
Abstract
A total number of 368 clinical isolates of Streptococcus agalactiae (group B Streptococcus, GBS) were collected in 2010–2016 from three hospitals in a region of central Italy. Fluoroquinolone (FQ)-resistant isolates were selected using levofloxacin. Levofloxacin-resistant (LR) strains (11/368, 2.99%) were characterized for several features, and their FQ resistance was analyzed phenotypically and genotypically using seven additional FQs. Their gyrA and parC quinolone resistance-determining regions were sequenced. Of the 11 LR isolates, 10 showed high-level and 1 low-level resistance. The former isolates exhibited higher minimal inhibitory concentrations also of the other FQs and all shared one amino acid substitution in ParC (Ser79Phe) and one in GyrA (Ser81Leu); only Ser79Phe in ParC was detected in the low-level LR isolate. The 11 LR strains exhibited distinctive relationships between their susceptibilities to non-FQ antibiotics and typing data. Remarkably, despite the very rare occurrence of chloramphenicol resistance in S. agalactiae, no <4 of the 11 LR isolates were chloramphenicol-resistant. Studies of GBS resistance to FQs in Europe remain scarce, notwithstanding the emergence of multidrug-resistant isolates. The incidence of LR GBS isolates is still limited in Italy, consistent with the moderate (though growing) rates reported in Europe, and much lower than the very high rates reported in East Asia. The intriguing relationships between FQ and chloramphenicol resistance deserve further investigation.
Introduction
B
Resistance to FQs primarily results from alterations in their target enzymes DNA gyrase and topoisomerase IV: both are involved in DNA replication and are heterotetrameric, being composed of two pairs of subunits, GyrA and GyrB (respectively encoded by the gyrA and gyrB genes) in DNA gyrase, and ParC and ParE (respectively encoded by parC and parE) in topoisomerase IV. FQ resistance can also be due to an efflux mechanism mediated by overexpression of multidrug resistance efflux pumps. In itself, efflux is normally unable to cause high-level resistance, but may facilitate mutational resistance by allowing short-term bacterial survival. Especially in Gram-positives—where the vast majority of studies of streptococci have focused on Streptococcus pneumoniae 13 —resistance typically develops in a stepwise fashion, due to amino acid changes occurring particularly in regions of the GyrA and/or ParC subunits known as the quinolone resistance-determining regions (QRDRs). 14 Highly FQ-resistant isolates detected in Streptococcus species of the viridans group (VGS), 15 in S. pyogenes 9 and in S. agalactiae8,16 displayed a mode of resistance comparable to the one extensively investigated in S. pneumoniae.
In this study, we explored the issue of FQ resistance in a collection of S. agalactiae clinical isolates recovered over the past 7 years from three hospitals of central Italy. The levofloxacin-resistant (LR) isolates were selected, characterized for several features, and their FQ resistance was analyzed phenotypically and genotypically using seven additional FQs.
Materials and Methods
Bacterial strains
We started from a collection of 368 nonduplicate isolates of S. agalactiae. Of them, 302 were nonsystematically collected in 2010–2015 from the clinical laboratories of three hospitals (Ancona Torrette, Jesi, and Fano) of a region in central Italy (Marche), whereas 66 were systematically collected in 2016 from Ancona Torrette hospital. Isolates were from pregnant women (genital carriage or infections), neonatal and pediatric infections, urine, and adult infections.
The 11 isolates selected for levofloxacin resistance (see Selection of LR strains under Results section) were confirmed as being Lancefield group B using a latex agglutination test (Slidex Strepto Plus; bioMérieux, Marcy-l'Étoile, France) and were designated SagLR1 to SagLR11 according to the chronological order of their isolation, irrespective of the hospital of origin.
Antibiotics and susceptibility tests
With the exception of ulifloxacin—the active metabolite of prulifloxacin, which was obtained from Angelini ACRAF, Pomezia, Italy—all other antibiotics were purchased from Sigma-Aldrich (Milan, Italy). Minimal inhibitory concentrations (MICs) were determined by standard broth microdilution. 17 The MICs of norfloxacin, ciprofloxacin, levofloxacin, and ulifloxacin were also determined in the presence of 10 μg/ml of reserpine (Sigma-Aldrich), to detect possible MIC reductions and estimate the contribution of efflux to FQ resistance. 14
The MLS phenotype was determined and categorized—as constitutive (cMLS), inducible (iMLS), or M—by the conventional erythromycin-clindamycin double-disk agar diffusion test. 17
Analysis of FQ resistance
The QRDRs of the gyrA and parC genes were investigated by PCR followed by restriction fragment length polymorphism (RFLP) according to a procedure well-established for S. pneumoniae, 18 using primer pairs specifically designed for GBS. 9 S. agalactiae 2603V/R (ATCC BAA-611, accession no. AE009948) was used as a FQ-susceptible reference strain for comparative analysis. For RFLP analysis, the amplicons obtained from gyrA and parC were digested with HinfI (Roche Applied Science, Basel, Switzerland) and electrophoresed in 2% agarose. The procedure allowed preliminary screening of the most common QRDR codons undergoing mutation in both gyrA (position 81) and parC (position 79). The amplicons were then sequenced and analyzed.
DNA sequencing and sequence analysis
DNA sequencing and sequence analysis were performed as described elsewhere. 19
Detection of resistance genes
A number of erythromycin, tetracycline, and chloramphenicol resistance genes were searched for by PCR assays as described elsewhere. 20 The erythromycin resistance genes included erm(B), erm(TR), erm(T), and mef, the mef subclass being identified as described previously 21 ; the tetracycline resistance genes included tet(M) and tet(O); and the chloramphenicol resistance genes included catpC194, catpC221, and catQ.
Typing
Macrorestriction with SmaI endonuclease (Roche Applied Science) and pulsed-field gel electrophoresis (PFGE) analysis of LR GBS isolates were carried out and pulsotypes were distinguished as described elsewhere. 22 A dendrogram was constructed using the Dice coefficient and the unweighted pair group method with arithmetic averages. Pulsotypes were clustered on the basis of a cutoff of 70% similarity. Capsular types were determined by a multiplex PCR assay. 23 Multilocus sequence typing (MLST) was performed using specific primers as described previously 24 and allelic profile assignment and sequence type determinations were made using the S. agalactiae MLST website (http://pubmlst.org/sagalactiae). Surface protein markers were inferred by using a multiplex PCR for the direct identification of the Alpha-like protein (Alp) family genes (alpha C, rib, alp2/3, alp4, and epsilon). 25
Results
Selection of LR strains
The whole GBS collection was selected for resistance to levofloxacin (MIC, >2 μg/ml). 17 Resistance was detected in 11 strains (2.99%), 9 from women and 2 from men, 6 isolated in 2010–2015 and 5 in 2016. Overall, 10 strains exhibited high-level levofloxacin resistance (MIC, ≥32 μg/ml) and 1 (SagLR4) exhibited low-level resistance (MIC, 4 μg/ml). SagLR4 was confirmed as nonsusceptible by the norfloxacin disk diffusion screen test. 17
Early characterization of LR strains
The main data from the early characterization of the 11 LR strains and the isolation and demographic data are summarized in Table 1. PFGE typing—performed only in the 10 highly LR isolates since SagLR4 was resistant to SmaI digestion—revealed seven different pulsotypes (1, 2a, 2b, 2c, 2d, 3, and 4). The resulting dendrogram is shown in Figure 1.
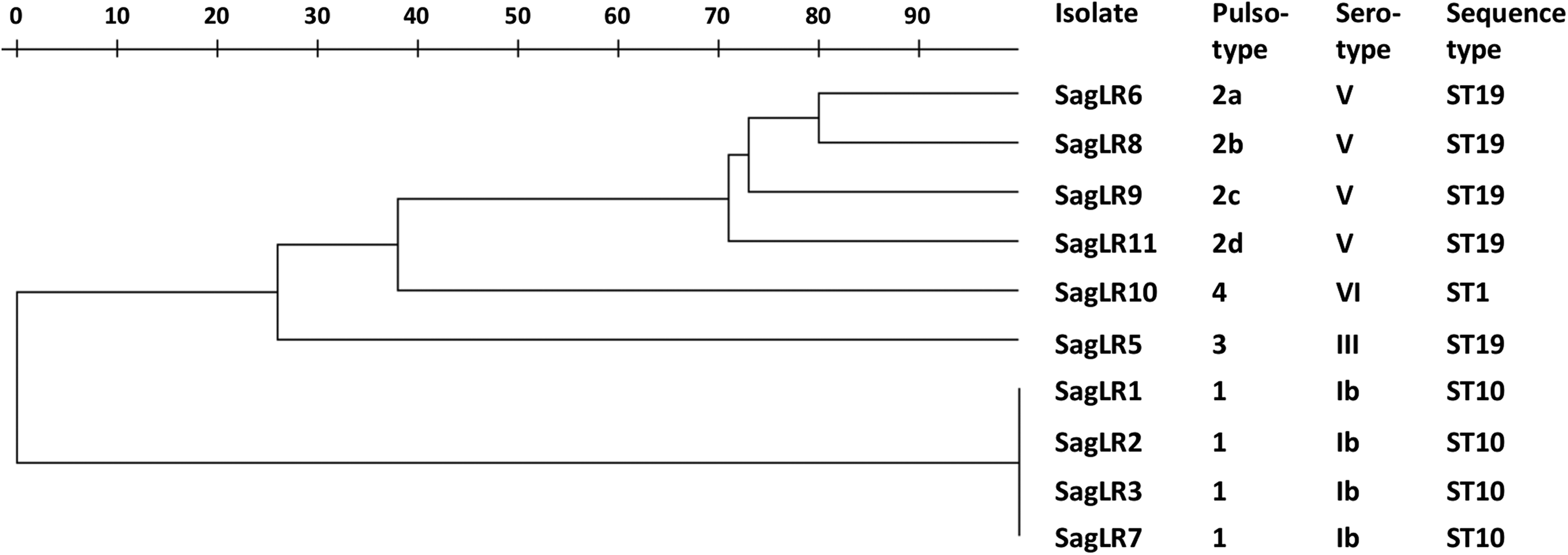
Dendrogram of the 11 levofloxacin-resistant group B Streptococcus isolates.
CHL, chloramphenicol; ERY, erythromycin; TET, tetracycline; ut, untypeable (due to resistance to SmaI digestion); Alp, Alpha-like protein; ST, sequence type. Hospitals: H1, Ancona Torrette; H2, Jesi; H3, Fano.
The 11 LR isolates were all susceptible to penicillin, vancomycin, and linezolid, and exhibited particular relationships between their susceptibilities to the other drugs and their typing data:
• Four clonally related strains (SagLR1, SagLR2, SagLR3, and SagLR7, the first three being the oldest isolates) were susceptible to all drugs and shared serotype Ib, the Alp gene alpha C, pulsotype 1, and ST10. • Another four strains (SagLR6, SagLR8, SagLR9, and SagLR11, the last three being 2016 isolates) shared serotype V, the Alp gene epsilon, ST19, and related PFGE profiles. All were multidrug-resistant: in addition to FQs, all were resistant to tetracycline [due to tet(M)] and erythromycin, three with the iMLS phenotype [one due to erm(TR), two due to erm(TR)+mef(I)] and one, the sole highly erythromycin-resistant, with the cMLS phenotype [due to erm(B)]. Three of these four isolates were also resistant to chloramphenicol [two due to catQ, and one—SagLR9, the one carrying erm(B)—due to catpC194]. Remarkably, in SagLR9 the catpC194 determinant was located—linked to and about 6 kb apart from erm(B)—in a chromosomal region whose characterization is in progress. • The remaining three strains (SagLR4, SagLR5, and SagLR10) were more heterogeneous, sharing only resistance to tetracycline [due to tet(M)], in addition to FQs, and the Alp gene rib. SagLR4, the only LR isolate with low-level resistance, showed no other antibiotic resistances and displayed serotype III and ST17, but was PFGE untypeable due to its resistance to SmaI digestion. SagLR5 was also resistant to erythromycin [due to erm(TR)+mef(I)] and chloramphenicol (due to catQ) and displayed serotype III, a distinctive pulsotype (3), and ST19. Finally, SagLR10 was also resistant to erythromycin [due to erm(TR)] and displayed serotype VI, a distinctive pulsotype (4), and ST1.
FQ susceptibility profiles
The FQ MICs determined in the 11 LR isolates are reported in Table 2, together with those of the FQ-susceptible reference GBS 2603V/R. SagLR4, the lower-level LR isolate, had lower MICs also of the other FQs: norfloxacin MIC, 32 μg/ml; other FQ MICs, 0.5–4 μg/ml, the 0.5 μg/ml value of moxifloxacin actually falling in the susceptibility range. 17 The other 10 LR isolates exhibited consistently higher MICs not only of levofloxacin (32–64 μg/ml), but also of the other FQs. The addition of reserpine did not change the MIC values of norfloxacin, ciprofloxacin, levofloxacin, or ulifloxacin.
Fluoroquinolone-susceptible reference strain 2603V/R is shown for comparison at the bottom of the table.
CIP, ciprofloxacin; LVX, levofloxacin; MICs, minimal inhibitory concentration values; MXF, moxifloxacin; NOR, norfloxacin; OFX, ofloxacin; SPX, sparfloxacin; TVA, trovafloxacin; ULX, ulifloxacin; wt, wild type.
Analysis of the QRDRs of gyrA and parC
The QRDRs of gyrA and parC were sequenced in all the 11 LR GBS isolates. Amino acid changes resulting from base pair substitutions were determined by comparing the sequences with those of reference strain 2603V/R. All LR isolates displayed Ser79Phe (codon TCC→TTC) in ParC, and all except SagLR4 displayed Ser81Leu (codon TCA→TTA) in GyrA. Such amino acid substitutions have previously been described as contributing to FQ resistance not only in S. agalactiae,8,11,12,16 but also in S. pyogenes, 26 S. pneumoniae, 27 and Streptococcus dysgalactiae subspecies equisimilis. 9
Discussion
The selection of LR GBS strains (11 in a collection of 368 isolates, 2.99%) deserves special attention because it yielded a number of unexpected, intriguing findings that primarily involved chloramphenicol resistance:
• Considering the very rare occurrence of chloramphenicol resistance in S. agalactiae,9,28–31 it was astonishing that no <4 of the 11 isolates selected for levofloxacin resistance were chloramphenicol-resistant, resistance being mediated in three isolates by catQ and in one by catpC194. These four isolates were the sole chloramphenicol-resistant strains in the collection; in other words, no chloramphenicol resistance was detected in the levofloxacin-susceptible isolates. • Interestingly, the older two (SagLR5 and SagLR6) of these four isolates were the protagonists—under the names of Sag236 and Sag403—of a previous study where the GBS collection had been selected for chloramphenicol resistance.
20
The two strains were found to harbor an apparently identical catQ-, mef(I)-, and erm(TR)-carrying mosaic integrative and conjugative element (ICESag236), which appeared to be transferable, at low frequency, only from the former strain. Further differences between the two isolates were recorded—and have now been confirmed and extended—in typing data: of the typing characters tested, they only shared the sequence type (ST19). • The third catQ-positive strain, SagLR8—which had not yet been recovered when ICESag236 was detected in SagLR5 and SagLR6—also harbors an ICESag236-like element, as shown by PCR mapping. It also shares ST19, whereas other typing features resemble those of SagLR6 but differ clearly from those of SagLR5. • catQ is a unique cat determinant originally detected in Clostridium perfringens.
32
Recently, however, it has been detected in several Streptococcus species, where it is typically linked to the macrolide efflux mef gene subclass mef(I)—or also mef(E) in VGS—to form the so-called IQ module.19–21,33–35 In a surveillance study of invasive S. agalactiae recovered during 2015 in the United States, chloramphenicol resistance was detected in only 2 out of 1975 isolates (0.1%): interestingly, both carried catQ.
31
• catpC194, harbored by SagLR9, was originally detected and sequenced in the Staphylococcus aureus plasmid pC194
36
; subsequently, it was largely described in S. pneumoniae in the same plasmid, integrated in linearized form in Tn5253.
37
In S. agalactiae, catpC194 appears to be very uncommon: in silico analysis identifies the gene in only one out of over 800 deposited genomes of this species. A completely different situation is found for SagLR9 where, according to an investigation currently in progress, catpC194 is not part of plasmid pC194.
The most interesting findings of the analysis of FQ resistance in the 11 LR GBS strains concerned SagLR4, the lower-level LR isolate. The other 10 isolates only shared fairly expectable features such as high-level resistance to all FQs tested and the two most common substitutions (Ser81Leu in GyrA and Ser79Phe in ParC) detected in FQ-resistant S. agalactiae, beginning from the first described highly resistant isolate of this species. 8 More interestingly, SagLR4 displayed (1) trovafloxacin and moxifloxacin MIC values (0.5 μg/ml) similar to those of the FQ-susceptible reference strain 2603 V/R; (2) a high MIC only of norfloxacin (32 μg/ml, just one dilution lower than for the other LR isolates); and (3) moderately increased MICs (no higher than 4 μg/ml) of the other FQs. No MIC decrease in the presence of reserpine being observed, the lower FQ MICs displayed by SagLR4 apparently depended on the sole amino acid substitution detected, Ser79Phe in ParC. On the other hand, in the stepwise progression of FQ resistance—well established chiefly in S. pneumoniae 13 —a first-step mutation in parC associated with low-level resistance normally precedes, and makes more likely, a second mutation in gyrA resulting in high-level FQ resistance. The second amino acid substitution (Ser81Leu) in GyrA in the 10 highly FQ-resistant isolates is consistent with the hypothesis that a similar stepwise progression of FQ resistance also applies to S. agalactiae.
An increase in LR GBS appears to have occurred recently in our region: of the 11 LR GBS isolates we collected starting from 2010, 5 were isolated in 2016. Moreover, the incidence of levofloxacin resistance recorded in this study (2.99%) is more than twice, despite different procedures followed for MIC testing, the one (1.4%) reported in the country in clinical GBS isolates collected in 2013–2014. 11 However, it is still a limited rate, consistent with the moderate, though growing, rates reported in Europe,9,12 far distant from the very high rates reported in some East Asian countries.7,38–41 The finding of FQ-resistant GBS may also vary with the patients and specimens examined: in particular, such isolates appear to be rarer in GBS from neonatal infections than from other sources. 12 Interestingly, none of our LR GBS isolates were from neonates.
The literature is not univocal on whether the development of FQ resistance in GBS is the prerogative of a few clonal strains16,38 or else it occurs in unrelated strains. 10 Our results indicate that only 4 of our 11 LR GBS isolates appeared clearly to belong to a clone, whereas clonal relationships were not as evident in the other seven isolates. On the other hand, some heterogeneity in LR GBS isolates sharing ST19 (which include the four chloramphenicol-resistant isolates) might depend on recent recombination events such as the acquisition of genetic elements carrying antibiotic resistance genes: a similar mechanism may have contributed to the genome plasticity and the expansion of the CC17 lineage described in Chinese GBS isolates. 42
Unfortunately, studies of GBS resistance to FQs in Europe remain scarce, notwithstanding the emergence of multidrug-resistant isolates. 12 Yet, the issue deserves greater attention, with follow-up studies addressing, in particular, the relationships between FQ and chloramphenicol resistance.
Moreover, since quinolones are not used to treat infections in pregnant women and children, the relevant GBS isolates are not usually tested for quinolones in the routine laboratory workup: it would therefore be important—at least for surveillance purposes—to modify such a practice by extending FQ susceptibility testing to all clinical GBS isolates.
Footnotes
Acknowledgment
This study was supported by internal funding.
Disclosure Statement
No competing financial interests exist.