
Abstract
Aim:
We investigated the prevalence of extended-spectrum β-lactamase (ESBL)-producing Enterobacteriaceae among wild fish from the coast of Bejaia (Algeria) in the Mediterranean Sea.
Results:
From March 2012 to August 2013, gut and gill samples of wild fish were screened for the presence of ESBL-producing Enterobacteriaceae. Strains were characterized with regard to antibiotic resistance, β-lactamase content, plasmid-mediated quinolone resistance, aminoglycoside resistance genes, and clonality (repetitive sequence-based polymerase chain reaction profiles and multilocus sequence typing). Virulence traits were performed for Escherichia coli and Klebsiella pneumoniae isolates. Of the 300 fish studied, 64 (21.3%) isolates were screened as positive for ESBL producing by the double-disc method. The isolates corresponded to E. coli, K. pneumoniae, Enterobacter cloacae, Morganella morganii, Citrobacter freundii, and Proteus vulgaris. A predominance of blaCTX-M gene was observed with a prevalence of 60.5% (n = 46). Furthermore, our study describes the association of important coresistance and virulence factors in E. coli and K. pneumoniae. Twelve of the ESBL producers carried genes of the qnr family and oqxAB gene and six carried the aac(6′)-Ib-cr gene.
Conclusions:
Our results highlight for the first time the diffusion of multidrug-resistant Enterobacteriaceae isolates carrying resistance and virulence genes in fish from the Mediterranean Sea in Algeria.
Introduction
A
The emergence of multidrug resistance (MDR) in bacteria from different habitats has been rising at a disquieting rate and is becoming a severe problem throughout the world.5,6 Horizontal gene transfer represents one of the main causes of the rapid proliferation of antibiotic resistance genes across a wide diversity of bacteria. In this way, the integrons are genetic elements that acquire, exchange, and express antibiotic resistance genes. They are widely distributed, especially in Gram-negative bacteria, and are involved in the antibiotic resistance spread in the environment (agriculture, industrial effluent, wastewater treatment plant, etc). 6 The contamination of different environmental niches in several regions and the role of integrons explain the low success of the MDR bacteria containment measures.6,7
There is growing evidence that Enterobacteriaceae harboring clinically relevant bla genes, especially genes encoding the CTX-M family of ESBLs, have spread into the aquatic environment. 8 Moreover, it has been suggested that one of the main routes for transfer of resistance bacteria and resistance genes from animals and plants to human is via the food chain. 9
Fish living in the natural environment are known to harbor pathogenic Enterobacteriaceae.10–12 Hence, fish are considered a potential vehicle of foodborne bacterial infections, which might be of importance in human public health. Contamination of fish with MDR bacteria could demonstrate the risk of the persistence of these bacteria in the fish gut flora and explain the possible human gut contamination.13,14
However, few studies have investigated ESBL-producing Enterobacteriaceae in wild fish. Contamination of surface waters in Algeria is of a great concern. Treated and untreated wastewaters have been discharged into rivers and streams, leading to possible waterborne infection outbreaks, which may represent a significant dissemination mechanism of antibiotic resistance genes among pathogenic bacterial populations. The purpose of this study was to investigate the occurrence of resistance genes in Enterobacteriaceae isolates recovered from wild fish from the coast of Bejaia (Algeria) in the Mediterranean Sea, to describe the clonal diversity of the ESBL isolates, and to characterize the content of virulence genes in these isolates.
Materials and Methods
Bacterial isolates
Enterobacteriaceae isolates were collected from different fish samples caught in the Mediterranean Sea near Bejaia (Algeria). A total of 300 fish samples, including 62 samples of Sardina pilchardus, 60 samples of Sarpa salpa, 55 samples of Pagellus acarne, 45 samples of Trachurus trachurus, 40 samples of Boops boops, and 38 samples of Engraulis encrasicolus, were collected from March 2012 to August 2013. All the fish were sampled by opening the gut or the gills using a sterile scalpel following washing the surface with sterile saline, sampling was carried out from gut and gills of each fish, and samples (∼10 g) were placed in 1 ml sterile 0.9% saline and then vortexed. To screen for the presence of ESBL-producing Enterobacteriaceae isolates, 100 μl of the suspension was streaked on two MacConkey agar plates supplemented either with cefotaxime (2 μg/ml) or ceftazidime (2 μg/ml) and incubated for 24 hours at 37°C under aerobic conditions. A single presumptive Enterobacteriaceae colony per plate was selected and identified using the matrix-assisted laser desorption and ionization time-of-flight mass spectrometry method (VITEK MS; bioMérieux). Klebsiella pneumoniae identification was confirmed by VITEK 2 identification card (bioMérieux).
Antimicrobial susceptibility testing
The antimicrobial susceptibility of bacterial isolates was determined using the agar diffusion method according to the CA-SFM/EUCAST recommendations (www.sfm-microbiologie.org). The following antimicrobials were tested: ceftazidime (CAZ: 10 μg), cefotaxime (CTX: 5 μg), cefepime (FEP: 30 μg), ertapenem (ERT: 10 μg), imipenem (IMP: 10 μg), gentamicin (GM: 10 μg), amikacin (AMK: 30 μg), netilmicin (NET: 10 μg), nalidixic acid (NAL: 30 μg), ofloxacin (OFX: 5 μg), ciprofloxacin (CIP: 5 μg), and cotrimoxazole (SXT: 1.35/23.75 μg). Minimal inhibitory concentration of colistin (0.5–128 mg/L) was determined using microbroth dilution (Umic®; Biocentric). Isolates were screened for ESBL production by the standard double-disc synergy test, as described previously. 15 Isolates resistant to ≥3 antimicrobial classes were considered as MDR. 16
Detection of antibiotic resistance gene
Bacterial DNA was extracted from the isolates by the EZ1 DNA Tissue kit following the manufacturer's instructions on the BioRobot EZ1 extraction platform (QIAGEN, Courtaboeuf, France). The genes coding for ESBL (blaTEM, blaSHV, blaCTX-M) and plasmid-mediated AmpC-type were detected by polymerase chain reaction (PCR) using specific primers as previously described17–20 followed by sequencing of the PCR products.
The isolates were investigated for the presence of the PMQR determinants qnrA, qnrB, qnrC, qnrD, qnrS, aac(6′)-Ib-cr, qepA, and oqxAB genes as previously published21–26 followed by sequencing of the PCR products. The aminoglycoside resistance genes aac(3′)-IV, aac(3′)-II, aac(3)-III, ant(2")-I, and aac(3′)-I were screened by PCR as previously described. 27 The sulfonamide resistance genes (sul1, sul2, sul3) were detected by PCR as previously described. 28
Phylogenetic grouping of E. coli
The E. coli isolates were assigned to the phylogenetic groups A, B1, B2, D, E, and F using a multiplex PCR as previously described. 29
Clonality study
The genetic relationship of the strains was studied using the DiversiLab system (bioMérieux), a semiautomated fingerprinting tool based on repetitive sequence-based PCR (rep-PCR), following the manufacturer's instructions. Isolates with identical strain patterns were considered indistinguishable if their similarity percentage was ≥95%.
Multilocus sequence typing (MLST) was carried out on E. coli and K. pneumoniae strains according to the Institut Pasteur's MLST scheme (bigsdb.web.pasteur.fr).
Detection of E. coli virulence-associated genes
The in vitro virulence potential of E. coli strains was evaluated by PCR30–32 with the following genes: papAH, papC, papEF, papG alleles I, II, III (encoding P fimbriae); sfaS and focG (S fimbriae and F1C fimbriae); afa/draBC (Dr family adhesin); fimH (mannose-specific adhesin subunit of type 1 fimbriae); hlyA (hemolysin); cnf1 (cytotoxic necrotizing factor-1); iutA (aerobactin siderophore receptor); kpsMT II and kpsMK I (capsule synthesis); traT (serum resistance-associated factor); malX (pathogenicity island marker from strain CFT073); iroN (iron siderophore receptor); and irp2 (yersiniabactin). Extraintestinal pathogenic E. coli (ExPEC) status (i.e., exhibition of ≥2 virulence factor (VF) genes among the following VFs: pap, sfa/focDE, afa/draBC, iutA, and kpsMT II) was determined as previously described. 31
Detection of Klebsiella virulence-associated genes and serotype
The in vitro virulence potential of K. pneumoniae strains was also tested by multiplex PCR with the following genes: wzy_K1 (capsular serotype K1 and hypermucoviscosity phenotype); rmpA (regulator of mucoid phenotype A); entB (siderophore); ybtS (yersiniabactin); kfu (iron transport and phosphotransferase function); iutA (hydroxamate siderophore); mrkD (adhesin type 3 fimbriae); allS (associated with allantoin metabolism); and the K2 capsular serotype genes, 33 cf29a, fimH, uge, wabG, ureA genes, and the genes of the capsular serotypes K5, K57, K54, and K20 were detected as previously described.33–35
Results
Bacterial collection
Of 300 fish samples collected, 64 (21.3%) were screened as positive for ESBL-producing Enterobacteriaceae by the double-disc method. Species identified were E. coli (22, 34.3%), K. pneumoniae (11, 17.2%), Enterobacter cloacae (10, 15.6%), Morganella morganii (9, 14.1%), Citrobacter freundii (8, 12.5%), and Proteus vulgaris (4, 6.3%).
The β-lactam resistance was as follows: 100% of isolates were resistant to cefotaxime, and 61 isolates (95.3%) were resistant to cefepime. All the isolates were susceptible to carbapenems and colistin. The other resistance profiles showed that resistance to nalidixic acid and ofloxacin was the most frequent (23 [35.9%] and 21 [32.8%] isolates, respectively), followed by resistance to ciprofloxacin (19 isolates, 29.7%), cotrimoxazole (15 isolates, 23.4%), and gentamicin (9 isolates, 14.1%) (Table 1). Interestingly, MDR was exclusively found in E. coli (n = 12) and Klebsiella spp. (n = 3). The other species were susceptible to aminoglycosides, cotrimoxazole, and quinolones (Table 1).
AMK, amikacin; CAZ, ceftazidime; CIP, ciprofloxacin; COL, colistin; CTX, cefotaxime; ETP, ertapenem; FEP, cefepime; GM, gentamicin; IMP, imipenem; NA, nalidixic acid; NET, netilmicin; OFX, ofloxacin; SXT, cotrimoxazole.
Characterization of ESBLs and ampC genes
The overall prevalence of ESBL was 21.3% (n = 64), with the prevalence of CTX-M, TEM, and SHV producers as 68.8% (n = 44), 23.4% (n = 15), and 7.8% (n = 5), respectively. The distribution of the different types of ESBLs in the different species recovered during this study is shown in Table 2.
ESBL, extended-spectrum β-lactamase.
Among the CTX-M producers, 88.6% produced CTX-M-15 β-lactamases. The other CTX-M producers were the CTX-M-8 group (9.1%, 4/44) and CTX-M-9 group (2.3%, 1/44). TEM and SHV producers were TEM-24 and SHV-5, respectively.
The prevalence of plasmidic ampC genes was 11.5% (n = 7). All were detected in C. freundii isolates, which harbored dha-1 gene.
Detection of PMQR and aminoglycoside genes
Among the quinolone-resistant strains, the prevalence of qnr was 17.2% (n = 11) (Table 2). Three qnrB2 were identified in two E. coli and one K. pneumoniae; all these isolates produced CTX-M-15 β-lactamases. Eight were qnrS1 + from six E. coli and two K. pneumoniae; six harbored blaCTX-M-15 genes and two blaTEM-24.
PCR and DNA sequencing results showed that the prevalence of aac(6′)-Ib-cr was 9.4% (n = 6) (Table 2). The resistance gene was identified in five E. coli and one K. pneumoniae; five producing CTX-M-15 β-lactamases. The prevalence of oqxAB was 17.2% (n = 11). All the K. pneumoniae isolates harbored this gene. Finally, we noted an association between qnrS1, aac6′-Ib-cr, and oqxAB in one CTX-M-15-producing K. pneumoniae isolate.
Resistance to aminoglycosides was associated with the presence of aac(3′)-IV (n = 8, 12.5%) in six E. coli and two K. pneumoniae, aac(3′)-II (n = 6, 9.4%) in four E. coli and two K. pneumoniae, aac(3)-III (n = 5, 22.7%), ant(2”)-I (n = 2, 3.1%), and aac(3′)-I (n = 1, 1.7%) in E. coli.
Resistance to sulfonamide was observed in 12 isolates (18.8%) mainly in E. coli (n = 11) (Table 2).
Analysis of clonality
High diversity was noted among the 22 E. coli strains, with 14 different profiles detected (Table 3). Five constituted major clones (including two or more isolates). The largest group included five isolates (ES1, ES8, ES12, ES14, and ES21). Singletons were observed in nine cases. Phylogenetic grouping revealed the presence of E. coli belonging to the four phylotypes A, B1, B2, and D with the repartition: A (n = 13, 59.1%), B1 (n = 2, 9.1%), B2 (n = 3, 13.6%), and D (n = 4, 18.2%). The main sequence type (ST) found were ST410 (n = 6, 27.3%), ST31 (n = 3, 13.6%), ST398 (n = 3, 13.6%), ST477 (n = 2, 9.1%), and ST131 (n = 2, 9.1%). The two ST131 isolates belonged to the international clone O25b:H4. The remaining strains were of the ST8, 21, 37, 66, 74, and 132.
In K. pneumoniae, rep-PCR revealed the existence of three different circulating clones among the isolates (Fig. 1) and five singletons. MLST identified ST14 (n = 3, 27.3%), ST39 (n = 3, 27.3%), ST219 (n = 3, 27.3%), and ST405 (n = 2, 18.1%).
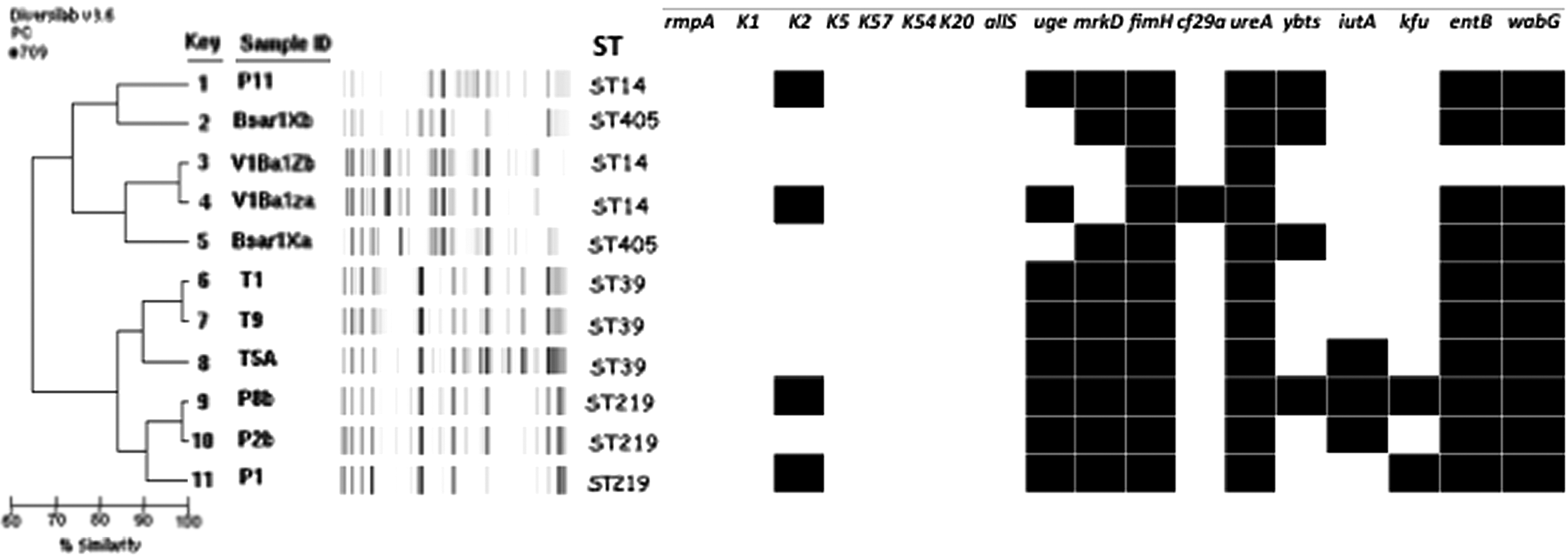
Dendrogram illustrating the genetic relatedness and the virulence gene profiles of 11 ESBL-producing Klebsiella pneumoniae strains based on the presence (black boxes) of a total of 18 virulence genes, phylotypes, sequence type, and additional resistance genes. ESBL, extended-spectrum β-lactamase.
Among the nine M. morganii strains, diversity was observed with four different profiles detected (including two or three isolates) and one singleton was observed. While among the eight C. freundii strains, four clones were noted and two contained three isolates and two singletons were observed. For the other species (E. cloacae and P. vulgaris), no clonality was observed (data not shown).
Detection of virulence-associated genes in E. coli and K. pneumoniae isolates
The distribution of the 19 virulence-associated genes tested in the 22 E. coli isolates is presented in Table 3. Three genes were present in all the isolates, including the adhesin gene fimH, the serum resistance-related traT, and the aerobactin gene iutA. In contrast, two genes were absent in all the studied strains, including the nonfimbrial adhesin gene papG1 and papG3. For iron metabolism, irp2 and iroN genes were detected in nine (40.9%) and eight (36.4%) isolates, respectively. cnf1 (40.9%) was more prevalent than hlyA (36.4%) in toxins-encoding genes.
The distribution of the 18 virulence-associated genes tested in the 11 K. pneumoniae isolates is presented in Fig. 1. All the isolates were negative for the capsule-encoding genes K5, K57, K54, and K20. The virulent hypermucoviscous K. pneumoniae associated with the magA and rmpA genes was also not found in our collection, while two virulence genes (ureA and fimH) were detected in all isolates. Finally, 4 isolates carried the K2 serotype gene. entB, wabG, and uge genes were particularly prevalent (10, 10, and 9 isolates, respectively).
Discussion
This study documents for the first time that wild fish in the Mediterranean Sea are reservoirs of enterobacteria resistant to oxyiminocephalosporins, aminoglycosides, sulfonamides, and quinolones. ESBL carriage is particularly high (21.3%), with a great diversity in species (E. coli, E. cloacae, K. pneumoniae, M. morganii, C. freundii, and P. vulgaris) and in ESBL enzymes detected (blaCTX-M, blaTEM, and blaSHV). This result suggests that these isolates were most likely derived from contamination of the fish from human sewage via river water and the growing amount of waste from urban, industrial, and agricultural activities discharged untreated into the sea near the coast.1,6 In our studied region, no control of this human sewage is performed. The sample area concerned fish living at more than 1,000 m off the coast, showing the large dissemination of the contamination. Furthermore, we report the predominance of the blaCTX-M gene that was found in 44 isolates (68.8%), in which blaCTX-M-15 was the predominant gene encountered (39/44). While CTX-M-1 has been reported to be very prevalent in animal isolates, CTX-M-15 is uncommon in animals, supporting the idea that human waste is a major source of ESBL genes in water. Detection of these β-lactamases in fish has been previously described in freshwater and farm fish in different parts of the world.17,36,37 The high prevalence of the CTX-M producers is in accordance with the observed data from community settings in the same region of Algeria.38,39 Moreover, CTX-M β-lactamases have also been identified in nosocomial and hospital environmental isolates from the same locality.40,41 Our study is consistent with previous studies that show the importance of water environments as critical reservoirs of different β-lactamase-encoding genes.42,43 However, it has been suggested that several human activities have caused the emergence and the diffusion of MDR bacteria from wild sources. 44
Coresistances to other antibiotic classes (e.g., aminoglycosides, cotrimoxazole and fluoroquinolones) are classically described in human ESBL-producing enterobacteria. We observed the same trend in our bacteria population, constituting a worrisome situation. Moreover, we link this coresistance with a high dissemination of aminoglycosides, PMQR and sulfonamide genes in wild fish. These genes are exclusively identified in E. coli and K. pneumoniae isolates, where 54.5% and 27.3% were MDR, respectively. The occurrence of the qnr determinant was particularly high (17.2%) in our isolates. qnrB2 and qnrS1 genes were associated with the presence of blaCTX-M-15 gene. qnrS1 gene was also linked to blaTEM-24 gene. These two qnr determinants have been previously described in many Gram-negative bacteria in farm fish in Egypt and Turkey.37,45 qnrA was also detected in Egypt in various enterobacteria, although we did not find this gene in our study. Comparison with clinical ESBL-producing enterobacteria isolated in Algeria confirms the emergence of PMQR genes in this country, but qnrB is the most prevalent qnr determinant followed by qnrS.41,46 Another interesting finding was the detection of the oqxAB gene in our isolates. This gene was present in all K. pneumoniae. To date, oqxAB has never been described in Algeria and was found for the first time in pigs. 47 In China, oqxAB gene was found in clinical E. coli and Salmonella isolates from food-producing animals.4,48 In E. coli an association between oqxAB and blaCTX-M has been suggested, 48 which we have observed in K. pneumoniae. Thus, the food chain might be a relevant vehicle for transmission of these enzymes in the environment. Finally, the studied strains also harbored aminoglycoside resistance genes. Very few data are available concerning this type of resistance, although one Danish article reported the presence of aac(3′)-II gene in E. coli isolated from both wastewater and patients. 49 Taken together, if we add the very high rates of resistance to aminoglycosides, quinolones, and cotrimoxazole in our collection, our study reflects the impact of the selective pressure due to an increased use of antibiotics in different settings (agriculture, animal food, and hospital).
This study showed a high diversity of clones among the E. coli isolates (14 different STs in 22 strains). For the first time we observed the presence of CTX-M-15-producing E. coli belonging to the worldwide clone O25b:H4-ST131. 50 Interestingly, these isolates harbored a virulence profile with diverse genes encoding adhesins and toxins as previously observed.2,51 Phylogenetic group B2, ST131 E. coli has become a dominant global human uropathogenic E. coli since the early 2000s, notably among those resistant to fluoroquinolones and/or producing CTX-M-15. 2 Moreover, our two isolates display an ExPEC status due to the presence of pap, afa/draBC, iutA, and kpsMT II. We also noted that the ST410 is mainly described to be associated with the ESBL determinant.52,53 All the studied E. coli had at least three virulence genes. FimH is known to contribute to the protection of E. coli from host heterophils. 54 This high prevalence has been observed in avian pathogenic E. coli.55,56 In Tunisia, Jouini et al. described the presence/absence of virulence genes in ESBL-producing E. coli recovered from chickens, cows, sheep, and a turkey sample. They noted the presence of fimA, aer, and eae and the absence of papGIII, papC, hly, or cnf1. 57 Finally, we observed that our E. coli collection belonged mainly to phylogenetic groups A and B1. These phylotypes correspond to commensal strains. However, in our collection these strains harbored different virulence determinants (fimH, iutA, traT). The balance between commensalism and infection depends on the status of the host and the production of virulence factors in the bacteria. In E. coli, the phylogenetic groups have also an influence on the intrinsic virulence of the bacterium. 58
In K. pneumoniae, we also noted the diversity of the clones detected. One of the major features leading to the worldwide spread of blaCTX-M-type genes corresponds to their association with successful clones. ST405 was described in European outbreaks involving strains from North Africa. 59 ST39 was previously associated with a nosocomial outbreak of ESBL-producing K. pneumoniae in pediatric and neonatal intensive care units from Annaba hospitals (Algeria). 60 ST14 was described around the world, notably in Oran hospital (Algeria). 61 Only ST219 has not been previously described. However, very few data have reported the presence of resistant K. pneumoniae in the environment or fish. A multidrug-resistant K. pneumoniae of ST11 causing sepsis in a free-ranging beaver (Castor fiber) has been noted in Switzerland. 62 ESBL-producing K. pneumoniae has been detected in rural water reservoirs in China63,64 and Bangladesh 65 and in rivers and lakes in Switzerland. 8 Our study highlights for the first time the propagation of highly prevalent clinical clones of K. pneumoniae in wild fish. Interestingly, we did not detect the presence of the new virulent, hypermucoviscous variant of K. pneumoniae. 33
In conclusion, this study is the first to demonstrate the existence of the presence of ESBL and other antibiotic resistance genes in wild fish. Our findings support the hypothesis that natural environments are potent reservoirs of MDR bacteria and associated genes. The importance of Enterobacteriaceae bacteria in aquatic environments as carriers of clinically resistance and virulence determinants was confirmed, and the need to more closely control water habitats as potential sources for the emergence and/or spread of antibiotic resistance and virulence gene in the environment was highlighted. The promotion of treatment systems of wastewater and the control/regulation of antibiotic consumption around the Mediterranean Sea are therefore essential.
Footnotes
Acknowledgment
We thank Sarah Kabani for her assistance in preparing and editing the article.
Disclosure Statement
No competing financial interests exist.