one isolate that carried blaCTX-M-15 without the ISEcp1 element, that is, F93.
Meanwhile, other two isolates harbored ISEcp1 + blaCTX-M-3 and ISEcp1 + blaCTX-M-24, respectively. The ISEcp1 is commonly identified to exist upstream of the blaCTX-M genes. This element could provide a promoter for blaCTX-M expression and also contribute to blaCTX-M gene transference. Thus, the genetic environment of this blaCTX-M-15 gene must be further investigated, for instance, by replacing a functional ISEcp1 in the upstream region. In contrast, the secretion system may also play a role in non-ESBL–producing isolates. To further investigate the mechanism underlying lack of expression of the blaCTX-M gene, we will analyze the whole genome by comparing these three isolates with that of some other K. pneumoniae that can express blaCTX-M genes well.
In the early 1990s, antibiotic resistance was mostly caused by SHV and TEM β-lactamases. 33 From such period to the last two decades, CTX-M has been becoming the most prevalent type in the ESBL family, especially in K. pneumoniae and E. coli. 33 A chromosome-encoded intrinsic gene in Kluyvera spp. was thought to be the progenitor of blaCTX-M genes. 34 This gene could be transferred to other bacteria when the upstream sequence is replaced by ISEcp1 or ISCR1. blaCTX-M gene transfer was achieved with the help of mobile element ISEcp1. 35
In our study, we identified two blaCTX-M genes that were silenced in K. pneumoniae isolates. When these two genes were cloned into competent E. coli DH5α with their promoter sequences, the recombinants became ESBL producers. This result indicated that blaCTX-M gene expression was influenced in the parent isolates. Isolates that carry resistance genes without expressing the associated resistant phenotypes are rarely noticed given our focus on antibiotic resistant pathogens. Once blaCTX-M genes were transferred to other isolates through plasmid transference or with help of ISEcp1, the recipient bacteria will carry ESBL gene and may produce ESBL. Thus, the non-ESBL–producing isolates in our study could potentially spread their antibiotic genes throughout the wards and cause an antibiotic-resistant bacteria outbreak.
In summary, this study confirmed an uncommon phenomenon in which clinical non-ESBL–producing K. pneumoniae isolates carried ESBL genes with intact promoter and gene sequences but without the correlated phenotype. Unexpressed resistance genes may be silenced by a specific regulating element and activated by frequent antibiotic pressure. However the mechanisms that underlie this silencing phenomenon need to be further elucidated. This will contribute to exploration of therapeutic strategies that target infectious diseases caused by multidrug-resistant bacteria.
Footnotes
Acknowledgments
The authors thank all the participant hospitals for collecting isolates for this study. The authors thank the team of curators of the nomenclature website for ESBLs for making them publicly available at ![]() The study was funded by National Natural Science Foundation of China (81470207) and National Key Research & Development Program of China from Ministry of Science and Technology (2016YFC0903800).
The study was funded by National Natural Science Foundation of China (81470207) and National Key Research & Development Program of China from Ministry of Science and Technology (2016YFC0903800).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.