
Abstract
Vancomycin-resistant enterococci (VRE) have been detected in wild animals representing a public health concern. The red-legged partridge (Alectoris rufa) is a common game bird and its meat is consumed in several countries, including Portugal. Three hundred five fecal samples of red-legged partridge from the north of Portugal were screened for VRE. Samples were cultured on Slanetz-Bartley agar supplemented with vancomycin (4 mg/L) and six vanA-Enterococcus faecium were recovered. Isolates were tested for antibiotic resistance and virulence genes. Multilocus sequence typing (MLST) was performed to study the genotypic diversity of vanA-containing E. faecium. The six isolates showed erythromycin resistance and harbored the erm(B) gene and the four that were tetracycline resistant showed the tet(M) gene. The C-terminal region of the pbp5 gene of the ampicillin-resistant isolates (minimal inhibitory concentration range of 256 μg/ml) was sequenced. Two different pbp5 alleles were detected when considering the changes of amino acid in 461–629 region. All isolates harbored the esp gene, whereas hyl, together with the esp gene, was detected in five isolates. MLST analysis grouped the isolates as ST448 (n = 1), ST139 (n = 1), and ST18 (n = 4). Our findings show that the red-legged partridges could be a reservoir of antimicrobial resistance genes and may contribute to the dissemination and transference of the resistance genes to other animals and humans.
Introduction
T
The objective of this study was to analyze the prevalence and molecular characterization of vanA-containing Enterococcus in fecal samples of partridges (Alectoris rufa) in Portugal.
Materials and Methods
Samples and bacterial isolates
Three hundred five wild red-legged partridges (A. rufa) were captured in the north of Portugal by hunting associations during the hunting season from October 2015 to January 2016. The partridge hunting is supervised by the Agriculture, Rural Development and Fishery Ministry of Portugal under the Decree-Law no. 201/2005. The partridges were hunted in their native lands, where they live and reproduce freely in the wild. The fecal samples (one per animal) were obtained after the necropsy from the colon of each partridge. Each sample was individually stored in plastic sterile tubes and transferred to peptone water. The samples were processed within 24 hr in the laboratory. To avoid isolation of intrinsically resistant enterococcal species, the samples were spread onto Slanetz-Bartley agar (Oxoid) plates supplemented with 4 mg/L of vancomycin and incubated at 37°C for 48 hr.
Colonies with typical enterococcal morphology were identified by Gram-staining, cultural characteristics, bile-aesculin reaction, biochemical tests using the API ID20 Strep system, and catalase test (BioMérieux, La Balme Les Grottes, France). The polymerase chain reaction (PCR) technique was performed to confirm species identification, using suitable primers and conditions for the most common species of Enterococcus. 11
Antimicrobial susceptibility
Antimicrobial susceptibility was tested by the disk diffusion method according to the criteria of the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2017) for enterococci. Eleven antibiotics were tested: ampicillin (10 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), quinupristin–dalfopristin (15 μg), streptomycin (300 μg), tetracycline (30 μg), teicoplanin (30 μg), vancomycin (30 μg), erythromycin (15 μg), gentamycin (120 μg), and kanamycin (120 μg). High-level resistance was considered for aminoglycosides. Enterococcus faecalis strain ATCC 29212 and Staphylococcus aureus strain ATCC 25923 were used for quality control in the susceptibility assays. Minimal inhibitory concentrations (MICs) of teicoplanin, vancomycin, and ampicillin were determined by the agar dilution method and by using the susceptibility breakpoints recommended by the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2017).
Antimicrobial resistance and virulence genes
The presence of antibiotic resistance and virulence genes was studied by PCR using specific primers (Sigma-Genosys Ltd., Cambridge, United Kingdom) as previously described.8,11,12 The resistant enterococci were tested for detection of the following resistance genes: van(A), van(B), and van(C) (vancomycin resistance); erm(A), erm(B), and erm(C) (erythromycin resistance); tet(L) and tet(M) (tetracycline resistance); ant(6′)-Ia, acc(6′)-Ie-aph(2″)-Ia, and aph(3′)-IIIa (aminoglycoside resistance); vat(D) and vat(E) (quinupristin–dalfopristin resistance). The study of pbp5 gene was performed by PCRnova and the products were purified with the QIAquick PCR purification kit (Qiagen, Inc.) according to the instructions of the manufacturer. The purified products were sequenced in both strands on the ABI Prism 3700 DNA sequencer (Perkin-Elmer). The obtained sequences were compared to that of pbp5 included in GenBank accession no. X84860.
The detection of genes encoding the virulence factors, including enterococcal surface protein (esp) and hyaluronidase (hyl), was determined in all Enterococcus spp. isolates using PCR. 13 The positive and negative controls (from the strain collection of University of Trás-os-Montes and Alto Douro, Portugal) were used.
Multilocus sequence typing
vanA-containing E. faecium isolates were characterized by multilocus sequence typing (MLST) as previously described following the guidelines published in the website www.mlst.net. 14 The produced sequences were studied to determine the corresponding sequence type (ST) and clonal complex (CC) at the (http://mlst.ucc.ie) URL.
Results
From the 305 fecal samples, six E. faecium isolates were detected, all of them being resistant to vancomycin (MIC >128 mg/l) and teicoplanin (MIC 64 mg/l) and carrying the vanA gene (Table 1).
AMP, ampicillin; CC, clonal complex; CIP, ciprofloxacin; ERY, erythromycin; GEN, gentamycin; KAN, kanamycin; MIC, minimal inhibitory concentrations; MLST, multilocus sequence typing; QD, quinupristin–dalfopristin; ST, sequence type; STR, streptomycin; TEI, teicoplanin; TET, tetracycline; VAN, vancomycin.
Phenotypic resistance to chloramphenicol was not identified in this study. Resistance to ampicillin was identified in five isolates; four isolates showed high-level resistance for aminoglycosides; four to tetracycline and ciprofloxacin, and only one showed quinupristin–dalfopristin resistance.
All the isolates were resistant to erythromycin and harbored the erm(B) gene. The tet(M) gene was identified in all of the tetracycline-resistant isolates. The vat(E) was detected in the quinupristin–dalfopristin-resistant isolate. The aminoglycoside resistance genes acc(6′)-Ie-aph(2″)-Ia, aph(3′)-IIIa, and ant(6′)-Ia were found in the four isolates showing high-level resistance for aminoglycosides. None of the VRE isolates was found to carry the van(B), van(C), tet(L), erm(A), erm(C), vat(D), or aad(E) genes.
The C-terminal region of the pbp5 gene was sequenced in ampicillin-resistant isolates (MIC range of 256 μg/ml) and it was compared to the pbp5 reference sequence (GenBank X84860). Two different pbp5 alleles were detected when considering the changes of amino acid in 461–629 region (A and B). Important amino acid changes (485M→A, 496N→K, 499A→T, 525E→D, 629E→V) were detected in the alleles and also a serine insertion at 466′ position. The alleles also showed amino acid changes in 499Q→K and 470H→Q. A correlation was detected between ampicillin MICs and the deduced amino acid changes in pbp5 (Table 2).
Isolates from red-legged partridges.
The allele designation (A and B) has been designed in this study.
The important amino acid positions are indicated in bold. In addition, amino acid changes at these positions with respect to the reference sequence X84860 are also indicated in bold.
The sequence included in GenBank with the accession number X84860 was used as reference for comparison purposes.
Genes encoding virulence factors were studied in all E. faecium isolates. The combination of hyl+esp genes was detected in five isolates, whereas the esp gene alone was found in only one isolate.
The STs detected with the MLST analysis of the six vanA-E. faecium isolates revealed that four isolates belonged to ST18 (E. faecium CC17), one isolate belonged to ST 448 (E. faecium CC17), and the last one belonged to ST139 (E. faecium CC5). Using eBURST, it was possible to compare the relationships between the obtained STs, as shown in Fig. 1.
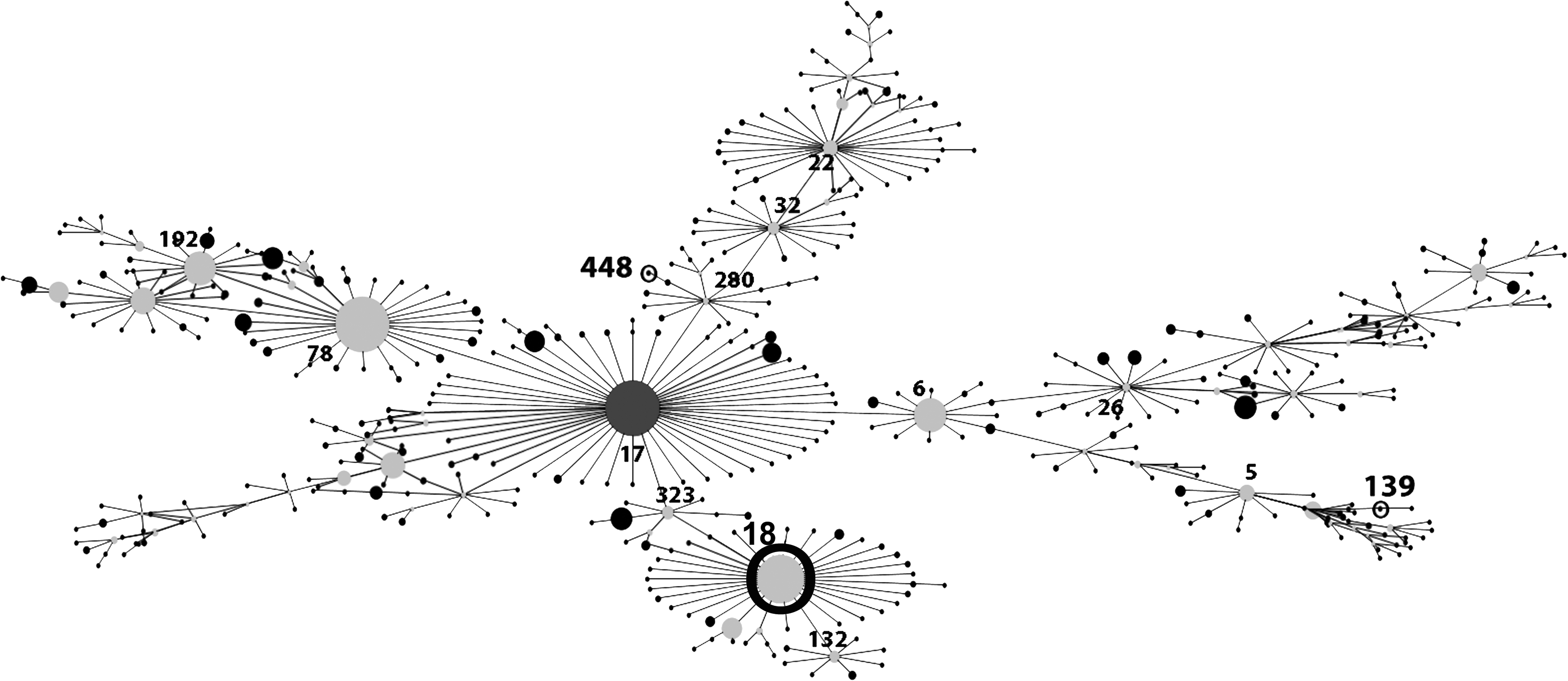
Clustering of Enterococcus faecium STs from fecal samples of red-legged partridges from Portugal. Each ST is represented as a node; lines connect single-locus variants. The size of each node is directly proportional to the prevalence of ST in the input data. STs found in this study are numbered in bold and circled. The complex of related STs and subsequent patterns of evolutionary descent between ST18, ST448, and ST139ST is represented in this figure. ST, sequence type.
Discussion
As far as we know, this is the first report of vanA-E. faecium strains in red-legged partridges (A. rufa) in Portugal. Several studies of antibiotic resistance in wild birds have been conducted.9,15–17 E. faecium harboring the vanA gene has already been found in wild birds in Portugal.12,15–17,18
All isolates of E. faecium showed multiresistance since they were resistant to almost all the antibiotics used in this study. Resistance to vancomycin and erythromycin was identified in all isolates. Similar to our findings, another study with red-legged partridges conducted in Spain demonstrated that erm(B) was the most commonly found erythromycin resistance gene in this species. 19 The resistance genes erm(A) and erm(C) were not detected in the isolates tested. Although the tetracycline-resistant isolates harbored the tet(M) gene, the tet(L) gene was not detected. The tet(M)gene is the most widespread gene among the tetracycline resistance genes.19,20 The vanA, erm(B), and tet(M) were the antibiotic resistance genes most detected in our study. The vanA gene cluster is located in the transposable element Tn1546, frequently integrated into conjugative plasmids, which often contains other antibiotic resistance genes.19,20
Phenotypic resistance to ampicillin was identified in five (83.3%) of the six isolates. Many studies found E. faecium resistant to ampicillin due to modifications in the penicillin-binding proteins (pbp5) leading to acquisition of resistance, however, this type of resistance is more frequently found in humans than animals.12,21 The five AMPR isolates showed changes at six positions when considering only the principal amino acid positions (466′, 485, 496, 499, 525, 586, and 629) of pbp5. Several studies have detected insertions of aspartic acid at 466′ position in strains that presented a high level of ampicillin resistance.12,22 In isolates with higher MICs (256 μg/ml), the conversion of methionine to alanine was detected at position 485. 23 The 629E→V substitution may be responsible for a decrease of the protein capacity to bind to β-lactam, since the residue in position 629 is near to the threshold of the active site. 24
Only one of our isolates showed quinupristin–dalfopristin resistance with the presence of vat(E) gene. The vat(D) gene was not detected. The vat(E) gene is often detected in vanA-containing E. faecium isolates from humans and animals. 20
Gentamycin resistance was identified in one isolate that harbored the acc(6′)-Ie-aph(2″)-Ia. The bifunctional enzyme acc(6′)-Ie-aph(2″)-Ia is responsible for gentamycin resistance. This enzyme also confers resistance to almost all aminoglycosides, including kanamycin. 25 Kanamycin resistance was found in three more isolates that harbored the aph(3′)-IIIa gene. Phenotypic resistance to streptomycin was found in only one isolate that contained the ant(6′)-Ia gene.
The mechanisms of human infections caused by enterococcal virulence genes are not quite understood yet. Virulent strains can colonize the intestinal tract and spread to other organs and body sites, such as the urinary tract. The presence of E. faecium alone does not mean the beginning of an infection since the presence of other mediators may be necessary and has not been completely characterized.26,27 The hyl gene was found in five (83.3%) of the six isolates and the esp gene was detected in all of them. These two genes are the most frequently found in hospital outbreaks caused by VRE strains and are usually associated with ampicillin and ciprofloxacin resistance. 28 Furthermore, in this study, almost all of the isolates showed ampicillin resistance.
The characterization by MLST grouped the isolates as ST18 (4), ST448 (1), and ST139 (1). The ST18 is a double-locus variant of ST17 belonging to the HiRCC CC17 and has already been isolated from wild birds. 15 This CC has been associated with E. faecium isolates that are responsible for many hospital outbreaks and it is known that they are able to develop adaptation to the hospital environments. 29 Studies demonstrated that ST18 often carries the esp gene, the vanA gene, and resistance to ampicillin (MLST database http://efaecium.mlst.net). 30 The ST448 is a single-locus variant of the ST750 found for the first time in American crows, which, in turn, is indirectly linked to ST17 and belongs to CC17. 14 The ST448 was previously found in crows and humans, namely in a hospitalized patient in Brazil.31,32 The ST139 is a triple-locus variant of the ST5 and belongs to CC5. E. faecium CC5 are commonly found in pigs in Europe and several studies have found swine-adapted E. faecium CC5 strains in humans.4,33 Wild partridges are a typical farmland animal that fed themselves mainly on insects and seeds. As omnivorous birds living in closeness to human activity, they could be colonized or may be transient carriers of clinically significant VRE originating from healthcare facilities. Sometimes, sludge from wastewater treatment plants from hospitals is used on agricultural lands where partridges feed. The untreated sewage outflow used in the agricultural fields and the livestock animals may also be a source of contamination that could explain the presence of VRE strains as well as the STs detected.
Based on the results obtained in this study, the red-legged partridges may be acting as reservoirs and donors of antibiotic and virulence determinants. The sources for colonization of wild partridges can be several, from farms to agricultural fields to hospitals. Since the partridge is a wild game bird, the ingestion of the contaminated meat by humans is a concern. Future studies should be carried out for a better understanding of the dissemination of VRE strains in wild ecosystems and their risk to human and animal health.
Footnotes
Disclosure Statement
No competing financial interests exist.