
Abstract
The occurrence of extended-spectrum beta-lactamase- (ESBL) and/or AmpC beta-lactamase- (AmpC) producing Enterobacteriaceae in livestock, especially in broiler fattening flocks, has been demonstrated in previous studies. Nevertheless, data on transmission routes of these resistant bacteria into the fattening farms are rare. Therefore, seven broiler fattening flocks were investigated for the occurrence of ESBL-/AmpC-producing Enterobacteriaceae during the course of the fattening period with the special focus on horizontal transmission routes. ESBL-/AmpC-producing Enterobacteriaceae from both individual animals and their housing environment were isolated at different time points and the housing environment was even sampled before the arrival of the chickens. All obtained ESBL-/AmpC-producing Enterobacteriaceae were examined for their bacterial species, Escherichia coli phylogroup, and occurrence of resistance genes. Selected isolates were further analyzed via whole-genome sequencing. All seven investigated flocks were tested positive for ESBL-/AmpC-producing Enterobacteriaceae with widely varying prevalence between the flocks. In one flock, the ESBL-/AmpC-producing Enterobacteriaceae were already detected in the housing environment before the arrival of the animals. In general, among the different types of ESBL-/AmpC-producing Enterobacteriaceae determined E. coli harboring a blaCMY-2 gene was the most frequent. Using whole-genome analyses we observed a horizontal transmission of ESBL-/AmpC-producing Enterobacteriaceae through contaminated housing environment as two flocks consecutively fattened in the same farm harbored closely related ESBL-producing isolates. This demonstrates the influence of a previous fattened flock on the ESBL-/AmpC-status of a following broiler flock and, therefore, the importance of hygiene measures on farm level.
Introduction
R
In addition, Projahn et al. 21 also showed a transmission of ESBL-/AmpC-producing Enterobacteriaceae from parent flocks into the hatchery. On the contrary, studies on broiler chickens assume that a contaminated barn might be an important factor for the occurrence of ESBL-/AmpC-producing Enterobacteriaceae.15,22 Luyckx et al. 23 support this hypothesis as they detected Escherichia coli in broiler houses even after cleaning and disinfection. Due to the presence of resistant Enterobacteriaceae even in empty fattening houses a colonization of the following animal flock seems possible. That transmission of ESBL-/AmpC-positive Enterobacteriaceae at farm level is defined as horizontal transmission in our study.
Aiming at an elucidation of these horizontal transmission routes on farm level, we conducted a long-term investigation of seven different broiler fattening flocks. The housing environment was investigated for the occurrence of ESBL-/AmpC-producing Enterobacteriaceae before the arrival of the chickens. Individual animals were sampled directly at arriving at the farm, ∼3–8 hours after hatching. In the following, the respective flocks were tracked during the fattening period by investigating both individual animals and their housing environment. Therefore, an evaluation of horizontal transmission routes for ESBL-/AmpC-producing Enterobacteriaceae in broiler farms was possible.
Materials and Methods
Sampled farms
Seven German broiler flocks (flock A–G, detailed information in Table 1), originating from ESBL-/AmpC-positive parent flocks, were investigated for ESBL-/AmpC-producing Enterobacteriaceae. The ESBL-/AmpC-status of the parent flocks was defined as positive when at least one out of four environmental samples (three pooled feces, one boot swab) was tested positive. The hatchlings from the investigated broiler flocks came from different parent flocks. For two flocks (E and F), a potential transmission from one flock to the following was investigated exemplarily. Therefore, the chickens originated from different parent flocks with different ESBL-/AmpC-producing E. coli strains (different resistance genes), and the flocks were fattened consecutively in the same barn. The chickens' transport from the hatchery to the farms took between 20 and 90 minutes. Only one of the seven flocks (flock A) was treated with an antibiotic (tylosin).
Tylosin on days 26–28, indication: enteritis caused by clostridia.
Flock F fattened consecutively to flock E in the same barn.
Preharvesting at day 31.
+, antibiotic treatment; −, no antibiotic treatment.
Sampling at the farm
The fattening flocks were analyzed at three different time points: first, second, and third sampling (Table 1).
The first sampling was carried out at the first day of the fattening period, both before and after the arrival of the chickens. The housing environment was investigated just before the animals arrived on the farm, except for flock C, where the samples were taken in parallel to the chickens' arrival. As environmental samples litter, feed, drinking water, and dust were collected from at least five different spots and a boot swab was taken by walking the whole length of the barn. Two air samples were collected according to Projahn et al. 21 In addition, different environmental swabs (Nerbe Plus, Winsen/Luhe, Germany) moistened with sterile phosphate-buffered saline (PBS, Phosphate Buffered Saline Tablets; Oxoid, Wesel, Germany) were taken from at least five different locations of water troughs, feeding troughs, the barns' walls (about 100 cm2) and, if possible, of the ventilator and hangers of the barns' equipment.
About half an hour later, directly after the arrival of the day-old chickens on the farm, 40 randomly chosen individual animals were sampled by taking cloacal swabs (Nerbe Plus).
The second sampling was carried out at the middle of the fattening period and the third sampling at the end. There, environmental samples and environmental swabs inside the barn were collected, analogous to the first sampling. In addition, a pooled feces sample consisting of at least 10 single randomly chosen feces heaps was collected. Again 40 individual animals were investigated by taking cloacal swabs. They were randomly chosen for every single sampling time point, which means that different animals were swabbed at the different samplings.
In flock F, that was fattened consecutively to flock E in the same farm house, an additional sampling was carried out 24 hours after the arrival of the chickens at the farm. At that time, 40 individual animals, pooled feces and a boot swab were investigated. Furthermore, for the same flock, pasture and soil were sampled at the third sampling from the outside area on the side of the barn where the exhaust air was emitted (“out”).
Laboratory methods
Sample preparation
Twenty gram each of the pooled feces, feed, and litter sample were mixed with Luria–Bertani bouillon (LB; Oxoid) in a 1:10 ratio, respectively, and 200 ml LB was added to the boot swabs. All samples were homogenized using a stomacher with 200 rpm for 2 minutes. Cloacal and environmental swabs as well as the air samples were investigated after enrichment in LB according to Projahn et al. 21 Dust samples were proceeded using PBS with the addition of 0.01% Tween 20 (Sigma-Aldrich, Darmstadt, Germany) as described by Laube et al. 15
After incubation (enrichment), 10 μl of each sample was streaked out on MacConkey Agar No. 3 (Oxoid) containing 1 mg/L cefotaxime (AppliChem, Darmstadt, Germany) (MC+) using an inoculation loop. The agar plates were aerobically incubated overnight at 37°C. Each batch of poured MC+ agar plates was verified by growth controls using an ESBL- and a non-ESBL-E. coli strain. Colonies with specific phenotypes grown on MC+ agar were suspected to be ESBL-/AmpC-producing Enterobacteriaceae. For each sample, at least 2 of those colonies per similar macromorphology were subcultured as isolates (up to 10 isolates per sample) for further investigations.
Species identification and phylogenetic grouping
For each isolate, the species was identified by matrix-assisted laser desorption time of flight (MALDI Microflex® LT and Biotyper database®; Bruker Daltonics, Bremen, Germany). Furthermore, the respective phylogroup of the E. coli isolates was determined by performing a multiplex PCR method 24 with modifications according to Projahn et al. 21
Antimicrobial susceptibility testing
For the selected Enterobacteriaceae isolates, suspected to be ESBL-/AmpC-producers, antimicrobial susceptibility was tested via agar disc diffusion according to the EUCAST guidelines 25 (Cefotaxime [30 μg], Cefotaxime [30 μg] with clavulanic acid [10 μg], Cefoxitin [30 μg] and Ceftazidime [30 μg]; Liofilchem® s.r.l., Roseto degli Abruzzi, Italy]. Inhibition diameters were measured and evaluated using the EUCAST breakpoint table. 26
Real-time PCR and sequencing
The detection of the most common class A beta-lactamase-genes blaCTX-M, blaSHV, blaTEM, and CIT-type AmpCs was performed for all ESBL-/AmpC-suspicious enterobacterial isolates as described by Roschanski et al. 27 Furthermore, the ESBL-/AmpC-genes were verified by sequencing at least one isolate of each sample using the primer set published by Projahn et al. 21
Whole-genome sequencing, single-nucleotide polymorphisms, Neighbor-Joining tree
To verify a transmission of ESBL-/AmpC-producing E. coli between two consecutively fattened flocks, their phylogenetic relationship was investigated by whole-genome sequencing (WGS). Therefore, 13 E. coli isolates with the same resistance gene (blaSHV12) and the same phylogroup (B1) from the consecutively fattened flocks E and F were chosen. Those isolates were selected both from individual animals as well as from environmental samples/swabs from different sampling time points. From flock E (the previously fattened flock), isolates from the third sampling time were analyzed: each one isolate of three environmental samples (boot swab, pooled feces, and litter), one environmental swab, and three randomly chosen cloacal swabs. For flock F (the consecutively fattened flock), isolates from all sampling times were investigated: each one isolate of two samples (boot swab and litter) from the first sampling, one isolate of a cloacal swab from the sampling time point 24 hours after arrival, each one isolate of a boot swab and a cloacal swab from the second sampling, and one isolate of the outside sample from the third sampling (Fig. 1).
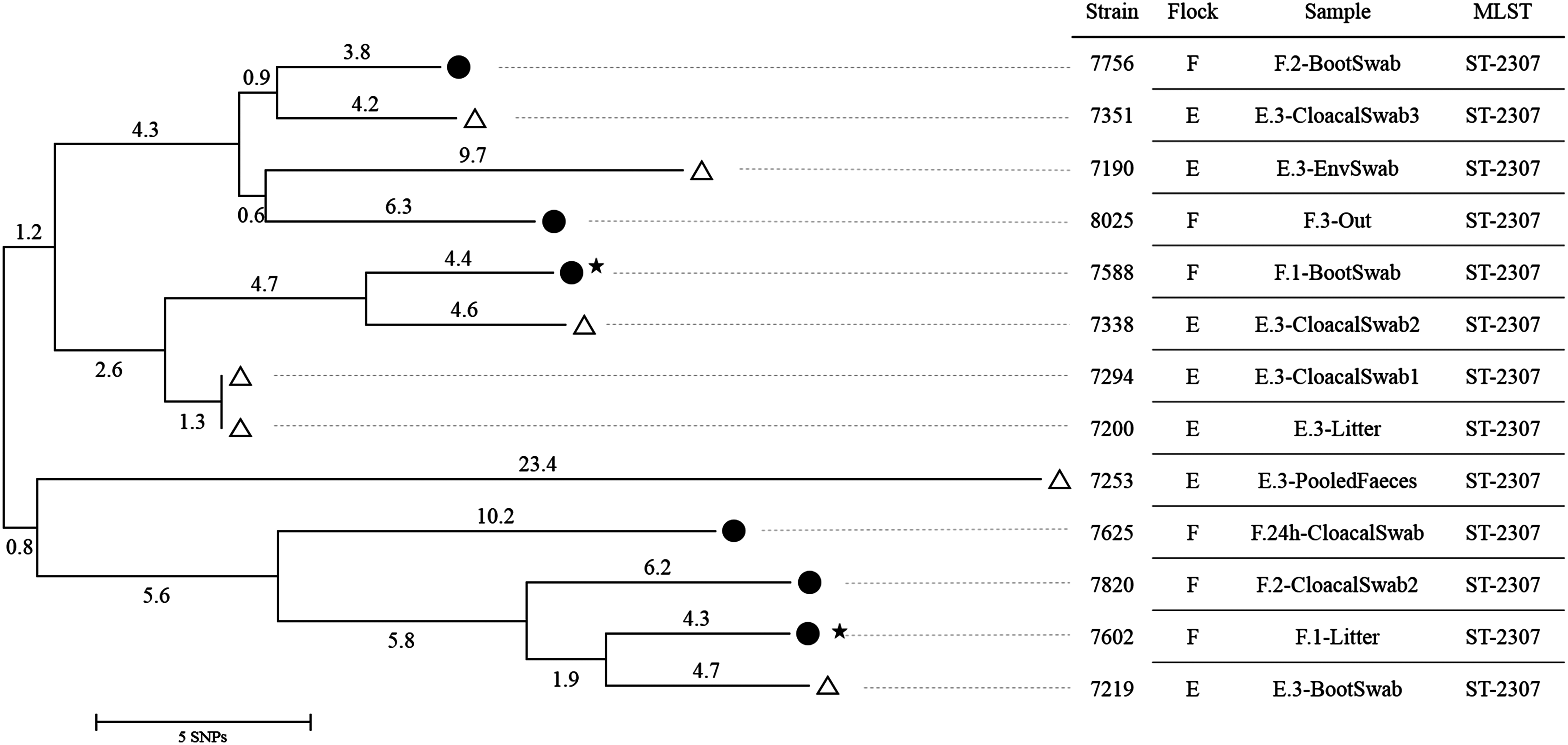
Neighbor-Joining tree for 13 Escherichia coli isolates (phylogroup B1, blaSHV-12) from flock E and F. •, flock F; ▵, flock E; ★, first sampling; before the chickens' arrival. E.3, flock E, third sampling; F.1, flock F, first sampling; F.24h, flock F, sampling at 24 hours; F.2, flock F, second sampling; F.3, flock F, third sampling.
DNA was extracted with the MasterPure™ DNA Purification Kit (Epicentre, Illumina) and WGS was performed by Illumina MiSeq 300-bp paired-end with a coverage between 50× and 120×. Quality control was performed with the NGS tool kit 28 and reads with a minimum of 70% of bases having a phred score greater than 20 were defined as high-quality reads. These resulted high-quality filtered reads were used for de novo assembly into contiguous sequences (contigs) using CLC Genomics Workbench 9.0 (Qiagen, Venlo, Netherlands). Multilocus sequence typing (MLST) was performed using MLSTFinder. 29
The core genome analysis and the numbering of single-nucleotide polymorphisms (SNPs) between the isolates were calculated with Harvest suite 1.0 using the parsnp algorithm, 30 a pairwise distances matrix was calculated using MEGA7.0, 31 and a Neighbor-Joining tree was constructed based on the number of SNP differences (1,000 bootstraps) using MEGA7.0 as well.
Results
Detection frequencies
In total, 1,133 samples were investigated. These include all 1,090 regular samples according the 3 samplings for all flocks (Table 2) as well as the 43 additional samples of flock F. Out of these samples, 289 were suspicious for ESBL-/AmpC-producing Enterobacteriaceae, and 1,261 isolates were obtained for further investigations. Using the described PCR approaches and antimicrobial susceptibility testing, the ESBL-/AmpC-production could be confirmed in 91.7% of the suspicious samples of flocks A–F (n = 253). In contrast, in flock G the ESBL-/AmpC-production was confirmed in 8.3% of the suspicious samples (n = 36), only. Overall, 235 samples (20.7%, n = 1,133) were tested positive for the ESBL-/AmpC-producing Enterobacteriaceae.
Env. samples: environmental samples (pooled feces, boot swab, litter, feed, drinking water, dust, air samples).
Env. swabs: environmental swabs (water troughs, feeding troughs, wall, ventilator, hangers of the barns' equipment).
The prevalence of the ESBL-/AmpC-producing Enterobacteriaceae at the three different sampling time points, detected in individual cloacal swabs, environmental samples and environmental swabs are shown in Table 2.
ESBL-/AmpC-producing Enterobacteriaceae could already be found in the boot swab and litter sample in one out of the seven flocks (flock F) before the arrival of the chickens. At the chickens' arrival at the farm, ESBL-producing Enterobacteriaceae were detected in cloacal swabs in two samples from one flock (flock B), only (0.7%, n = 280).
At the second and third sampling (middle and end of the fattening period), ESBL-/AmpC-producing Enterobacteriaceae were detected in all seven flocks, however, the prevalence between the seven flocks showed wide variations from 1.9% to 92.3% regarding all samples and with respect to the cloacal swabs from 0% to 100% (Table 2).
At the additional sampling at flock F, 24 hours after the chickens' arrival at the farm, 5% of the investigated cloacal swabs (n = 40) and both environmental samples (boot swab and pooled feces) were tested positive for ESBL-/AmpC-producing Enterobacteriaceae. The additional sample from the outside environment surrounding the exhaust air ventilators (“out”) at the third sampling of flock F was positive for ESBL-producing E. coli, too.
Isolate characterization
Various strains of ESBL-/AmpC-producing Enterobacteriaceae were detected in the seven investigated fattening flocks. Overall, E. coli strains belonging to phylogroups E, E/D, A, F, B1, and B2 encoding the CMY-2, SHV-12, SHV-2, and CTX-M-1 beta-lactamase enzymes were identified in isolates from all flocks and sampling times. Details for each flock are shown in Table 3. In addition, we isolated Klebsiella pneumoniae strains harboring the blaSHV-2 gene and Citrobacter freundii strains encoding for CIT-type AmpCs.
Overall number of positive samples does not correspond to the number of investigated samples (Table 2) as some samples harbor more than one ESBL-/AmpC-positive strain.
Hangers of the barn's equipment.
Feeding trough, wall, ventilator, hangers of the barn's equipment.
Feeding trough, ventilator.
Ventilator.
Feeding trough.
AmpC, AmpC beta-lactamase; ESBL, extended-spectrum beta-lactamase.
At the first sampling at flock C, CMY-2 encoding E. coli strains of phylogroup F were detected in the boot swab and litter samples. Isolates with equal characteristics were also found during the fattening period both in individual animal samples and environmental samples.
In flock E, E. coli isolates harboring the blaSHV-12 gene (phylogroup B1) were detected during the fattening period. In flock F, that was fattened consecutively to flock E in the same barn, E. coli isolates encoding the blaSHV-12 gene (phylogroup B1) were also detected throughout the entire fattening period, even in samples from the housing environment (boot swab and litter) before the animals arrived at the farm (first sampling). Furthermore, E. coli strains with characteristics (phylogroup B1, blaSHV-12) as detected in the housing environment and in individual animals in flock F also were detected in the sample from the outside environment surrounding the exhaust air ventilators (“out”) at the third sampling.
WGS, SNPs, Neighbor-Joining tree
Isolates with equal molecular characteristics (E. coli, phylogroup B1, blaSHV-12), detected in the consecutively fattened flocks E and F were further investigated. Based on the WGS-data, all 13 isolates were assigned to the MLST type ST-2307. The analyses of SNPs in the core genome showed a close relationship between the isolates of these 2 consecutively fattened flocks with maximum differences of 43 SNPs (Fig. 1).
Discussion
Many studies showed that ESBL- and/or AmpC-producing Enterobacteriaceae occur in broiler fattening farms with high prevalence.14,15,32 In our study, the prevalence of these resistant bacteria varied in a wide range between the seven flocks and even a flock without any positive cloacal swab was detected (flock G). In this flock, however, the environment was contaminated. This indicates that ESBL-/AmpC-positive housing environment does not necessarily result in high-prevalent flocks.
Interestingly, in recent studies ESBL/AmpC-producing Enterobacteriaceae were even detected in 1-day-old chickens, partially with high prevalence.15,20 In our study, only 0.7% of the 1-day-old chickens were tested positive. Previous studies discussed that chickens may get colonized with ESBL-/AmpC-producing Enterobacteriaceae by the uptake of these resistant bacteria from the environment of the hatchery 21 and arrive already colonized at the farms. 20 However, the time from the oral uptake to the colonization of the chickens' intestine and to the fecal shedding of commensal ESBL-/AmpC-producing E. coli is not known yet. Further investigations about that are needed. Therefore, the sampling time point in the first hours/days of life influences the detection of ESBL-/AmpC-producing Enterobacteriaceae. Maybe a sampling immediately following the chickens' placement in the barn after a short transportation time between 20 and 90 minutes, as performed in our study, did not allow to detect the resistant bacteria in cloacal swabs yet.
Another cause for the occurrence of ESBL-/AmpC-producing Enterobacteriaceae could be the use of antibiotics associating with a selection of resistant bacteria. 20 In our study, only one of the seven investigated flocks (flock A) was treated with a macrolide antibiotic at the end of the fattening period (day 26–28). However, even this flock was already tested positive for ESBL-/AmpC-producing Enterobacteriaceae before the antibiotic treatment (day 19). Therefore, the usage of antibiotics cannot be responsible for the occurrence or even a coselection of ESBL-/AmpC-producing Enterobacteriaceae with the observed high prevalence in the seven investigated broiler fattening flocks of up to 100% positive cloacal swabs. This is in accordance to Huijbers et al., 14 as they detected ESBL-/AmpC-producers even in organic farms without any antibiotic treatment. Furthermore, Guenther et al. 33 detected ESBL-producing E. coli also in wild birds that are normally not exposed to antimicrobial agents.
The housing environment of the barn itself seems to play an important role in the occurrence of resistant bacteria in broiler flocks. Luyckx et al. 23 showed that especially floor cracks and drain holes were difficult to clean and disinfect and, therefore, can still be contaminated with E. coli after disinfection. The occurrence of ESBL-/AmpC-producing Enterobacteriaceae in broiler flocks due to contaminated housing environment was also assumed in other studies.14,15,22 If resistant bacteria occur in a broiler flock and cannot be eliminated during the cleaning and disinfection procedure between two different flocks in one barn, remaining bacteria could be causative for the colonization of the chickens of consecutively fattened flocks. We confirmed this hypothesis of a circulation of ESBL-/AmpC-producing Enterobacteriaceae between following fattening flocks by determining close epidemiologic relationship between isolates from two consecutively fattened flocks (flock E and F). In this study, E. coli isolates of phylogroup B1, harboring the blaSHV-12 gene, as detected in the previously fattened flock E were also detected in the barns environment (boot swab and litter) before the animals of flock F arrived. Later on, isolates with these characteristics were found in animal and environmental samples of flock F. However, only two animal samples of the consecutively fattened flock F were positive in the whole fattening period. Maybe this transmission route does not result in high prevalence but in ESBL-/AmpC-positive broiler flocks. The dynamics of the spread of resistant bacteria, that are horizontally transmitted, should be investigated in detailed studies. In our study, 13 isolates with the same phylogroup and the same ESBL encoding gene both from the end of the fattening period of flock E (previously fattened flock) and from all sampling time points of flock F (consecutively fattened flock) were further investigated for the phylogenetic relationship using WGS. All these isolates belong to the MLST type 2307 and differ in at most 43 SNPs. On basis of the E. coli genome of ∼5 Mbp, the 43 SNPs (8.6 SNPs Mbp−1) represent 0.0012% divergence only. This overall low number of SNPs per Mbp is slightly higher than the one described for clonal EHEC strains during the German outbreak (1.8 SNPs Mbp−1).34,35 For some isolates from flock E and F (Fig. 1) the number of SNPs was 8 or 9 SNPs in total resulting in 1.7 or 2.0 SNPs per Mbp proving a direct clonal relationship of these isolates (E7351 vs. F7756 and F7602 vs. E7219).
Since we demonstrated this direct influence of a previous fattened flock on the ESBL-/AmpC-status on a following broiler flock due to contaminated housing environment, despite the cleaning and disinfection procedure, management measures should be further investigated in detail. In general, cleaning and disinfection procedures should reduce and minimize the load of bacteria in the barns and, therefore, should minimize the load of ESBL-/AmpC-producing Enterobacteriaceae. The procedures have to achieve such a low load of ESBL-/AmpC-producers that is insufficient to colonize the chickens and, therefore, cannot cause the spread of these bacteria in the broiler flock. To get more information on this, studies on the colonization of chickens with ESBL-/AmpC-producing Enterobacteriaceae and, here especially, the minimum necessary dose should be performed.
Also, the environment of flock C was contaminated with CMY-2 encoding E. coli strains of phylogroup F at the time the chickens arrived. In all other flocks, ESBL-/AmpC-producing Enterobacteriaceae were not detected in the barns environment before housing the animals, nevertheless, these resistant bacteria occurred in all flocks during the fattening period. However, even intensive samplings in the barns, as performed in our study, does not ensure the detection of all present bacteria, and therefore, horizontal transmission cannot be excluded. Maybe some resistant bacteria remained after cleaning and disinfection of the barns in a few locations. Since the barns with about 22,000 animals on a floor area of about 1,000 square meters are very big, it is impossible to analyze the entire environment, and we may have missed some ESBL-/AmpC-producers, which were already present at our first sampling. Also, our laboratory method was very sensitive with an antibiotic supplementation in the agar plates of only 1 mg/L cefotaxime. So, not all suspicious colonies are finally ESBL-/AmpC-producing bacteria. A confirmation of the ESBL-/AmpC-production using disk diffusion tests and sequencing of detected genes was necessary. In flock G, for example, the most suspected ESBL-/AmpC-positive colonies could not be confirmed (8.3% in contrast to 91.7% conformation rate for flocks A–F).
Interestingly, in flock F, closely related E. coli strains (phylogroup B1, blaSHV-12, MLST type 2307) were also detected in a sample from the outside environment (pasture and soil) surrounding the ventilators exhausting air. This indicates a possible airborne contamination of surrounding areas with ESBL-/AmpC-producing Enterobacteriaceae, as also shown by Laube et al. 36 Therefore, also a recurring entry of the bacteria due to contaminated shoes or barn equipment from the outside seems possible.
To summarize, in our investigation, we showed that ESBL-/AmpC-producing Enterobacteriaceae occurred in all seven investigated broiler flocks, even though the chickens were mainly tested negative at the beginning of the fattening period and an antibiotic treatment was performed in one flock only. In our study, the horizontal transmission of ESBL-/AmpC-producing Enterobacteriaceae through contaminated housing environment as a reason for the occurrence of these resistant bacteria in broiler flocks was proven with whole-genome analyses. Therefore, we showed the direct influence of a previous fattened flock on the ESBL-/AmpC-status of a following flock in the same barn, despite an all-in all-out management with a cleaning and disinfection between the flocks. To reduce the occurrence of ESBL-/AmpC-producing Enterobacteriaceae in broilers caused by this horizontal transmission, cleaning, and disinfection procedures in the barns should be improved and, therefore, need to be further investigated in detail.
Footnotes
Acknowledgments
We thank all farmers and all coordinating staff for their kind cooperation and permanent support. Many thanks to Nicole Roschanski and Sebastian Guenther for scientific advice and Maja Thieck for excellent technical assistance in the laboratory. This project was funded by the Federal Ministry of Education and Research (grant 01KI1313C) and is part of the RESET research consortium (![]() ).
).
Disclosure Statement
No competing financial interests exist.